
Abstract
Understanding factors that affect heterosexual transmission of HIV in women is of great importance. Lactobacilli in the lower genital tract of women utilize glycogen in vaginal epithelial cells as an energy source and produce lactic acid. The resultant vaginal acidity is believed to provide protection against HIV infection. Conversely, bacterial vaginosis (BV) is characterized by less lactic acid and a higher pH, and is associated with increased susceptibility to HIV infection. Because vaginal infection of macaques with simian immunodeficiency virus (SIV) or simian-human immunodeficiency virus (SHIV) is used as a model to study HIV sexual transmission, and because previous studies have shown a paucity of lactobacilli in rhesus macaques' lower genital tract, we compared lactic acid and glycogen levels in the genital fluid of rhesus and pigtail macaques with levels found in humans. The levels of lactic acid were lower in both rhesus (median=1.2 mol lactate/mg protein) and pigtail macaques (median=0.7 mol/mg) compared to women with healthy genital microbiota (median=4.2 mol/mg). Glycogen levels were significantly lower in both rhesus (median=0.004 μg glycogen/μg protein) and pigtail macaques (median=0 μg/μg) than in women (median=0.2 μg/μg). No significant differences in glycogen or lactate levels were observed comparing longitudinally collected samples from cycling pigtail macaques. These data show that the previously reported scarcity of lactobacilli in macaques correlates with low glycogen and lactic acid levels. These findings have important implications for studies of vaginal infection of macaques with SIV or SHIV and further our understanding of how the bacterial microbiota influences HIV infection.
Introduction
B
Vaginal infection of macaques with SIV or SHIV is an important model for studying HIV sexual transmission. However, despite the established correlation between HIV susceptibility and changes in vaginal microbiota in humans, few studies have addressed the microbial factors that affect lower genital tract microbiota and their metabolic products in macaques. A recent analysis of rhesus macaques' lower genital tract bacterial microbiota by direct sequencing showed that it had similarities to that seen in women with BV in that they both were polymicrobial and had low levels of lactobacilli. 10 A number of studies show that macaque vaginal fluid has a relatively high pH. 10 –12 The goal of the present study was to characterize the lower genital tract environment of macaques with respect to lactic acid and glycogen levels. Here, we report that the genital fluids from these animals have significantly lower levels of lactic acid when compared to the genital fluid of women who do not have BV and were similar to that of women with BV. Moreover, macaques had significantly less glycogen in their genital fluid than women with or without BV. This was true in both cross-sectionally and longitudinally collected samples. Together, the data suggest that low glycogen levels may contribute to low frequencies of lactobacilli, resulting in the reduced vaginal acidity observed in macaques.
Materials and Methods
Macaque vaginal samples
Rhesus macaques (Macaca mulatta) of Indian origin were housed at the Tulane National Primate Research Center (TNPRC, n=11). Additional rhesus macaques of Chinese origin (n=12) and pigtail macaques (n=18) were housed at the Centers for Disease Control and Prevention (CDC). All animal studies were reviewed and approved by the Tulane University Animal Care and Use Committee or CDC's Institutional Animal Care and Use Committee. All animals were housed in accordance with the American Association for Accreditation of Laboratory Animal Care, and the Guide for Care and Use of Laboratory Animals, National Academy Press. For the Tulane samples, macaques were purchased and imported from the Caribbean Primate Research Center 7 months prior to first sample collection. Prior to importation, the animals were born and raised in group breeding colonies, so their parentage and prior sexual activity were unknown. Following arrival at the TNPRC, macaques were given complete physical examinations, blood work (CBC, chemistries), and routine stool examinations for parasites, and all animals were deemed to be normal, healthy, cycling mature females 4–14 years of age at sample collection, weighing 4–11 kg. The CDC's Chinese rhesus monkeys were obtained from Battelle Biomedical Research Center. The CDC's pigtail macaques were purchased from either the breeding colony of Yerkes National Primate Research Center (n=4) or from New Iberia Research Center (n=14). Following arrival at CDC, macaques were routinely given complete physical examinations, blood work (CBC, chemistries), and an evaluation of the female reproductive tract including colposcopic examination for signs of inflammation. All were deemed to be normal, healthy, cycling mature females 10–16 years of age at sample collection, weighing 6–11 kg. All macaques tested negative for simian immunodeficiency virus (SIV), simian retroviruses, and simian T cell leukemia viruses. Animals were not Depo-provera treated or synchronized for menstrual cycle.
