
Abstract
We investigated for the first time the subtype distribution, prevalence of multiple HIV-1 infections, sexual networks, and partnership histories in a cohort of women engaged in high-risk sexual behavior such as female sex workers (FSWs) and women employed in entertainment facilities. Viral RNA was extracted from blood samples collected from 324 HIV-1-positive women; the gp-41 and pol-IN genes were directly sequenced. Women found to have closely related viruses and those with recombinant viruses were further analyzed in the pol-IN gene by clonal sequencing to determine HIV-1 multiple infections. Individual partnership histories were used to provide information on when sex work was undertaken and where. Subtyping in both gp-41 and pol-IN was successfully done in 210/324 (64.8%) women. Subtype distribution in these two genes was 54.3% (n=114) A/A, 2.9% (n=6) C/C, 24.3% (n=51) D/D, 11.9% (n=25) A/D, 4.8% (n=10) D/A, 0.5% (n=1) C/A, 1.0% (n=2) B/A, and 0.5% (n=1) B/D. Sexual networks were identified in six pairs and one triplet of women with closely related subtype A viruses. Partnership histories showed that women having phylogenetically similar viruses had worked in the same localities. Five cases of multiple infections were confirmed: four dual infections and one triple infection. In this first molecular epidemiology study among FSWs in Kampala, subtype A was the predominant subtype. About 9% of a subgroup had multiple infections. Partnership histories and multiple infections observed in this population suggest sexual mixing of the FSWs and their clients confirming their high-risk characteristics.
Introduction
T
In sub-Saharan Africa, all of these subtypes, CRFs, and their recombinants are cocirculating. The frequent surveillance of circulating subtypes, recombinants, and transmitted resistance mutations in a population is essential in preparation for vaccine studies, roll out of antiretroviral therapy, and other interventions. Molecular epidemiology has also been used to track transmission within risk groups.
Work in Uganda has identified subtypes A and D and their recombinants to be predominantly circulating. In rural southwestern Uganda, subtype D is reported to be the major circulating subtype, 3 –7 whereas in the central urban areas, some reports have shown an equal distribution of subtype A and D or in some cases a predominance of subtype A. 8,9
Several studies from East Africa including Tanzania and Kenya have documented high rates of HIV-1 among women involved in high-risk sexual behaviour such as sex workers and bar workers. 10 –20 Groups with high HIV prevalence and incidence, such as female sex workers (FSWs), can provide valuable opportunities to conduct HIV molecular epidemiological studies. A recent UNAIDS 2010 report provided evidence that such populations contribute to the spread of the epidemic; it is estimated that almost one-third of new HIV infections in Ghana, 14% in Kenya, and 10% in Uganda are linked to sex work. 21,22
Several studies have been done on FSWs in Africa showing the predominance of different subtypes in particular geographic regions 23 –26 ; however, little information is available from Uganda. Our group conducted a study along the Trans-Africa Highway to determine the relationship between the HIV strains and sexual mixing of residents. 27 The major subtypes identified were A and D, but there was no evidence of clustering of sequences associated with place of residence as had been documented by sociological studies of sexual networks in these villages. 28 –30
People at high risk of HIV infection are likely to be superinfected or coinfected, providing opportunities for viral recombination contributing further to HIV diversity. We have recently shown in our cohorts recombination among individuals with multiple infection. 31 Our recently established cohort of women involved in high-risk sexual behavior in Kampala Uganda found that 37% of the study participants were HIV positive, 32 compared to 7% in the general population (2004–2005 National HIV serobehavioral survey).
In this article we report the findings of a study undertaken to describe the HIV-1 subtypes circulating in women engaged in female sex work in Kampala, to estimate the prevalence of multiple infections. We also use partnership histories of the women identified as having phylogenetically similar viruses to understand better the HIV epidemic in this high-risk population.
Materials and Methods
Study subjects
The Good Health for Women Project (GHWP), a cohort of 1027 women involved in high-risk sexual behavior in Kampala and therefore at high risk of acquiring and/or transmitting HIV-1 and other sexually transmitted infections (STIs), was established between March 2008 and April 2009. All women, either self-reporting sex workers or women employed in local entertainment facilities in Kampala, were eligible for enrollment. At three monthly intervals, sociodemographic and behavioral information was collected using structured questionnaires. In addition, a gynecological examination was performed, and blood and genital samples were tested for HIV and other STIs. All consenting women had access to the GHWP clinic offering free primary and reproductive health care including screening for and treatment of STIs, family planning, antenatal care, voluntary HIV counseling and testing (VCT), health education, and condom distribution. Women who required initiation of antiretroviral treatment (ART) and other HIV-related care were referred to qualified health centers.
