
Abstract
Accessory and regulatory proteins (nonstructural proteins) have received increasing attention as components in novel HIV/SIV vaccine design. However, the complicated interactions between nonstructural proteins and structural proteins remain poorly understood, especially their effects on immunogenicity. In this study, the immunogenicity of structural proteins in the presence and absence of nonstructural proteins was compared. First, a series of recombinant plasmids and adenoviral vectors carrying various SIVmac239 nonstructural and structural genes was constructed. Then mice were primed with DNA plasmids and boosted with corresponding Ad5 vectors of different combinations, and the resulting immune responses were measured. Our results demonstrated that when the individual Gag, Pol, or Env gene products were coimmunized with the whole repertoire of nonstructural proteins, the Gag-specific CD8+ T response was greatly enhanced, while the Env- and Pol-specific CD8+ T responses were significantly reduced. The same pattern was not observed in CD4+ T cell responses. Antibody responses against both the Gag and Env proteins were elicited more effectively when these structural antigens were immunized together with nonstructural antigens. These findings may provide helpful insights into the development of novel HIV/SIV vaccines.
Introduction
T
In addition to structural proteins (Gag, Pol, and Env), which have traditionally been the major targets in HIV/SIV vaccine design, the genome of HIV/SIV contains genes encoding regulatory (Tat and Rev) and accessory proteins (Nef, Vif, Vpr, and Vpu/Vpx). Tat and Rev are essential for viral replication. 19 –21 Because the mRNAs of Tat, Rev, and Nef are fully spliced and transported from the nucleus to the cytoplasm, these are the first proteins expressed during the life cycle of HIV/SIV. Although Nef, Vif, Vpr, Vpx and Vpu are dispensable for viral growth, 22,23 these accessory proteins are necessary for viral pathogenesis and play very important roles in maintaining high levels of viral replication and carrying out other functions. 24 Given the importance of regulatory and accessory proteins in viral survival, these proteins are promising targets for vaccines against HIV/SIV. Moreover, by screening HIV-1 patients with overlapping viral peptide pools, it has been shown that all regulatory and accessory proteins serve as targets for cytotoxic T lymphocytes (CTL). 25 –31 Therefore, to broaden T cell responses, increasing numbers of nonstructural proteins are being included as components of HIV/SIV vaccines. 16,18,32 –38 However, it is still poorly understood how interactions between structural and nonstructural HIV/SIV proteins influence immunogenicity. Therefore, in the present work, we performed a comprehensive study to clarify how entire nonstructural proteins affect the immunogenicity of Gag, Env, or Pol.
Materials and Methods
Preparation of the SIV vaccine regimen
The full-length genes encoding the structure proteins of SIVmac239 virus (Gag, Pol, and Env) were cloned into pVax1 expression vectors (Invitrogen, USA) and replication-incompetent adenovirus type 5 vectors (Ad5-SIVgag, Ad5-SIVpol, and Ad5-SIVenv) individually as previously described. 36 The sequences of regulatory and accessory proteins (Gag, Pol, Env, Nef, Rev, Tat, Vif, Vpr, and Vpx) were derived from SIVmac239 and optimized in accordance with the preferred codon usage of mammalian cells, which has also been previously described. In addition, Flag protein tags were attached to the C-termini of Nef, Rev, Tat, Vif, Vpr, and Vpx, which were then cloned into pVax1 expression vectors (Invitrogen, USA) (generating pVax1-Nef, pVax1-Rev, pVax1-Tat, pVax1-Vif, pVax1-Vpr, and pVax1-Vpx, respectively). The regulatory and accessory proteins were divided into two groups to construct two fusion genes using the overlapping polymerase chain reaction (PCR) method in series: VRN (Vpr-Rev-Nef) and VVT (Vif-Vpx-Tat). A Furin cleavage site and an effective FMDV 2A self-cleavage sequence were synthesized together and inserted between Vpr, Rev, and Nef in VRN and Vif, Vpx, and Tat in VVT. The sequences of Furin and 2A are RAKR and APVKQTLNFDLLKLAGDVESNPGP, respectively, as previously described. 39,40 The amplified fusion genes were cloned into separate pVax1 vectors (to generate pVax1-VRN and pVax1-VVT) and replication-incompetent adenovirus type 5 vectors (to generate Ad5-VRN and Ad5-VVT). Recombinant adenoviruses were generated by homologous recombination methods, as previously published. 41 All of the constructs of the plasmids and adenoviruses were confirmed by direct sequencing.