Macaques were anesthetized using standard doses of ketamine hydrochloride and genital tract samples were collected by cervical-vaginal lavage (CVL), performed by irrigation of the vaginal vault with either 4 ml of sterile nonbacteriostatic saline (Tulane samples) or 8 ml of phosphate-buffered saline (PBS) (CDC samples). CVL was held on ice until processing, which occurred within 6 h of collection. At CDC, blood was also collected from pigtail macaques into Vacutainer tubes (CPT; B.D., San Jose, CA) as described by the manufacturer to assess progesterone levels. Sample collection occurred weekly in otherwise untreated animals for 12 consecutive weeks.
Human subjects
All studies were approved by the Institutional review boards of Rush University Medical Center and Cook County Stroger Hospital. The study population was recruited at the Ruth M. Rothstein CORE Center STD Screening Clinic of Cook County Stroger Hospital. Female patients presenting for an STD evaluation were approached for study participation and written informed consent was obtained from all subjects. Standard of care genital examinations were performed and swabs taken for diagnosis. CVL samples were then obtained by irrigation of the cervix with 10 ml of nonbacteriostatic sterile saline, followed by aspiration from the posterior fornix. Urine was also obtained for a pregnancy test. None of the subjects was pregnant. All subjects used in this study were negative for STDs using the following tests: BD Probetec ET (Becton Dickinson, Franklin Lakes, NJ) for Chlamydia trachomatis and Neisseria gonorrhoeae; culture for herpes; a rapid plasma reagin test (Arlington Scientific, Utah) for syphilis; an enzyme immunoassay (Bio-Rad Laboratories, Hercules, CA) for HIV; wet mount examination and ELISA for the p65 protein (HyTest, Turku, Finland) were both performed for Trichomonas; wet mount for clue cells, vaginal pH, whiff test, and Nugent Gram stain for bacterial vaginosis; and a KOH preparation was examined microscopically for Candida.
Lactate, protein, and glycogen measurements
Lactate levels were measured in CVL using the Lactate Assay Kit (Eaton Biosciences, San Diego, CA). Protein in CVL was measured using the BCA Protein Assay Kit (Thermo Fisher Scientific, Rockford, IL). Glycogen in CVL was measured using the Glycogen Assay Kit (Biovision, Mountain View, CA). Glycogen and lactate levels were standardized to total protein levels in the CVL. All measurements were within the standard curve of each respective assay. Plasma samples were shipped on dry ice to the University of Wisconsin National Primate Research Center where progesterone levels were measured using an enzyme-linked immunosorbent assay (ELISA). 13
Statistical analysis
Data were first analyzed for homogeneity of their distributions. For Figs. 1, 3, and 4, the Kruskal–Wallis test was performed and p-values were calculated using Dunn's Multiple Comparison post test because samples showed similar distributions. For Fig. 2, a Mood's Median test was used because of differences in the shapes of the data distribution in BV+ women, which, similar to that of both pigtail and rhesus macaques, were skewed. The glycogen levels in non-BV women were more normally distributed. Data were analyzed using SAS (Figs. 1, 3, and 4) and R (Fig. 2) softwares.