Blood samples were collected in EDTA tubes from 324 consenting HIV-infected women with unknown seroconversion dates at testing. Samples were transported to the laboratory within 12 h of collection and plasma prepared by centrifugation at 400×g for 10 min. Two cryovials of plasma and one of whole blood were stored at −80°C for further research studies. In addition, as part of our data collection, women were invited to tell their life histories, including partnership history, to a social science team member. A detailed description of this cohort has been reported. 32
Ethical considerations
The study was approved by the Uganda Virus Research Institute (UVRI) Institutional Review Board and the Uganda National Council of Science and Technology (UNCST). All participants provided written informed consent for collection and subsequent analysis of specimens.
RNA extraction and RT-nested PCR
Viral RNA was extracted from cryopreserved plasma using QIAamp Viral RNA Mini Kit (Qiagen, Hilden, Germany). Reverse transcription (RT) and first round polymerase chain reaction (PCR) was done using the QIAGEN OneStep RT-PCR Kit (Qiagen, Hilden, Germany) to amplify the gp-41 and pol-IN genes using universal primers.
Envelope gp-41 analysis
First round PCR primers GP40F1 (forward; 5′TCTTAGGAGCAGCAGGAAGCACTATGGG3′; nucleotides 7789 to 7816) based on HXB2 and GP41R1 (reverse; 5′AACGACAAAGGTGAGTATCCCTGCCTAA3′; nucleotides 8347 to 8374) and the nested PCR, primers GP46F2 (forward; 5′ACAATTATTGTCTGGTATAGTGCAACAGCA3′; nucleotides 7850 to 7879) and GP47R2 (reverse; 5′TTAAACCTATCAAGCCTCCTACTATCATTA3′; nucleotides 8281to 8310), were used to amplify a 460-bp gp-41gene product. 33
The RT-PCR reaction contained 10 μl 5X buffer, 2 μl dNTP mix, 2 μl Taq enzyme mix, 0.5 μl RNase inhibitor, 3 μl 10 μM Primer GP40F1, 3 μl 10 μM Primer GP41R1, 4.5 μl water, and 25 μl of RNA. The cycling conditions were 50°C 30 min; 95°C 15min (95°C 30 s, 54°C 30 s, 72°C 90 s)×40 72°C 10 min 4°C hold. The second round PCR reaction contained 10 μl 10X PCR buffer, 4 μl 10 mM dNTP mix, 10 μl 1.5 mM MgCl2, 2 μl of each 10 mM Primer GP46F2/GP47R2, 0.6 μl Taq Polymerase, 61.4 μl dH2O, and 10 μl cDNA. The cycling conditions were 1 cycle of 94°C for 5 min, 35 cycles of 94°C 10 s, 50°C 45 s, 72°C 1 min, then 1 cycle of 72°C 10 min and a final hold at 4°C.
pol-IN analysis
Viral RNA was extracted as earlier described and the pol-IN amplified using first round PCR primers Unipol5/Unipol6 and second round primers Unipol1/Unipol2. The PCR thermocycling reaction conditions have been described elsewhere. 31
The gp-41 and pol-IN PCR products were visualised on a 1.5% agarose gel to confirm positive PCR amplification and purified using the QIAquick PCR Purification Kit (Qiagen, Hilden, Germany).
Sequencing and phylogenetic analysis
The purified PCR products were directly sequenced in the sense and antisense directions with primers GP46F2 and GP47R2 for gp-41 and Unipol1 and Unipol2 for pol-IN. The sequencing procedure has been described previously.