Western blot
To analyze the expression and cleavage characteristics of the fusion proteins, HEK 293 cells were transfected with 24 μg of the pVax1-nef, pVax1-Tat, pVax1-VRN, and pVax1-VVT plasmids using 60 μl of Lipofectamine 2000 (Invitrogen, USA) in 10-cm dishes. After 24 to 48 h, the cells were lysed in lysis buffer (50 mM Tris-Cl, pH 7.4,150 mM NaCl, 1 mM EDTA, 1% Triton X-100) with PMSF. In addition, HEK 293 cells were infected with purified Ad5-VRN, Ad5-VVT, or Ad5-empty virions. After 3 days, the cells were collected and lysed by freezing and thawing. All of the lysates were loaded onto sodium dodecyl sulfate (SDS)-polyacrylamide gels and separated. Then proteins were transferred to PVDF membranes and blocked with 5% nonfat milk in phosphate-buffered saline (PBS) overnight at 4°C. After washing with PBS/PBST, membranes were incubated with primary antibodies at room temperature for 1 h. Anti-Flag (Sigma-Aldrich, USA) and anti-Nef (Abcom Ltd., Hong Kong) monoclonal antibodies were diluted 1:1000 with PBST, and the anti-Tat (Santa Cruz, Inc., USA) polyclonal antibody was used at a dilution of 1:2500. Membranes were then washed and incubated with horseradish peroxidase-conjugated secondary antibodies for 1 h. After extensive washes, membranes were incubated with Chemiluminescent HRP substrate (Millipore Corporation, Billerrica, MA) for 3 min before being exposed to autoradiography film.
Fluorescence microscopy
HeLa cells were seeded onto glass slides and transfected with pVax1-Nef, pVax1-Tat, pVax1-VRN, or pVax1-VVT expression plasmids using Lipofectamine 2000 transfection reagents. After 24 to 48 h, the cells were washed with PBS and then fixed with 4% paraformaldehyde in PBS for 15 min at 4°C. The cells were permeabilized with 0.05% Triton X-100 in PBS for 5 min and blocked with 5% bovine serum albumin in PBS for 1 h. Then the cells were incubated with a 1:50 dilution of either anti-Flag antibody, anti-Nef antibody, or anti-Tat antibody in blocking buffer for 1 h at room temperature, followed by a 1:100 dilution of appropriate Cy3-conjugated secondary antibodies for 30 min. The cells were then stained with DAPI (4'-6-diamidino-2-phenylindole). Lastly, images of slides were observed and photographed using an immunofluorescence microscope (LEICA DMI6000B).
Animals and immunizations
The animals used in this study were 5-week-old female C57BL/6 mice, which were bred at the Experimental Animals Center of the Guangzhou Institute of Biomedicine and Health (GIBH, Guangzhou, China) in compliance with the appropriate institutional regulatory committees. All of the animal experiments in this study were approved by the GIBH Institutional Animal Care and Use Committee (IACUC).
The mice were divided into eight groups of five mice, as shown in Table 1, and the group immunized with pVax1-empty and Ad5-empty vector was used as a control. The quadriceps of the mice were first injected 24 h before each DNA inoculation with a total of 100 μl of 0.5% procaine hydrochloride as a pretreatment. Then at 0, 2, and 4 weeks, each mouse received 50 μg of the appropriate plasmids dissolved in PBS, injected intramuscularly into the same quadriceps as the procaine injection. At 7 weeks, the mice were boosted with 1×1010 vp (viral particles) of the corresponding Ad5 vectors. All of the mice in each group were sacrificed at week 9, and splenocytes were isolated for ELISPOT, intracellular cytokine staining, and CFSE-based T cell proliferation assays.
SIV peptide pools
As published previously, 36 SIVmac239-specific peptide pools in this study were provided by the NIH AIDS Research and Reference Reagent Program. Peptides in this pool were 15 amino acids in length and overlapped by 11 amino acids; together, they spanned the entire expressed genome of SIVmac239. All of the peptides were dissolved at 0.4 mg/ml in DMSO and included in nine different peptide matrix systems: Gag, Pol, Env, Nef, Rev, Tat, Vif, Vpr, and Vpx.
Interferon-γ ELISPOT assays
The gamma interferon (IFN-γ) ELISPOT assays were conducted according to previously described methods.
36,41
Freshly isolated splenocytes were used for the detection of IFN-γ-secreting cells. Ninety-six-well plates containing Immobilon-P membrane (Millipore, USA) were coated with antimouse IFN-γ monoclonal antibody (BD Pharmingen) overnight at 4°C and blocked with R10 medium (RPMI 1640, 0.05 mM 2-mercaptoethanol, 1 mM sodium pyruvate, 2 mM
Intracellular cytokine staining (ICS)
As previously described, 36,41 2×106 splenocytes were obtained from the mice and stimulated with either the SIV-specific peptides or DMSO as a control for 1 h at 37°C. After adding brefeldin A (BD Biosciences), the splenocytes were incubated for an additional 16 h at 37°C. The cells were then washed and stained with APC-conjugated anti-CD3, PerCP-conjugated anti-CD8, and FITC-conjugated anti-CD4 antibodies (BD Pharmingen) for 1 h, suspended in 250 μl of Cytofix/Cytoperm (BD Pharmingen) solution for 20 min, washed with Perm/Wash solution (BD Pharmingen), intracellularly stained with PE-conjugated antimouse IFN-γ antibody (BD Pharmingen) for 30 min, and analyzed by flow cytometry (BD FACS Calibur platform). The results of unstimulated samples were considered to be background and were subtracted from the experimental samples.
