
Abstract
Our objective was to determine whether monitoring HIV-1 DNA concentration or new resistance mutations in peripheral blood mononuclear cells (PBMCs) during effective antiretroviral therapy (ART) predicts virologic failure. A retrospective analysis used blood specimens and clinical data from three nevirapine containing arms of a four-arm, open-label, randomized trial comparing ART regimens in HIV-1-infected children who had failed mono- or dual-nucleoside therapy. Sensitive assays compared cell-associated HIV-1 DNA concentrations and nevirapine (NVP) and lamivudine (3TC) resistance mutations in children with plasma HIV-1 RNA <400 copies(c)/ml who did or did not experience subsequent virologic failure. Forty-six children were analyzed through the last available follow-up specimen, collected at 48 (n=16) or 96 (n=30) weeks of ART. Thirty-five (76%) had sustained viral suppression and 11 (24%) had plasma viral rebound to ≥400 c/ml (virologic failure detected at a median of 36 weeks). HIV-1 DNA levels at baseline, 24, 48, and 96 weeks of ART were similar in children who did vs. did not experience virologic failure (p=0.82). HIV-1 DNA levels did not increase prior to viral rebound. NVP resistance mutations were detected in 91% of subjects in the failure group vs. 3% in the suppressed group (p <0.0001). Among nine evaluable children, NVP mutations were first detected prior to virologic failure in two (22%), at viral rebound in five (56%), and after failure in two (22%) children. HIV-1 DNA concentrations did not predict virologic failure in this cohort. New drug resistance mutations were detected in the PBMCs of a minority of virologically suppressed children who subsequently failed ART.
Introduction
A
HIV-1 causes persistent infection of a subset of host cells. 6 This reservoir of viral DNA is established soon after infection, 7 and is variably diminished by ART. 8 –10 HIV-1 DNA in peripheral blood mononuclear cells (PBMCs) has been quantified in studies of viral dynamics and pathogenesis. However, it remains unclear whether this marker could be used to monitor treatment efficacy and anticipate failure. HIV-1 DNA concentrations predicted disease progression in adults treated with mono- or dual-ARV. 11 In addition, during effective ART children with ≥1 log10 decrease in PBMC HIV-1 DNA had no evidence of low-level viral replication while those with stable HIV-1 DNA had genetic evidence of ongoing low-level viral replication, indicated by a shift in the prevalence of resistance mutations selected by the ARV in their ongoing ART. 12 Thus, we hypothesized that monitoring HIV-1 DNA concentration in PBMCs during effective ART would predict virologic failure.
Moreover, multiple studies conducted during effective ART have detected evidence of low-level viral replication in a subset of individuals. 12 –17 Given that low intracellular ARV concentrations allow viral replication and the selection of drug-resistant HIV-1, we hypothesized that the detection of new drug resistance mutations during suppressive ART would predict subsequent virologic failure.
The Pediatric AIDS Clinical Trials Group (PACTG) 377 Study was a prospective randomized, open-label, four arm trial that provided the opportunity to study children receiving ART that included one or two drugs, nevirapine (NVP) and lamivudine (3TC), with low genetic barriers to resistance. 4,18 The PACTG 377 Study was completed when this substudy was conceived. Virologic failure had occurred in approximately 25% of the participants who had ART-associated suppression of viral replication. The objectives of this study were to evaluate whether the detection of new drug resistance mutations in PBMCs during effective ART or if monitoring viral DNA load in PBMCs could predict virologic failure.
Materials and Methods
Study subjects
The subset of children who achieved suppression of plasma HIV-1 RNA to <400 c/ml while taking a NVP-containing ART regimen in PACTG 377 4,18 was studied (see Table 1). PACTG 377 enrolled 181 HIV-1-infected children aged 4 months to 17 years from December 1997 through September 1998. All children were naive to stavudine (d4T), 3TC, protease inhibitors (PI), and nonnucleoside reverse transcriptase inhibitors (NNRTI). Between 44% and 69% of children in each study arm achieved undetectable viral loads (HIV-1 RNA <400 c/ml) after 12 weeks of therapy, and 30–52% had sustained HIV-1 RNA suppression after 48 weeks of therapy. Stored PBMC specimens, collected at 12-week intervals between weeks 0 (baseline) and 96 of ART, were analyzed from children who had specimens from baseline and/or the 12-week visit, and specimens from at least two subsequent study visits. The linkage of the HIV-1 RNA levels and the virologic outcome for each subject were not provided to the substudy team until after the analysis of PBMCs was completed.