31
Subtype reference sequences of HIV-1 group M available from the Los Alamos Sequence database (
pol-IN clonal sequencing and phylogenetic analysis
Individuals found to have phylogenetically similar viruses and those with recombinant viruses were analyzed further in the pol-IN gene. The cleaned pol-IN secondary PCR products were cloned, and between 10 and 20 positive clones were sequenced using Unipol1/Unipol2 primer sets as previously described. 31
Partnership histories
Life histories, which included their partnership history, were collected from 200 women, including the women identified as having phylogenetically similar viruses. This was part of a social science GHWP substudy of the social context of sexual partnerships among women at high risk of STI infection. Three in-depth interviews were conducted with each woman over a period of 1 month, gathering information on their day-to-day life and their sexual partnerships.
Results
HIV-1 subtyping and phylogenetic analysis
We were able to obtain subtype sequence data for 244 gp-41 (75.3%) and 245 (75.6%) pol-IN out of the 324 individual samples analyzed. The failure to obtain sequence data was either due to failed PCR or failed sequencing probably as a result of low viral loads as some of the women were reported to be on ART. Sequencing failure could also have been the result of PCR primer mismatch due to the high diversity of HIV-1 subtypes in this population. Subtype sequence data for both gp-41 and pol-IN were available for 210 (64.8%) individuals (Table 1). Of the subtype distribution in the gp-41 gene, 64.3% (n=157) were subtype A, 1.2% (n=3) B, 3.7% (n=9) C, and 30.7% (n=75) D. The subtype distribution in the pol-IN gene was 59.2% (n=145) A, 2.9% (n=7) C, and 38.0% (n=93) D. The overall subtype distribution in both gp-41/pol-IN genes was 54.3% (n=114) A/A, 2.9% (n=6) C/C, 24.3% (n=51) D/D, 11.9% (n=25) A/D, 4.8% (n=10) D/A, 0.5% (n=1) C/A, 1.0% (n=2) B/A, and 0.5% (n=1) B/D (Table 1, Figs. 1 and 2).
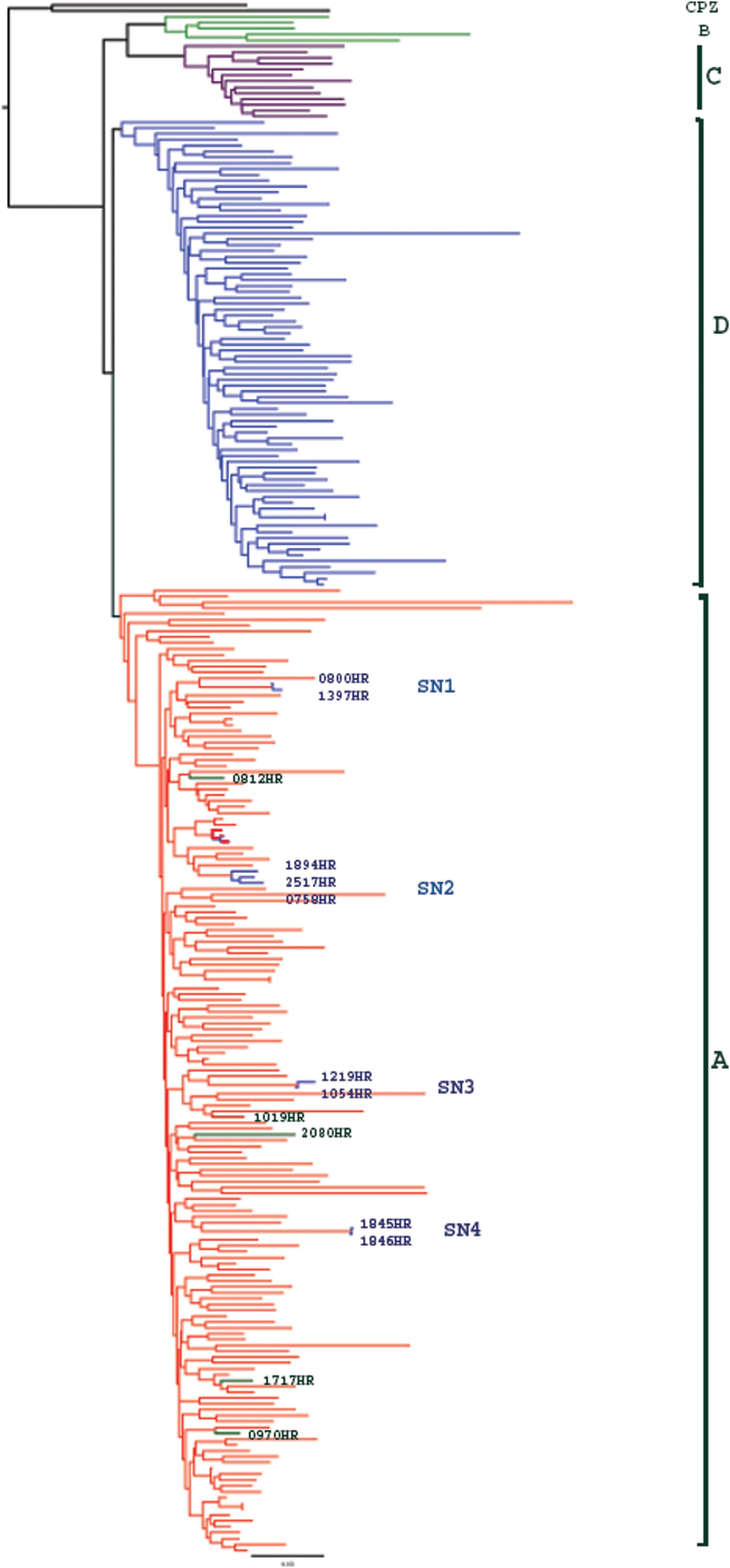
Phylogenetic analysis of the gp-41 sequences (HXB2 location 7850–8310) for 244 participants. Subtype A is colored red, B green, C purple, and D blue. Sexual Networks (SN) 1, 2, 3, and 4 are shown. The phylogenetic plots represent consensus sequences of the participants. Color images available online at

Phylogenetic analysis of the pol-IN sequences (HXB2 location 4470–4807) for 245 participants. Subtype A is colored red, C purple, and D blue. Sexual Networks (SN) 1, 3, 5, 6, and 7 are shown. The phylogenetic plots represent consensus sequences of the participants. Color images available online at