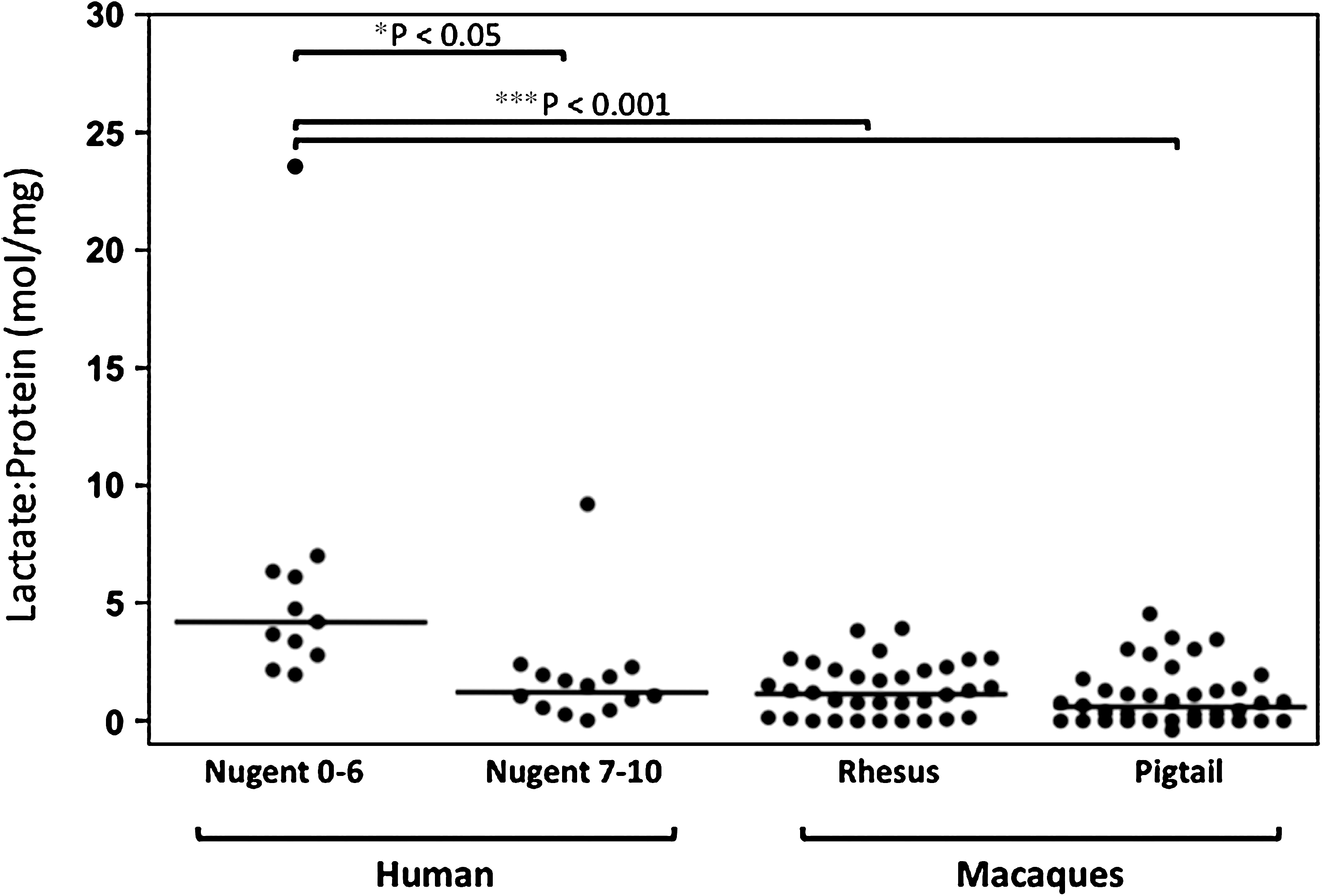
Lactate:protein levels in women and macaques. Cervical-vaginal lavage (CVL) samples from 25 human subjects and 23 rhesus and 18 pigtail macaques were collected as described in Materials and Methods. Some macaques were assessed more than once. Lactate and protein levels were measured per manufacturer's instructions. The two-tailed p-values were calculated using the Dunn's Multiple Comparisons post test. Bacterial vaginosis (BV) in women was determined by Nugent Gram stain values of 7–10, while non-BV was 0–6. No significant differences were found in humans with BV compared to rhesus or pigtail macaques.