Four children were excluded from the calculation due to HIV RNA <400 c/ml at baseline; the total population used for Arm B was n=50 and for Arm D was n=42.
Includes all subjects with specimens available in the repository.
ART, antiretroviral therapy; d4T, stavudine; NVP, nevirapine; RTV, ritonavir; 3TC, lamivudine; NFV, nelfinavir; ddI, didanosine; ZDV, zidovudine.
Quantification of HIV-1 DNA
Genomic DNA was extracted from PBMC aliquots using the Puregene DNA Purification Kit (Gentra Systems, Minneapolis, MN). To determine HIV-1 DNA concentrations, HIV-1 gag and human beta-globin DNA (used to normalized HIV-1 to host DNA) were quantified in each specimen by real-time polymerase chain reaction (PCR). 19 All standards and samples were assayed in duplicate in the iCycler IQ Real-Time Detection System (Bio-Rad, Hercules, CA), and each subject's specimens were batched in the same assay run. The mean intraassay and interassay variabilities for the real-time assay of HIV-1 gag control across multiple assays were 0.03 and 0.05 log10 copies, respectively. The mean intraassay variability among the test specimens was 0.09 log10 copies. The HIV-1 DNA concentration was expressed as the log10 copies per 106 PBMCs.
HIV-1 genotyping
HIV-1 drug resistance mutations were detected using an oligonucleotide ligation assay (OLA)
20,21
(
Statistical analyses
The nonparametric Wilcoxon test was used for comparing continuous variables by virologic outcome, such as baseline age, CD4 cell counts, HIV-1 DNA/RNA levels, or changes in HIV-1 DNA concentration from baseline to virologic failure. When comparing the within-subject changes of DNA levels from prior to virologic failure, to virologic failure, and to the subsequent visit, paired Wilcoxon signed-rank test was performed.
Fisher's exact test was performed for categorical variables, comparing results between virologic failure and nonfailure groups, such as detection of the 215F/Y mutation at baseline, new NVP resistance mutations, and other parameters. The Freeman–Halton exact test was applied for multiple comparison groups, e.g., comparing the frequency of virologic failure among the three different study arms.
The Wei–Johnson nonparametric global test was performed to compare the overall trends of HIV-1 DNA levels during the study follow-up period (i.e., from baseline to study weeks 12, 24, 36, 48, and 96) between the virologic failure and nonfailure groups (Fig. 1A).
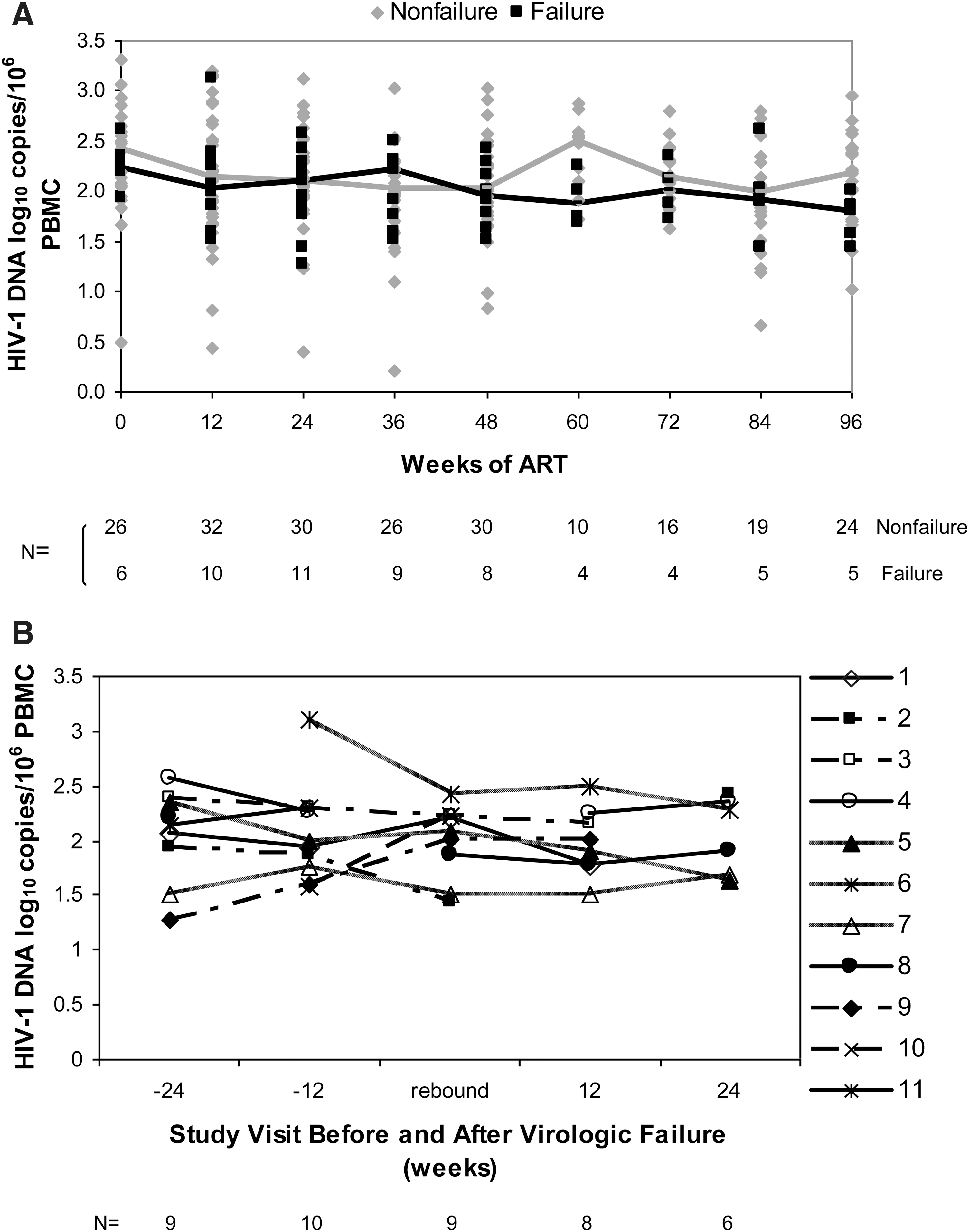
HIV-1 DNA concentrations in the peripheral blood mononuclear cells (PBMCs) of children before and during up to 2 years of antiretroviral therapy (ART).