Analysis of the gp-41 gene showed that there were three pairs (n=6) of women that each had phylogenetically similar subtype A viruses, possibly indicating that each pair or sexual dyad (SN1, 3, and 4) of women may have been part of the same sexual network, with the same source of infection (Fig. 1). The average genetic distances were SN1 (1.5%), SN3 (1.5%), and SN4 (0.6%). To rule out the possibility that phylogenetically similar viruses observed in women was due to sample contamination, we sequenced samples from the same women bled at their subsequent clinic visit and we still observed phylogenetically similar viruses. In addition, all these women had different clinic visit dates and their samples were processed and sequenced on different dates, which further ruled out sample contamination. We further observed another group of three women (SN2) who also had phylogenetically similar subtype A viruses (Fig. 1) with an average genetic distance of 3.6%.
Analysis of the pol-IN gene also showed five pairs (n=10) of women (SN1, 3, 5, 6, and 7) that each had phylogenetically similar subtype A viruses, again possibly indicating that infections in each pair originated from the same source (Fig. 2). The average genetic distance in SN1, 3, 5, 6, and 7 was 0.9%, 2.3%, 2.7%, 1.4%, and 0.5%, respectively. Two pairs of women with similar viruses identified in pol-IN had also been identified in the gp-41 gene (SN1and 3). The sexual network links that were not identified in both genes were mainly due to failure to generate sequence data from both genes even after repeated sequencing, as was the case in SN4 where data were missing for 1845HR in the pol-IN gene.
One observation was that in some groups of individuals, the viruses clustered together in one gene and differently in another gene as was the observation in SN2, 5, 6, and 7 (Figs. 1 and 2). In another observation 1894HR's virus clustered with 2517HR and 0758HR in gp-41(Fig. 1), but in pol-IN her virus clustered with 1019HR (Fig. 2). We were, however, unable to observe the clustering of 2517HR and 0758HR as we failed to sequence 2517HR in the pol-IN.
When we did clonal sequencing of the pol-IN gene of all sexual networks (n=15) and those with gp-41/pol-IN recombinant viruses (n=39), we identified 5 (9.3%) cases of multiple infections out of the 54 samples that were analyzed (Fig. 3). The multiple infections included three intrasubtype A dual infections (0785HR, 0792HR, and 1858HR), one intrasubtype D dual infection (1840HR), and one triple infection with subtype D and two intrasubtype A strains (0992HR).