Ex vivo T lymphocyte proliferation assay by CFSE staining
Splenocytes were suspended at 1×106 cells/ml and mixed with 0.25 mM carboxyfluorescein succinimidyl ester (CFSE; Molecular Probes). After a 10-min incubation at 37°C, ice-cold RPMI containing 10% fetal bovine serum (FBS) was added to the mixture to stop staining. Cells were then washed with cold 10% FBS/RPMI solution, resuspended in complete RPMI media, and stimulated with or without the SIV-specific peptide pool (2 μg/ml) using SEB as a positive control. Six days later, the cells were collected, washed with PBS, stained for 30 min with APC-conjugated anti-CD8 and PerCP-Cy5.5-conjugated anti-CD3 antibodies (BD Biosciences), and then washed twice before flow cytometric analysis (BD FACS Calibur platform). The results of unstimulated samples were considered to be background and were subtracted from the experimental samples that responded to SIV-specific antigen.
ELISA assay for SIV-binding antibodies
An enzyme-linked immunosorbent assay (ELISA) to detect antibodies specific to SIVmac239 was performed as previously described. 36,42 The 96-well plates were coated with 100 μl of whole, disrupted SIVmac239 viruses at 1 mg/ml in 0.1 M sodium carbonate buffer (pH 9.6). The plates were incubated overnight at 4°C, and the wells were then blocked with 200 μl of 5% nonfat powdered milk in PBST for 1 h at 37°C. Serum samples were serially diluted (1:2), and 100 μl was added to the wells in duplicate. After a 1 h incubation at 37°C, antibody binding was detected by incubation with a 1:5000 dilution of horseradish peroxidase (HRP)-conjugated goat antimouse antibody (100 μl per well) for 1 h at 37°C. The wells were developed by adding 100 μl of TMB substrate (Chem-icon, Temecula, USA) for 5 min, and the reaction was stopped with 100 μl of 0.5 M H2SO4 before the OD at 450 nm was read.
Data analysis
Flow cytometric data were analyzed using WinMDI v2.9 software (Scripps Research Institute). Statistical analyses were computed with JMP version 6.0.3 (SAS Institute Inc.). A Student's t-test was used for comparison of immune responses between different groups, and a p-value of less than 0.05 was considered significant. Graphic presentations were drawn using Excel 2003.
Results
Construction and expression of the VRN (Vpr-Rev-Nef) and VVT (Vif-Vpx-Tat) fusion constructs with Furin-2A proteolytic cleavage sites
The SIVmac239 accessory and regulatory genes were first optimized to enhance their level of expression, in accordance with codons frequently used in mammalian cells, 36 and then cloned into the pVax1 expression vector with individual Flag protein tags (pVax1-Nef, pVax1-Rev, pVax1-Tat, pVax1-Vpr, pVax1-Vif, and pVax1-Vpx). All of these plasmids were then used as the foundation of the multigene DNA expression vector. As shown in the schematic diagrams in Fig. 1A and D, the sequences of Nef, Rev, and Vpr and the sequences of Tat, Vif, and Vpx were fused in two frames in series, referred to as VRN (Vpr-Rev-Nef) and VVT (Vif-Vpx-Tat), respectively. Meanwhile, the tags of all six proteins and the stop codons of Rev, Vpr, Vif, and Vpx were removed, and FMDV 2A self-cleavage sites were added following the coding regions of Rev, Vpr, Vif, and Vpx. Because the 2A self-processing cleavage occurred between the last two amino acids at the C terminus of the 2A peptide, the anterior protein in the cassette had 23 additional amino acid residues at its C terminus. To maintain the natural structure of the anterior protein in frame and to eliminate possible adverse effects caused by the remaining 2A residues, Furin cleavage site sequences (RAKR) were inserted in front of the 2A sequence as previously described. 39,40 Then the fragments of VRN and VVT were cloned into the pVax1 and Ad5-based vectors.