Results
Study population
Specimens from 46 children were analyzed: 11 randomized to study Arm A [d4T, NVP, ritonavir (RTV)], 13 randomized to Arm C [d4T, NVP, nelfinavir (NFV)], and 22 randomized to Arm D (d4T, 3TC, NVP, NFV) (Table 1). Of these, 30 children had specimens through 96 weeks of follow-up, while 16 had specimens through 36–48 weeks. Baseline HIV-1 plasma RNA and CD4+ cell counts were similar for children in the nonfailure and failure groups (Table 2). Viral replication was suppressed (plasma HIV-1 RNA levels <400 c/ml) in the majority of children (44/46) by week 12 of ART and in the remaining two children by week 24. Thirty-five (76%) of the 46 children had sustained suppression of viral replication for the duration of follow-up (nonfailure group) and 11 (24% of all children; 27% of children in Arm A, 38% in Arm C, and 14% in Arm D, p=0.24) experienced virologic failure, defined as plasma HIV-1 RNA rebound to ≥400 c/ml after an initial reduction to undetectable levels (failure group, Table 2). The time to viral rebound and the HIV-1 RNA load at time of rebound were similar in the three study arms, with rebound at a median of 36 weeks (range 24–48) and a median rebound plasma HIV-1 RNA of 3.68 log10 c/ml (range 2.65–4.43).
Comparison of virologic failure between Arms.
PBMC, peripheral blood mononuclear cells.
HIV-1 DNA concentrations in PBMC
The HIV-1 DNA concentrations at baseline (n=32 subjects with available specimens) ranged from 0.49 to 3.3 log10 c/106 PBMCs. The median HIV-1 DNA concentration at baseline was similar for children whose plasma virus remained suppressed vs. children with virologic failure (2.42 vs. 2.24, p=0.72; Table 2). Overall, the HIV-1 DNA levels did not vary significantly between baseline and follow-up samples, with median values of 2.31, 2.09, 2.01, 2.12, and 2.09 log10 c/106 cells at 0, 24, 48, 72, and 96 weeks of ART, respectively. Furthermore, there was no difference in the viral DNA concentrations over time in the nonfailure vs. failure groups (p=0.82, Wei–Johnson nonparametric global test, Fig. 1A). Among children with virologic failure, the HIV-1 DNA concentration at the visit prior to plasma viral rebound was not higher than the concentration at the previous study visit (p=0.84, Fig. 1B). In addition, the median HIV-1 DNA concentration was similar at the visit before virologic failure and the time of virologic failure (1.96 vs. 2.09 log10 c/106 cells, p=0.99) and at virologic failure and the subsequent study visit (2.09 vs. 1.96 log10 c/106 cells, p=0.19).