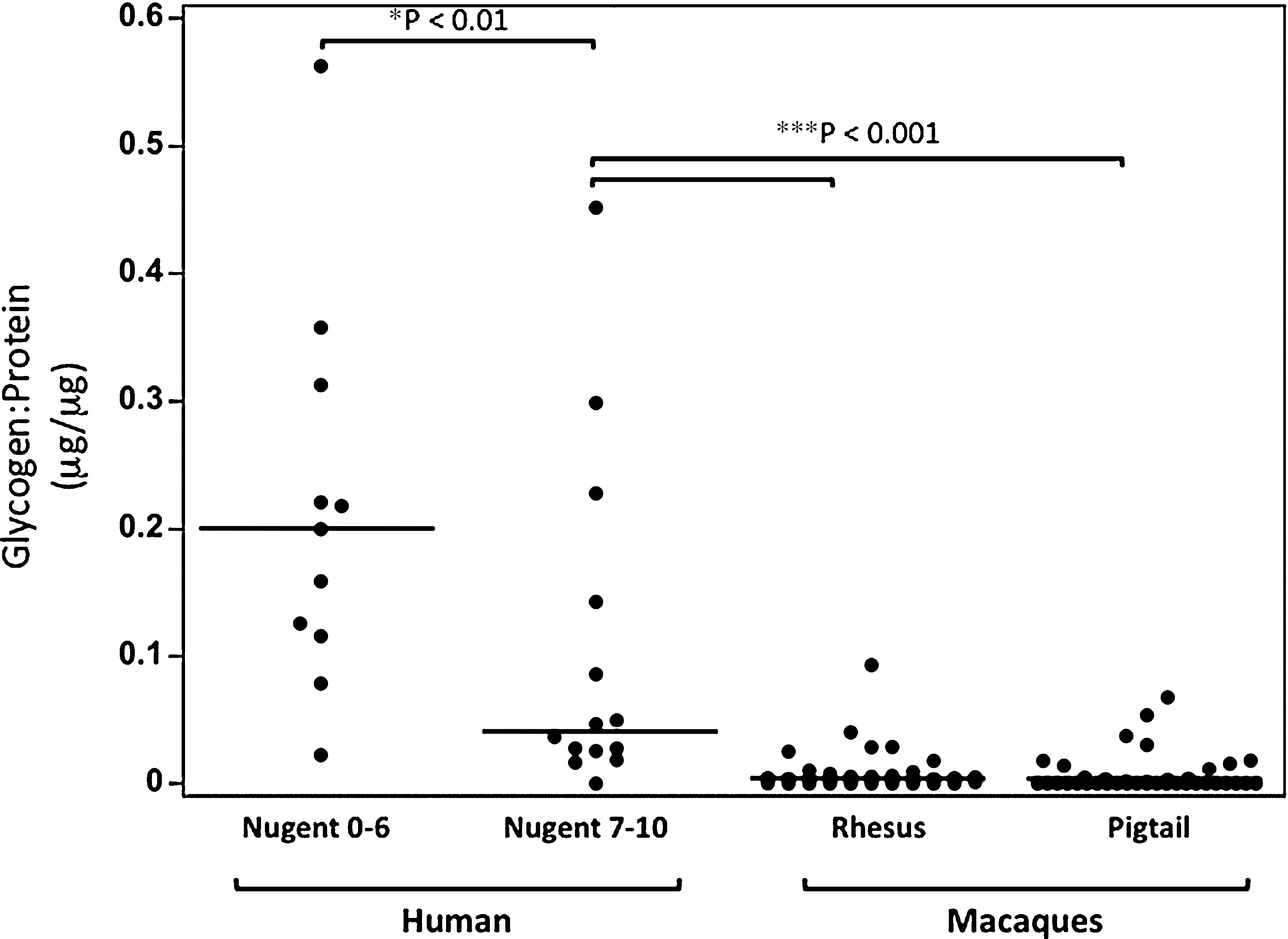
Glycogen:protein levels in woman and macaques. CVL samples were collected as described above (Fig. 1 and Materials and Methods). Glycogen and protein levels were measured per manufacturer's instructions. The two-tailed p-values were calculated using the Mood's Median test. As in Fig. 1, BV in women was determined by Nugent Gram stain values of 7–10, while non-BV was 0–6. Glycogen:protein levels in macaques were significantly lower than in women without BV (p<0.001, Mood's Median test), but were omitted from the figure for the sake of clarity.