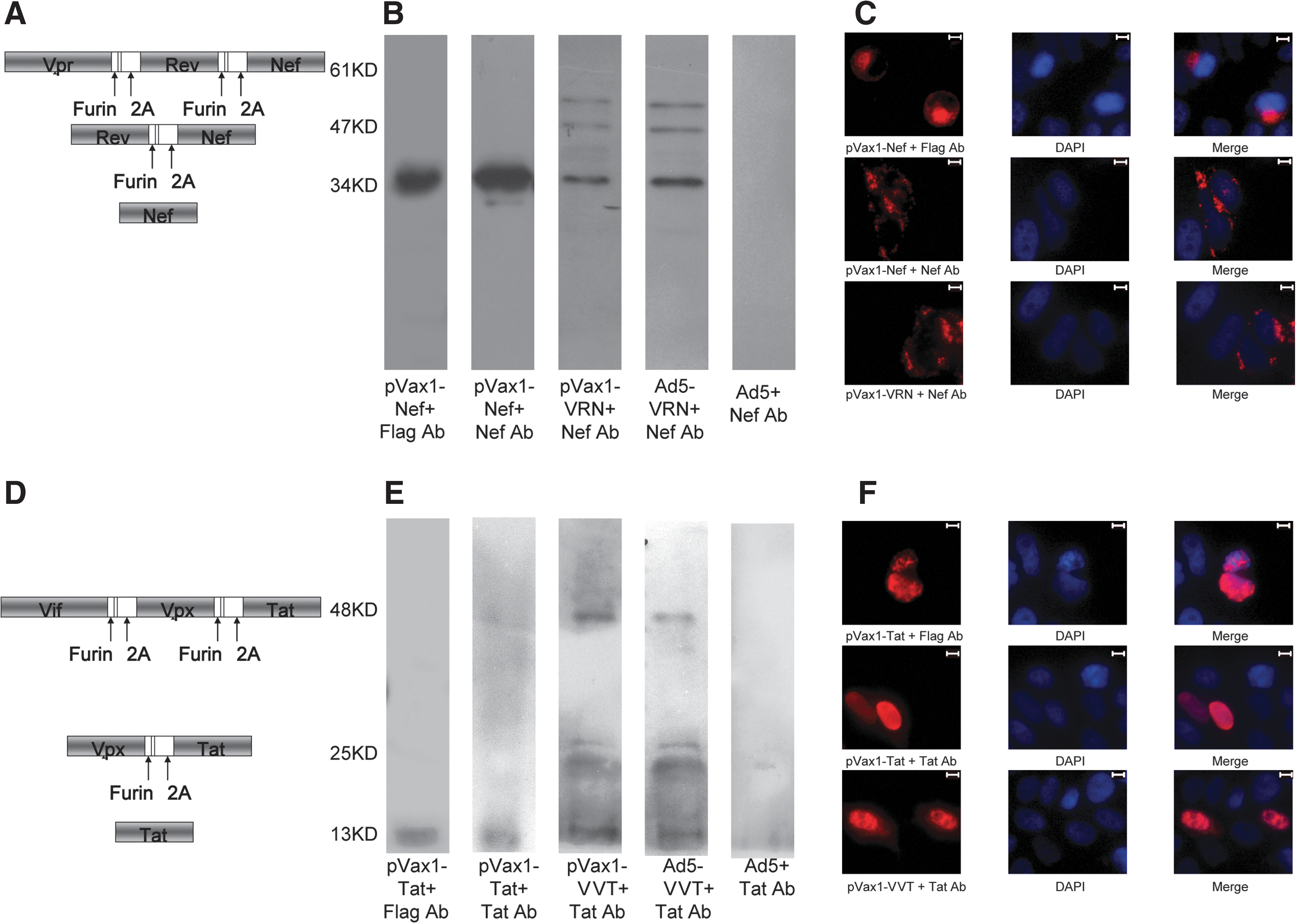
Construction, expression, and subcellular localization of different expression cassettes.
Because the only commercially available antibodies against the accessory and regulatory proteins of SIV were Nef and Tat specific, these alignments (Vpr-Rev-Nef, Vif-Vpx-Tat) could be used to detect the expression and cleavage of VRN and VVT. HEK293 cells were either transfected with plasmids of pVax1-Nef, pVax1-Tat, pVax1-VRN, or pVax1-VVT or infected with Ad5-VRN or Ad5- VVT viruses. As shown in Fig. 1B and E, cell lysates were incubated with the corresponding antibody against Flag, Nef, or Tat and subjected to Western blot analysis. Transfection of pVax1-Nef and pVax1-Tat resulted in a 34-kDa Nef protein and 13-kDa Tat protein, recognized by the anti-Flag antibody and the anti-Nef or anti-Tat antibodies, respectively. Transfection of pVax1-VRN and infection with Ad5-VRN resulted in three protein bands with molecular weights of 34, 47, and 61 kDa. Similarly, transfection of pVax1-VVT and infection with Ad5-VVT resulted in three bands with molecular weights of 13, 25, and 48 kDa. The schematic diagrams in Fig. 1A and D indicate the cleavage pattern of VRN and VVT and explain the formation of three different protein bands in each set. Because the anti-Nef and the anti-Tat antibodies detect only distinct epitopes of Nef or Tat, respectively, the 34-, 47-, and 61-kDa bands of VRN corresponded to the Nef, Rev-Nef, and Vpr-Rev-Nef proteins, respectively, while the 13-, 25-, and 48-kDa bands of VVT corresponded to the Tat, Vpx-Tat, and Vif-Vpx-Tat proteins, respectively. As expected, infection with the Ad5-empty virus resulted in no recognized bands.
Subcellular localization of the VRN and VVT fusion proteins in HeLa cells
To examine whether the subcellular localization of separate proteins derived from the fusion proteins would differ from that of the original proteins, we transfected HeLa cells with the pVax1-Nef, pVax1-Tat, pVax1-VRN, and pVax1-VVT plasmids and performed immunofluorescence studies. Because of antibody limitations, we assessed the localization of only Tat and Nef. Transfection with pVax1-Nef and pVax1-Tat, which could be detected by anti-Flag and anti-Nef/Tat antibodies, showed that Tat was mainly localized to the nuclei and Nef was mainly localized to the cytoplasm, consistent with previous reports 43 (Fig. 1C and F). We then compared the localization of the Nef and Tat proteins derived from pVax1-Nef and pVax-Tat, respectively, and the fusion proteins derived from pVax-VRN/VVT. The results indicated that fusing the genes did not alter the subcellular distribution patterns of their products.