Analysis of drug resistance mutations in longitudinal PBMC samples
Amino acid substitutions conferring drug resistance that require only one base change are spontaneously generated during viral replication, and, thus, these mutants are selected more rapidly compared to resistance requiring two or more base substitutions. Therefore, our analysis focused on the detection of single-base mutations that confer high-level resistance to the relevant study drugs NVP (K103N, Y181C, G190A) and 3TC (M184V). Because a high proportion of the children had received ZDV prior to enrollment in PACTG 377, we also screened for T215F/Y, which is associated with high-level resistance to both ZDV and the study drug d4T. Resistance to PI was uncommon in PACTG 377, with PI mutations rarely detected at the time of ART failure. 22 Therefore, we did not screen for mutations associated with resistance to NFV and RTV.
Drug resistance mutations were evaluated in specimens from 44 of the 46 children. One child did not have sufficient HIV-1 DNA copies for OLA analysis, and specimens from the other failed to amplify HIV-1 pol. Both of these children were in the nonfailure group (study Arm D).
Drug resistance mutations at study entry (baseline)
At baseline, the T215F/Y mutation was more prevalent in the nonfailure group compared to the failure group, 64% vs. 9% (p=0.004, Table 3). When detected at baseline, this mutation persisted at high concentrations throughout the follow-up period, except for decay in one child in the nonfailure group (data not shown). Mutations conferring resistance to NVP or 3TC were not detected in any of the children at baseline (n=30 evaluated at week 0). One child in the failure group had K103N detected in the week 12 specimen; however, as his baseline specimen was not available for analysis, we could not determine whether this mutation was present prior to the start of ART.
Baseline mutations include specimens from week 0, or week 12 (n=14) if baseline specimen was unavailable, as depleted in previous assays.
6/8, 6/8, and 9/17 subjects in Arms A, C, and D, respectively.
Subject #3 in Arm A.
Mutation detected at week 12 in Subject #4 without baseline specimen.
Mutations detected in Subject #19 at low concentrations and at one study visit only.
Only 10/11 children had PBMC specimens from the time of virologic failure and later.
p=0.004.
p<0.0001.
OLA, oligonucleotide ligation assay.
New drug resistance mutations detected during the study
After the initiation of ART, new resistance mutations were detected in one child (3%) in the nonfailure group. This child had NVP resistance mutations Y181C and G190A detected at low concentrations, and each mutation was detected at only one study visit (at weeks 84 and 36, respectively). In contrast, 100% of children in the failure group had new NVP resistance mutations detected (n=10 children with specimens from viral rebound and later visits, p<0.0001). The most frequent mutation detected was K103N (73%) followed by G190A (46%) and Y181C (36%). Only one of three Arm D children with virologic failure had M184V detected.
New resistance-mutations were detected in the PBMC before plasma viral rebound in two (22%), at the time of virologic failure in five (56%), and after virologic failure in two (22%) of nine children with a complete set of follow-up specimens for analysis (Table 4). The two subjects with NVP resistance mutants detected prior to failure had low concentrations of mutant present over 12 to 36 weeks prior to virologic failure, but no unique characteristics or behaviors were evident in these two compared to the other subjects with virologic failure. In addition to the nine reported above, one child (#8) had mutations detected at the time of virologic failure; however, a prefailure specimen was not available for resistance testing to determine whether the mutations were selected prior to failure. Another child (#11) did not have PBMCs available from the time of virologic failure or later. However, analysis of drug resistance in plasma RNA from the time of viral rebound and subsequent visit found high concentrations of K103N.
Baseline specimen not available, no history of NNRTI use.
Mutation detected at low concentrations in one study visit only.
Specimen from before plasma RNA rebound not available.
PBMCs from the time of virologic failure and later not available; K103N detected in plasma RNA at the time of viral rebound and the following visit.
NA, not available.