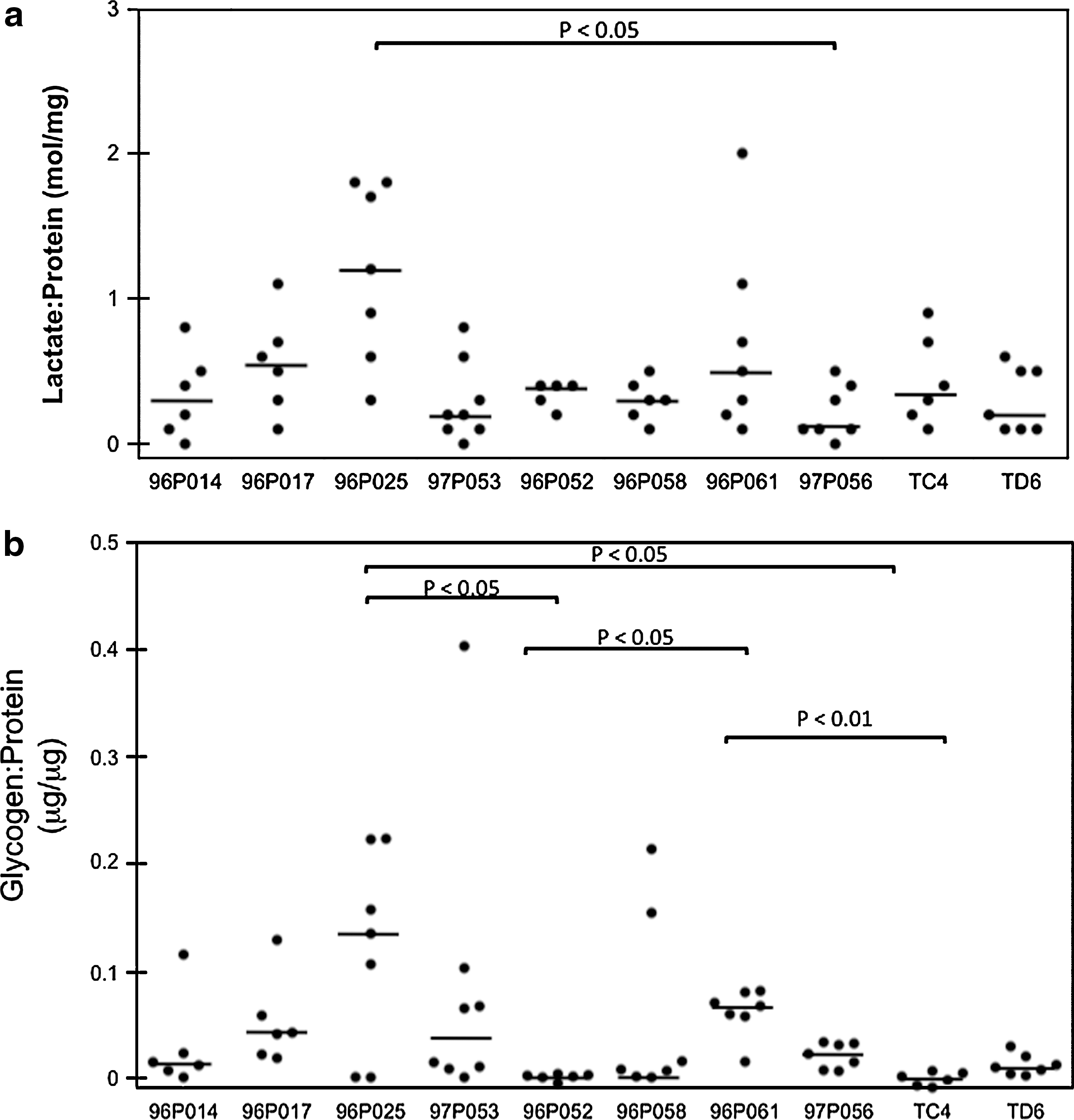
Lactate and glycogen levels collected over the menstrual cycle in pigtail macaques. CVL samples were collected as described in Materials and Methods. Lactate

Lactate:protein and glycogen:protein levels at different stages of menstrual cycle. CVL samples were collected and lactate
Results
Comparison of lactate levels in rhesus and pigtail macaques with humans
Since the lactic acid produced by lactobacilli is primarily responsible for the acidic pH of the vagina in women with microbiota dominated by lactobacilli, 7,8 we compared the lactic acid levels in mucosal fluid of humans and macaques. As shown in Fig. 1, lactate:protein ratios were significantly higher in women without BV [Nugent score 0–6, median=4.2 mol lactate/mg protein (mol/mg)] than those with BV (Nugent score 7–10, median=1.3 mol/mg, p<0.05, Dunn's Multiple Comparisons post test) while the lactate:protein levels in the fluids collected from rhesus and pigtail macaques were similar to those in women with BV (median=1.2 and 0.7 mol/mg, respectively). Because the animals' habitat could influence the genital microflora, we tested macaques from two different primate centers and found that they had similarly low lactate:protein ratios regardless of the species tested (rhesus vs. pigtail) or the facility in which they were housed (Tulane University vs. the CDC, data not shown). As previously reported, 10 only 2 out of the 11 rhesus monkeys, housed at the Tulane facility and whose microbiota were analyzed by pyrosequencing, were colonized with lactobacilli. Interestingly, these two monkeys had increased levels of lactate:protein; one had 11% lactobacilli and a lactate:protein of 2.7 mol/mg, while the other had 39% lactobacilli and a lactate:protein of 2.3 mol/mg, compared to their cohort's median of 1.2 mol/mg. These data show that the reported scarcity of lactobacilli observed in the macaques 10 is associated with relatively low levels of lactic acid when compared to women without BV.
Comparison of glycogen levels in rhesus and pigtail macaques with humans