Antigen-specific cellular immune responses elicited by different antigen regimens
An IFN-γ ELISPOT assay was used to assess SIV-specific cellular immune responses against SIV Gag, Pol, Env, Nef, Vif, Vpx, Vpr, Rev, and Tat peptide pools. Interestingly, as shown in Fig. 2, the level of the immune response against Gag was significantly higher following immunization with Gag+VRN and VVT compared to the response induced by immunization with Gag alone (p=0.035, Fig. 2A, and the SFCs were increased by 133%). In contrast, the immune responses against Pol and Env were significantly lower when Pol (p=0.013 for Pol1, and p=0.018 for Pol2, Fig. 2B; the SFCs were decreased by 77% and 13%, respectively) or Env (p=0.019 for Env1, and p=0.174 for Env2, Fig. 2C; the SFCs were decreased by 59% and 67%, respectively) were coadministered with VRN and VVT.
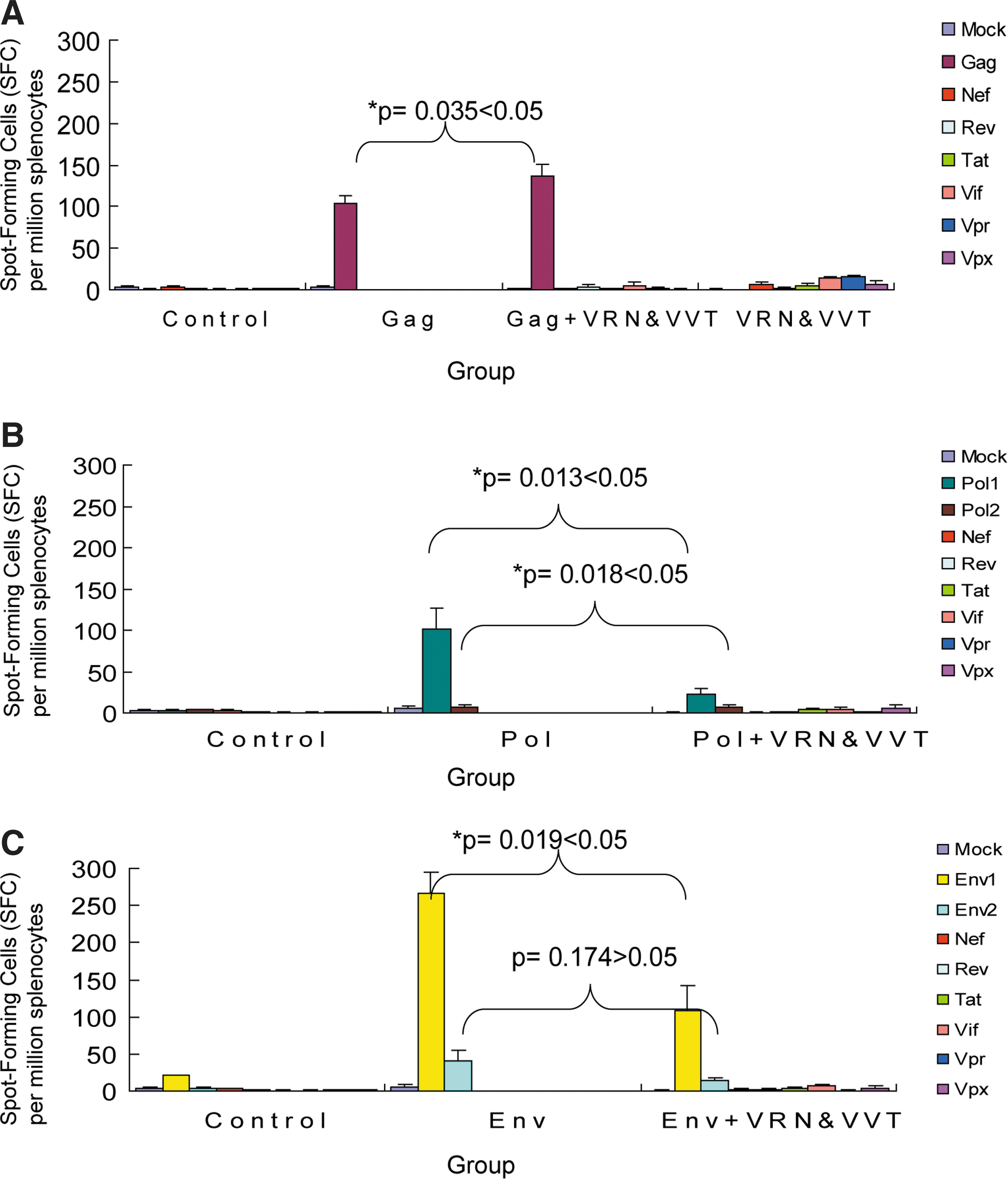
SIV-specific cellular responses in mice detected by interferon (IFN)-γ ELISPOT assays. SIV-specific cellular immune responses were assessed by IFN-γ ELISPOT assays following stimulation with peptide pools. The group immunized with pVax1-empty and Ad5-empty vectors was used as a negative control. Cellular responses were compared between
To confirm these results, an ICS (intracellular cytokine staining) assay was performed to assess SIV-specific cellular responses. As shown in Fig. 3, both CD4+ and CD8+ IFN-γ secreting T cell populations were evaluated. Consistent with the ELISPOT results, the level of the response against the accessory and regulatory proteins was still lower than the response against the structural gene products (Gag, Pol, Env), and it declined in all three coadministration groups (Gag+VRN and VVT, Pol+VRN and VVT, and Env+VRN and VVT). The frequency of CD8+ T cells that produced IFN-γ was much greater than the frequency of CD4+ T cells that produced IFN-γ, suggesting that the immunization may elicit a more intense CD8+ T cell response. As demonstrated in Fig. 3B and C, the group that received coadministration of Gag and VRN and VVT exhibited more IFN-γ-secreting CD8+ T cells than the group that received a single administration of Gag (increased by 130%), which was consistent with the ELISPOT results. The number of Pol-specific and Env-specific IFN-γ-secreting CD8+ T cells was lower in the coadministration groups (Pol+VRN and VVT and Env+VRN and VVT group) than in the single administration groups (Pol group and Gag group). In contrast, the number of IFN-γ-secreting CD4+ T cells was not reduced (Fig. 3D–G).