Phylogenetic analysis of the pol-IN clonal and direct sequences (HXB2 location 4470–4807) for the multiple infected participants 0785HR, 0792HR, 0992HR, 1840HR, and 1858HR. Color images available online at
Partnership histories
The individual partnership histories provided information on when sex work was undertaken and where. This is shown in the summaries of four dyads (parts of the four sexual networks).
Dyad 1
1397HR lives and works as a cooked food seller in the area in which many of the FSWs work. She says she can count the number of partners she has had on the fingers of her hands. She found out she was infected with HIV in 2008. In 2007 she had started a 3-year relationship with a man who was “always hungry for sex.” She knows he visited FSWs. 0800HR began work as an FSW in 2007 when a relative who had helped since her parents died stopped sending money. She needed money to support her siblings. She works in the area in which 1397HR lives and works. 0800HR has many different casual partners.
Dyad 4
1846HR was introduced to sex work in 2005 by a friend. She used to sell cooked food in Kampala and sex work supplemented her income. She tested positive for HIV in 2008. She has had many different partners whom she meets in lodges and bars. 1845HR lost her husband to AIDS in 2003. She became an FSW soon after because she needed money. In 2005 she got a regular partner and had a child in 2006. When she was pregnant she learned she was infected with HIV. She continued as an FSW after the birth, working in the same lodges as 1846HR.
Dyad 6
2080HR began work as an FSW at a truck stop outside Kampala in 2001. She has worked as an FSW in Kampala since 2002 and she tested positive for HIV in 2005. She currently runs a bar and provides FSW only on an occasional basis because she is not well. The other woman in this dyad, 0970HR, began FSW in 2004 in Kampala. She has a routine of rotating between bars on different nights of the week, including those in the area where 2080HR has worked. She tested positive for HIV in 2007 when she joined GHWP.
Dyad 7
1019HR started FSW in 2003 when she came to Kampala after her marriage dissolved. She has worked only in Kampala and always in the same area in southern Kampala. She tested positive for HIV when she joined GHWP in 2007. 1894HR lost her long-term partner to HIV in 2000 in Kampala. She began work in a bar in a border town after his death, and provided sex to some men to get extra food. She then worked at fish landing sites as a bar girl in Uganda and Tanzania before moving back to Kampala in 2004. She works as an FSW in the same bars as 1019HR.
Discussion
This is the first study in Kampala, Uganda that describes the subtype distribution of HIV-1 in a cohort of high-risk sexual behavior FSWs. We have shown that in this population, subtype A is more prevalent (60%) than the other subtypes and recombinants. Some studies done earlier in Kampala and a few surrounding towns had shown either a predominance of subtype A or an equal distribution of subtype A and D 8–9, 38 ; this study shows that subtype A has remained the most prevalent, although with a trend suggestive of an increase in the ratio of subtype A compared to D.
A change in prevalence of subtype A and D has been observed in some rural cohorts in southwest Uganda. Over the years, subtype D has been more prevalent than subtype A 4,6,39 however; recent reports from Rakai district in Uganda have shown that the prevalence of subtype A has increased in this region of Uganda. 40 The Rakai study showed a significant decrease in the prevalence of subtype D from 70.2% in 1994 to 62.4% in 2002 and a significant increase of subtype A from 16.7% in 1994 to 23.3% in 2002.
The observed changes in subtype prevalence over the years may have several explanations but none is entirely satisfactory. The migratory nature of some of the populations in these regions may facilitate the in/out movement of individuals infected with different subtypes over the years. 41 A study in Rakai has shown that subtype A has a significantly higher rate of heterosexual transmission than subtype D 42 and this coupled with earlier findings that individuals infected with subtype D had a faster disease progression compared to those infected with subtype A 4,7,43,44 may further explain why subtype A has remained or is becoming more prevalent in our populations.
However, our observation of the subtypes in this cohort may not be a true reflection of the circulating subtypes due to the fact that we sequenced a small fragment of only two genes. It may be that this is the reason that we were able to identify only 18.6% (n=39) of viruses as intergene recombinants and yet in populations that are considered to be low-risk sexual behavior populations, 30% of the viruses were either intragene or intergene recombinants. 39 It is therefore possible that the prevalence of recombinants may be higher than that observed if we had sequenced more or larger gene fragments.