Transient virologic rebound and selection of drug resistance mutations
Among the 11 children in the failure group, plasma HIV-1 RNA ≥400 c/ml was sustained in six for the entire follow-up period, and five had either transient or intermittent viremias ≥400 c/ml. Drug-resistant mutants were selected in all five children with intermittent viremias, with mutant frequency and concentration similar to children with sustained virologic failure (Fig. 2). In this small subgroup, specific NVP mutations or the number of new mutations detected were not associated with either the magnitude of the viral load after failure or whether children had sustained or transient virologic failure.
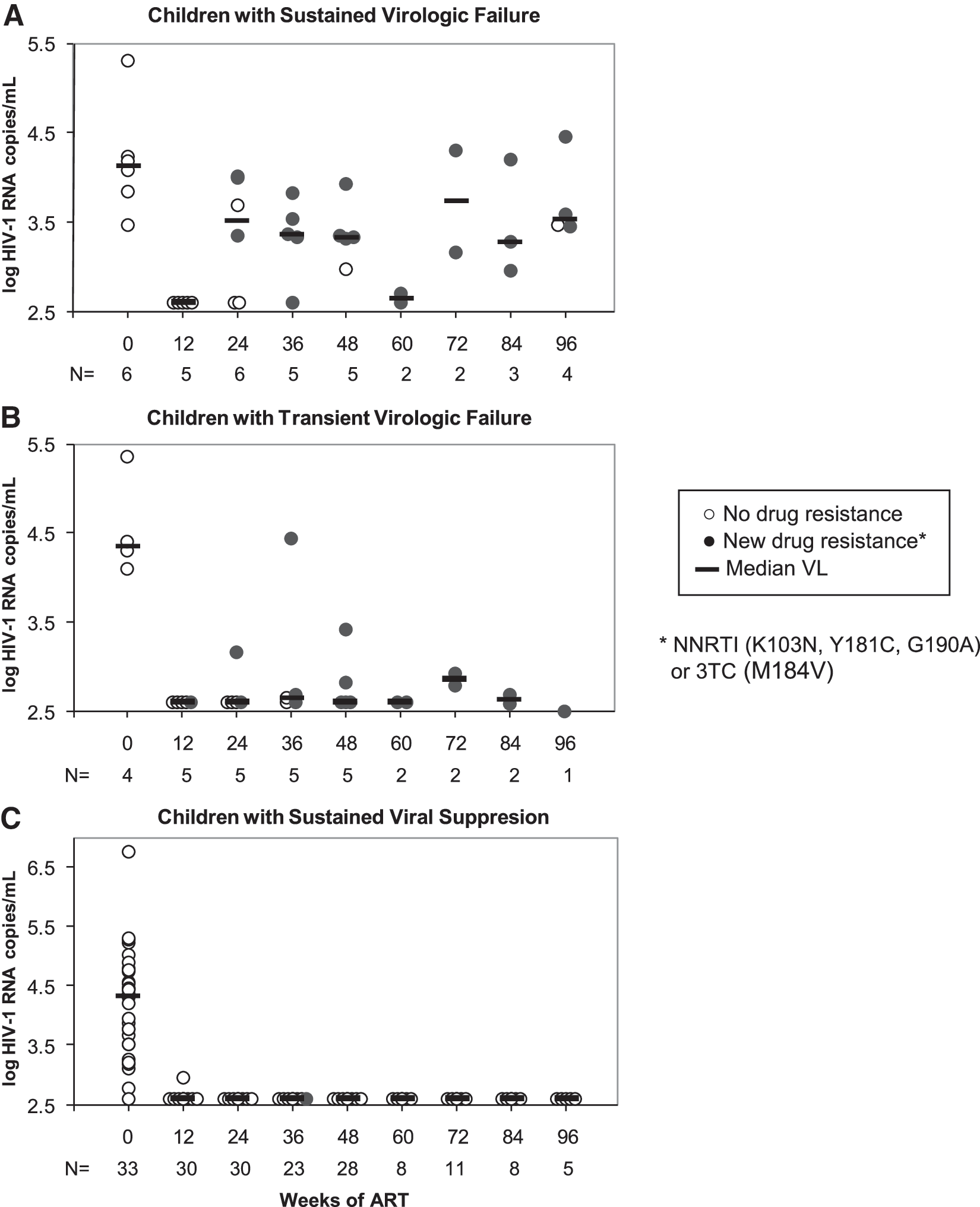
Virologic outcome of antiretroviral therapy (ART) and selection of HIV-1 drug resistance in children with initial suppression of plasma viral RNA. Plasma HIV-1 RNA is shown from initiation of ART through 96 weeks of follow-up for
Discussion
Our retrospective study found that monitoring of HIV-1 DNA concentrations or new onset of NVP and 3TC resistance mutations in PBMCs during effective ART did not identify individuals who subsequently experienced virologic failure of ART. New resistance mutations in PBMCs were detected in only 22% prior to rebound of plasma HIV-1 RNA (≥400 c/ml) and no consistent changes in HIV-1 DNA load were evident. Thus, neither parameter, when assessed quarterly, reliably predicted virologic failure in this cohort of subjects.