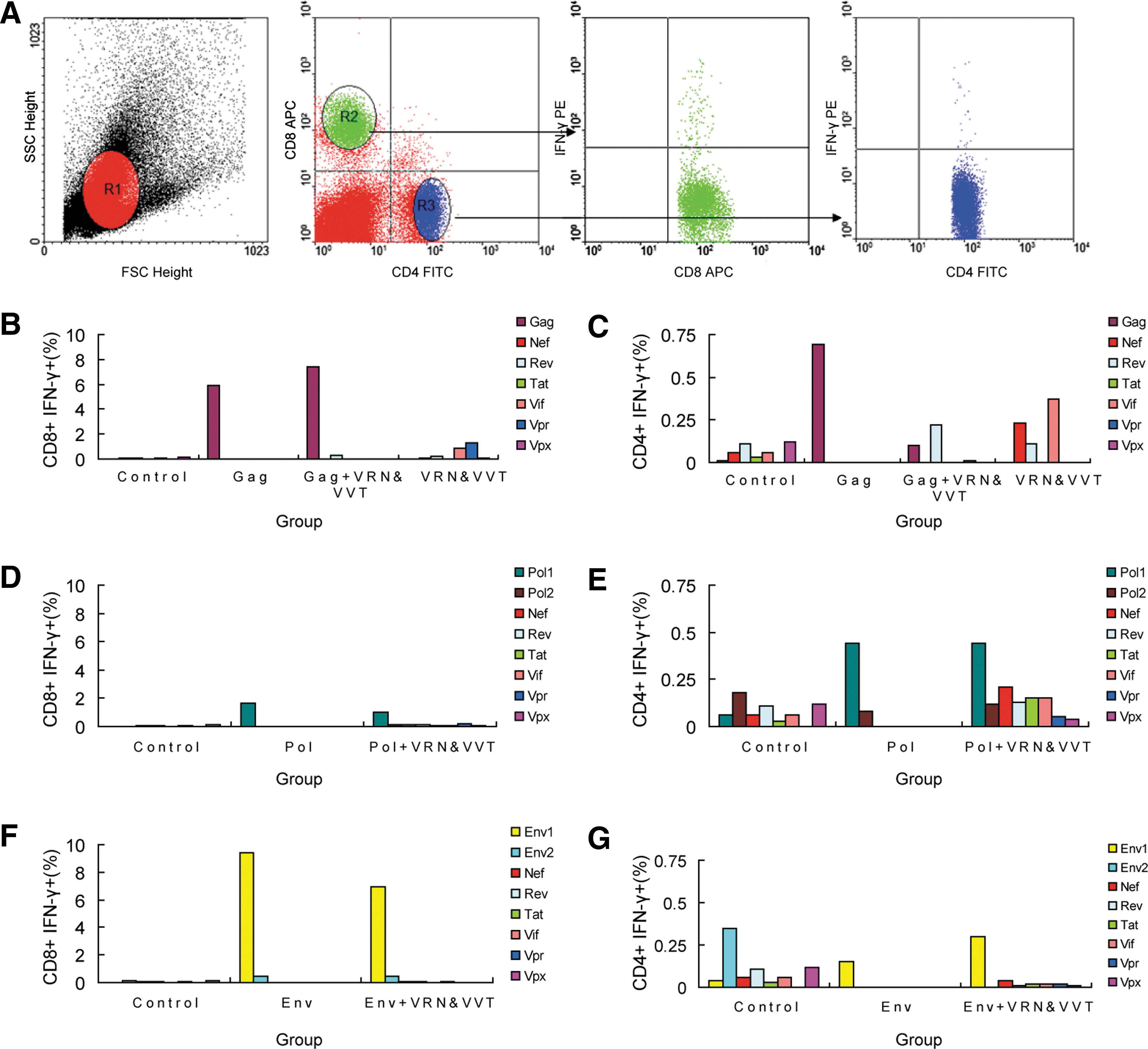
SIV-specific CD4+ and CD8+ T cell responses in mice detected by intracellular cytokine staining (ICS) assays.