The observed links (parts of larger sexual networks) when we analyzed the gp-41 and pol-IN among some women are indicators of possible frequent partner exchange among the FSWs.
In the gp-41, we identified four sexual networks whereas five sexual networks were observed in the pol-IN. However, the sexual network links were observed in both gp-41 and pol-IN genes in only two networks, with the rest of the sexual network links observed in only one gene. We were unable to observe networks in both genes for some of the networks because of missing sequence data for some women in their respective networks as was the case in SN4. In one of the networks though (SN2), in gp-41 the women had phylogenetically similar viruses but phylogenetically different viruses in the pol-IN gene. This observation can be possible if one of the women was infected with more than one virus that resulted in a recombinant virus or infected with a pure subtype and a recombinant virus. In such a case scenario, it is possible to select the same virus in one region but a different virus in another region from the two women, an observation that has been made in another study in Kenya. 45 There is therefore a possibility that observation of a sexual network link in one gene and nonobservation in another may be an indication of multiple or recombinant infections.
The summaries of the partnership histories provide information that supports the phylogenetic clusters that were observed in the seven sexual networks. The women not only have multiple partners but have also worked in the same bars, at truck stops, fish landing sites, and border towns, facilitating sexual mixing and the transmission of different forms of the virus.
In our cohort, the prevalence (9.3%) of multiple infections in the 54 women whose samples were cloned and sequenced was relatively low compared to that reported from most of the high-risk populations; however, lower rates of multiple infection (3%) have been reported in a cohort of commercial sex workers in Burkina Faso. 25 A study in the Mbeya region in Tanzania showed that the prevalence of multiple infections in the low-risk and high-risk sexual behavior populations was 9% and 19%, respectively, 11 which is more than we observed. Even in a low-risk sexual behavior population in Rakai, Uganda, an 5.8% prevalence of multiple infections was found. 44
The differences observed in the prevalence of multiple infections in these populations may also be a result of the methods used. In our study, we used both direct sequencing and clonal sequencing of the pol-IN gene. By analyzing only one gene, there is a possibilty of not detecting multiple infections that result in recombinant viruses 45 and this may have been the case in our study. The study in Mwanza and Rakai used the Multi-region Hybridisation Assay (MHA), a PCR subtype-specific probe-based assay across five genomic regions. 46 By analyzing at least five genomic regions, the chances of identifying more multiple infections and recombinants are increased; however, for this assay to work best, probes must be well designed to identify the circulating subtypes within a population. On the other hand, MHA could also overestimate multiple infections if nonspecific probe binding occurs.
The relatively low prevalence of multiple infections in this population can also be due to reduced high-risk behavior after HIV diagnosis. Some studies have documented changes in sexual behavior before and after HIV diagnosis among sex workers. 47 –49 Another cohort in Kenya demonstrated that after seroconversion, women reported safer sexual behaviors such as abstinence, 100% condom use, fewer partners, and fewer sexual contacts. 50
In conclusion, in this first molecular epidemiology study among FSWs in Kampala, subtype A was the predominant subtype. About 9% of a subgroup had multiple infections. The partnership histories, partial sexual networks, and multiple infections observed in this population suggest sexual mixing in the FSWs and their clients and confirm their high-risk characteristics.
Sequence Data
The sequences have been deposited into GenBank under accession numbers HQ709446–HQ710484.
Footnotes
Acknowledgments
D.S., N.N., F.L., J.B., J.S., J.V., H.G., and P.K. contributed to the study design and writing of the article. D.S., N.N., and F.L. contributed to the laboratory experiments and phylogenetic analysis. J.B., J.V., and J.S. contributed to the analysis of the partnership histories. H.G. and P.K. were responsible for the overall supervision of the study. We are grateful to Prof. George William Lubega for reviewing the article. The authors are also grateful to the participants and the clinic and laboratory staff of the MRC/UVRI. Thanks to Brian Magambo, Maria Nanyonjo, and Anne Kapaata for laboratory technical assistance. Funding for this study was provided by the Medical Research Council UK, the European and Developing Countries Clinical Trials Partnership (EDCTP CG_ct_05_33070), and EDCTP Senior Fellowship award TA.2007.40200.011.
Author Disclosure Statement
No competing financial interests exist.