Multiple studies suggest that viral replication is insignificant when plasma HIV-1 RNA is <50 c/ml or intermittently detectable at concentrations <400 c/ml. 23 –25 However, our detection of mutants accumulating in the PBMC of a subset of subjects who later experienced virologic failure suggests replication was ongoing in spite of a plasma viral load <400 c/ml. This observation is consistent with our 12,16 and others' 13 –15 studies showing evidence of viral replication in select individuals when plasma viral loads were below the limit of detection.
The detection of resistance mutations prior to virologic failure suggests a gradual selection of drug-resistant variants. Gradual selection could result from low fitness of the mutant to ongoing ART with a high genetic barrier to resistance, or from occasional periods of time with marginally therapeutic ARV concentrations from intermittent nonadherence, inadequate doses of ARV, or variations in drug metabolism. In contrast, detection of mutant at or after the time of virologic failure suggests rapid onset of resistance. Subjects may have stopped taking their ART acutely with NVP resistance selected after the other drugs were excreted due to the long half-life of NVP. Once selected, NVP-resistant mutants may have quickly replicated to levels detectable in plasma. In this case, screening at 12 week intervals may have been too infrequent to capture the resistance before viral rebound. The recognition of two patterns of virologic failure confirms our previous observations in a different cohort 12 that a minority of individuals have evidence of viral replication during ART that suppresses plasma HIV-1 RNA to below the limit of detection.
An alternative explanation for the low detection of drug resistance in PBMCs prior to viral RNA rebound is that while the OLA detects mutant at ≥2% concentration in the viral population, and ∼150 HIV-1 copies per specimen were submitted to the assay, resistant mutants may have initially evolved at a level below the limit of detection of our assay. Further studies, with more sensitive assays and testing at more frequent intervals, may better characterize the subgroup that could benefit from monitoring, and may possibly lead to the development of strategies to prevent virologic failure.
NVP resistance was detected at some point in all subjects who experienced virologic failure, likely due to the long half-life of NVP with selection of mutants occurring after the other ARV were excreted. In contrast, detection of the 3TC resistance mutation M184V was infrequent. The Arm D ART regimen included four drugs, and the selection of M184V was likely precluded due to the longer half-life of NVP enhancing the genetic barrier to selection of 3TC-resistance.
The mutations encoding T215F/Y were frequently detected in the PBMCs of children at study entry (50%), consistent with previous exposure to ZDV. This mutation has been associated with high-level resistance to the study drug d4T; however, as noted previously, 22 its presence was not associated with virologic failure. The higher frequency of T215F/Y in subjects with sustained viral suppression compared to those with viral rebound (p=0.004), as previously suggested, 22 may indicate poor fitness and thus improved virologic response to multiclass ART.
Prior to sustained virologic failure several children had transient rebound of plasma HIV-1 RNA with selection of NVP resistance mutations. Mutations accumulated and persisted in the PBMCs even when the HIV-1 RNA levels in plasma returned to concentrations of <400 c/ml. These transient viremias appear to be the result of active viral replication and not the expression of nonreplicating archival virus, also known as “blips.” 16,26 The resuppression of plasma viral RNA to undetectable levels in spite of NVP resistance most likely occurred because the ART regimens used in PACTG 377 included three classes of ARV and therefore posed a relatively high genetic barrier to resistance.
Virologic rebound was less frequent in the four drug Arm D, 18 suggesting a relatively higher efficacy at maintaining viral suppression compared to the other three drug regimens. Arm C, which contained the less potent PI nelfinavir, had the highest failure rate; however, the difference in the rate of virologic failure among Arms A, C, and D was not statistically significant (p=0.24) in this substudy.