SIV-specific CD4+ and CD8+ T cell proliferation assay by CFSE staining
In accordance with previously described methods, 36 CD4+ and CD8+ T cell proliferation induced by SIV peptide pools was assessed by CFSE staining. Consistent with the cellular responses described above and shown in Fig. 4B and C, CD8+ T cell proliferative responses in the Gag+VRN and VVT immunization group were enhanced relative to those from the Gag alone group. In contrast, the opposite trend was observed for CD4+ T cell proliferative responses. In addition, the proliferating populations of both CD8+ and CD4+ T cells from the Pol+VRN and VVT and Env+VRN and VVT immunization groups were decreased relative to those from the Pol and Env immunization groups, as shown in Fig. 4D–G.

SIV-specific CD4+ and CD8+ T cell proliferation detected with CFSE staining.
Detection of SIV-specific humoral responses against different immunogens
We next evaluated humoral immunity elicited by these immunization regimens. As shown in Fig. 5, except for samples from groups immunized with Gag, Gag+VRN and VVT, Env, and Env+VRN and VVT, other groups showed low levels of SIV-specific antibodies, mostly because the genes in these other groups are usually expressed as intracellular and nonstructural proteins contained within the SIV virus and are not expressed at the same levels as the structural proteins, Gag and Env. The highest titers of SIV-specific antibodies were found in the group immunized with the Env+VRN and VVT antigen regimen. Most interestingly, both Gag- and Env-specific antibody responses were elicited more effectively when these structural antigens were immunized together with nonstructural antigens.
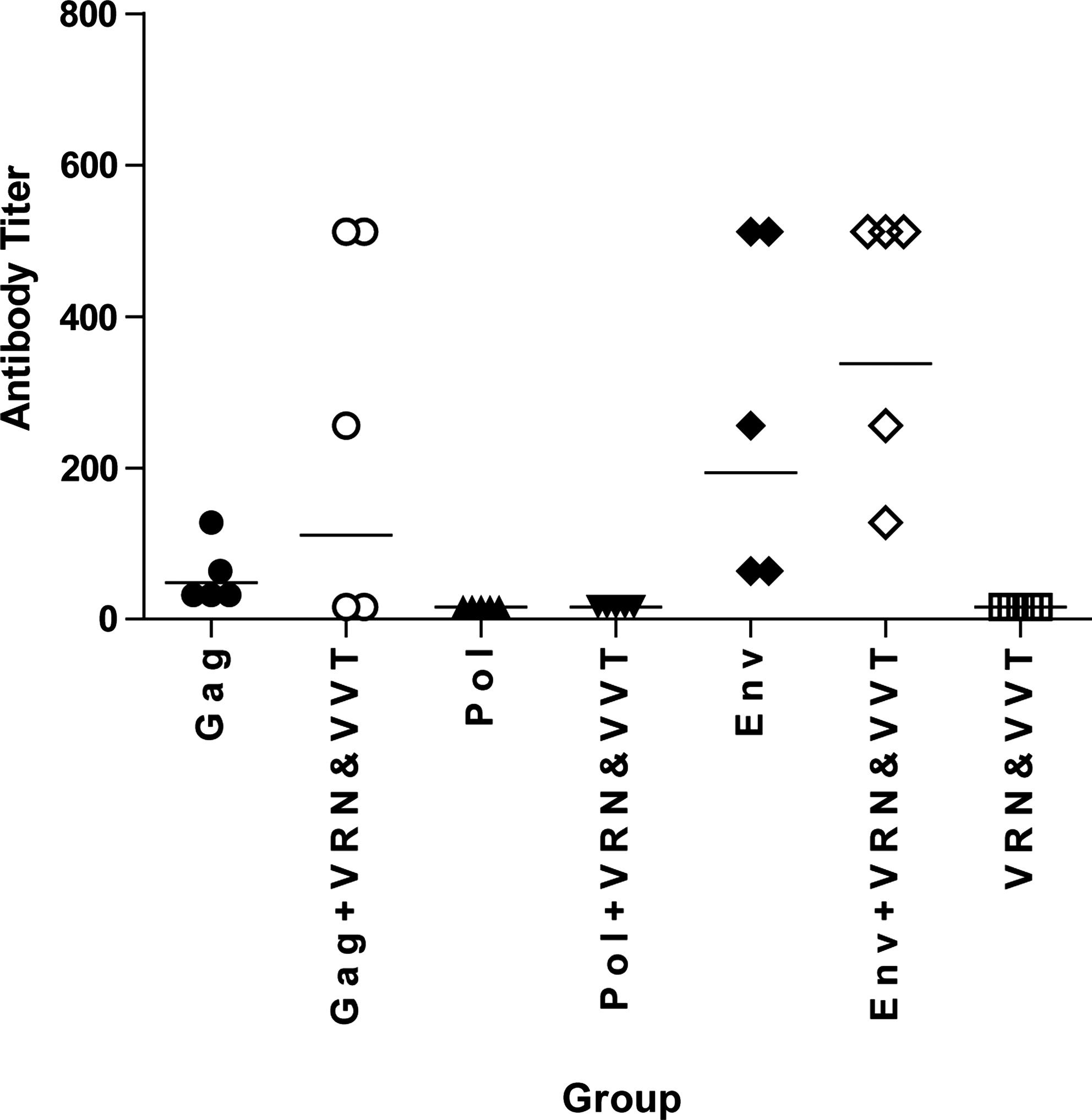
Vaccination-elicited antibody responses in mice. The SIV-binding antibody titers of Gag, Gag + VRN and VVT, Pol, Pol + VRN and VVT, Env, Env + VRN and VVT, and VRN and VVT groups were assessed. Each data point represents the antibody titer from an individual mouse. Lines represent the geometric mean titer for each group.
Discussion
Traditionally, the structural proteins, Gag, Pol, and Env, have been the major targets in HIV/SIV vaccine studies. Recently, accessory and regulatory proteins have received increasing attention as components in novel HIV/SIV vaccine design. However, the potential effects of the inclusion of nonstructural proteins on overall vaccine immunogenicity are still not clear.
Immune modulations between different SIV genes have been reported previously; for example, vaccinating monkeys with SIV Env/Rev and Gag together enhances the cellular immune response to Env significantly; vaccination with SIV Env/Rev, Nef, and Gag together reduces the immune response to Nef 44 ; and SIV Tat is known to enhance and broaden cellular immune responses to SIV Env and Gag. 45,46 Therefore, complicated interactions have resulted when increasing numbers of nonstructural proteins have been included as components of HIV/SIV vaccines. Recently, a vaccine regimen in which all accessory and regulatory proteins were coimmunized with Gag and Pol enhanced the breadth and frequencies of T cell responses and efficiently controlled the virus. 18 Although this study detected the immunogenicity of various SIV antigens, including structural and nonstructural proteins, the interactions between these various antigens were not assessed. Herein, we report a comprehensive study to clarify how the entire nonstructural proteins affect the immunogenicity of Gag, Env, and Pol.
Due to antigenic competition 37 and the negative effects of accessory and regulatory proteins on the immune response, 47 –49 it may be hypothesized that the immunogenicity of the structural antigens will be reduced if they are immunized together with nonstructural antigens. As expected, in this study, the cellular immune responses against Pol and Env were dramatically reduced when these proteins were coimmunized with accessory and regulatory proteins. Surprisingly, however, we observed, for the first time, that the IFN-γ response against Gag, as detected by ELISPOT, was enhanced when it was immunized together with nonstructural antigens. To confirm this result, an ICS assay was performed, which further showed that coimmunization enhanced the Gag-specific CD8+ T cell response but did not significantly change the Gag-specific CD4+ T cell response. Moreover, this observation was supported by the results of CFSE-based T cell proliferation assays. Specifically, the SIV-specific proliferative responses of CD8+ T cells in the Gag+VRN and VVT immunization group were enhanced relative to those from the Gag alone immunization group. This result suggests that a greater quantity of Gag-specific CD8+ T cells is produced because of their enhanced proliferation and that this higher proportion of Gag-specific IFN-γ-secreting CD8+ T cell accounts for the enhanced Gag-specific cellular response.
Although the specific mechanisms require further investigation, the findings in this study may help to explain certain phenomena, such as the disparate immunogenicity of various viral proteins during natural HIV/SIV infection. The fact that regulatory and accessory proteins can enhance CD8+ T cell responses against Gag and reduce the responses against Env and Pol may account for the disproportionately high level of the cellular response against Gag compared to Pol and Env, even though the Pol and Env proteins are longer and possess a greater number of recognizable epitopes than the Gag protein. 27
Overall, we described a novel strategy to construct two fragments that can express entire accessory and regulatory SIV proteins with optimized codons. Moreover, we examined how accessory and regulatory proteins affect the immunogenicity of structural proteins when used as components of vaccines. When these structural antigens were immunized together with nonstructural antigens, Gag-specific CD8+ T cell responses were enhanced, whereas the Pol- and Env-specific responses were reduced, and antibody responses against both Gag and Env proteins were elicited more effectively. In sum, this study may provide useful guidance for designing novel HIV/SIV vaccine regimens that include nonstructural proteins as a component.
Footnotes
Acknowledgments
We gratefully acknowledge Mr. Yichu Liu and Mr. Maochao Zhang for their technical support. This study was supported by the National Natural Science Foundation of China (81000737), Guangdong Natural Science Foundation (S2011010004564), the Knowledge Innovation Program of the Chinese Academy of Sciences (KSCX1-YW-10, KSCX2-EW-J-27), the National Key Science & Technology Specific Projects of China (2008ZX10001-011), and the National Science Fund for Distinguished Young Scholars of China (30688004).
Author Disclosure Statement
No competing financial interests exist.