In this study, monitoring cellular HIV-1 DNA concentrations during ART failed to discriminate between children who maintained suppression of plasma virus and those who experienced virologic failure. The HIV-1 DNA levels measured in PBMCs showed great heterogeneity both at baseline and in response to ART. And changes in HIV-1 DNA concentrations between baseline and follow-up samples did not vary by treatment outcome. Our observations agree with studies of ART in adults that did not find associations between HIV-1 DNA concentrations and virologic failure, 27,28 or duration of viral suppression. 27,29 The small changes in HIV-1 DNA concentration from baseline to weeks 12, 24, 48, and 96 observed in this study were similar to previous reports in chronically infected children 30 and adults 27,28 receiving ART. In contrast, a separate study that followed 14 children on suppressive ART for up to 48 months reported a significant reduction of HIV-1 DNA by 48 and 96 weeks (0.75 and 0.9 log10 HIV-1 DNA c/106 cells). 31 In the latter study, more than 60% of children were therapy naive at the initiation of ART, in contrast to the children in our substudy of PACTG 377, which has been associated with a greater reduction of the HIV-1 DNA concentrations during ART. 8,28
Several studies have shown 28,31,32 that cell-associated HIV-1 DNA follows a two-phase decay after initiation of ART: an initial phase of short duration with a fast decay and a second slower phase thereafter with significant decay observed only after 1 or 2 years of therapy. The initial decay appears similar in naive and pretreated individuals, 28 in those with sustained suppression or with subsequent virologic failure, as observed in this and other studies, 27,28 and appears to correlate with suppression of plasma HIV-1 RNA to undetectable levels. 28,31,33 The second phase of HIV-1 DNA decay is thought to represent decay of long-lived cells, 6 which during suppressive ART show little variation over time, with a half-life of months to years. 9,10,34 Two factors thought to contribute to the stability of this reservoir are the intrinsic stability of resting memory CD4+ T cells and replenishment by low-level viral replication. Evolution of drug resistance mutations during suppressive ART 12 –16 suggests low-level viral replication; however, replenishment by residual replication has been estimated to contribute minimally to the persistence of the reservoir. 35 Growing evidence suggests that latently infected cells proliferate, 16,26 which may represent a third process that replenishes the reservoir during ART. 36,37 In this context, it seems unlikely that measurements of cellular HIV-1 DNA would identify individuals who will fail ART. Direct measurement of viral replication parameters such as cell-associated HIV-1 RNA 31,38,39 or episomal circularized HIV-1 DNA with two long-terminal repeats (2-LTR) 40,41 may provide a better prognostic tool; these tests, however, have not been standardized or validated for use in the clinical setting.
In summary, the use of sensitive assays to monitor the concentration of HIV-1 DNA and the new onset of drug resistance mutations in PBMCs of children undergoing suppressive ART did not reliably predict virologic failure. High variability in cellular HIV-1 DNA loads and a relatively small decline during the first year of ART suggest that measurements of HIV-1 DNA are not suitable to gauge low levels of ongoing viral replication. Monitoring for new drug resistance mutations in PBMCs may identify impending virologic failure in a subset of individuals experiencing low-level viral replication that results in gradual selection of mutants. Unfortunately, capturing resistance prior to viral rebound in individuals with rapid selection of resistance would require frequent screening, which would not be practical unless sensitive, inexpensive, and simple tests are developed.
Footnotes
Acknowledgments
This work was supported by 1 U01 AI068632 awarded (L.M.F.) for a Developmental Virology Laboratory, and a Society for Pediatric Research awards (M.J. and J.H.). Overall support for the International Maternal Pediatric Adolescent AIDS Clinical Trials Group (IMPAACT) was provided by the National Institute of Allergy and Infectious Diseases (NIAID) [U01 AI068632], the Eunice Kennedy Shriver National Institute of Child Health and Human Development (NICHD), and the National Institute of Mental Health (NIMH) [AI068632]. The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIH. This work was supported by the Statistical and Data Analysis Center at Harvard School of Public Health, under the National Institute of Allergy and Infectious Diseases cooperative agreement #5 U01 AI41110 with the Pediatric AIDS Clinical Trials Group (PACTG) and #1 U01 AI068616 with the IMPAACT Group. Support of the sites was provided by the National Institute of Allergy and Infectious Diseases (NIAID) and the NICHD International and Domestic Pediatric and Maternal HIV Clinical Trials Network funded by NICHD (contract number N01-DK-9-001/HHSN267200800001C). We thank James Ham for his valuable contribution to this work.
Author Disclosure Statement
No competing financial interests exist.