Lactobacilli are thought to metabolize the glycogen made by vaginal epithelial cells to produce lactic acid. 6 –8 Since we found that lactate:protein levels were low in macaques and women with BV relative to women without BV, we also assessed glycogen levels in mucosal fluids. As shown in Fig. 2, there were significantly higher levels of glycogen:protein in genital fluids of women without BV [Nugent score 0–6, median=0.2 μg glycogen/μg protein (μg/μg)] than in women with BV (Nugent score 7–10, median=0.04 μg/μg, p<0.01, Mood's Median test). Strikingly, the glycogen:protein ratios were on average 50-fold lower in macaques than they were in non-BV women and more than 10-fold lower than the BV+ samples (medians=0.004 μg/μg and 0 μg/μg for rhesus and pigtail monkeys, respectively). The glycogen:protein levels in macaques were significantly lower than in women with or without BV (p<0.001, Mood's Median test). No institution-based differences in glycogen levels were observed in the macaques (data not shown).
Lactate and glycogen levels collected over the menstrual cycle in pigtail macaques
Since glycogen abundance in the lower genital tract of humans and macaques is thought to be affected by estrogen, 6 –8 we examined 67 vaginal fluid samples collected from 10 pigtail macaques at different times during the menstrual cycle to determine if the lactate and glycogen levels fluctuated. As shown in Fig. 3, the lactate:protein as well as glycogen:protein levels generally remained low in these animals over the approximately 2 months of observation (medians=0.4 mol/mg and=0.015 μg/μg, respectively). However, there was some animal-to-animal variability. For example, animal 96P025 had significantly higher lactate:protein ratios (median 1.2 mol/mg) than animal 96P056 (median=0.1 mol/mg, Fig. 3a, p-value<0.05, Dunn's Multiple Comparisons post test). Also, glycogen:protein ratios were significantly higher in monkey 96P025 compared to monkey 96P052 (medians=0.13 μg/μg and 0.001 μg/μg, respectively, p-value<0.05, Dunn's Multiple Comparisons post test, see Fig. 3b). In addition, we observed that lactate:protein and glycogen:protein ratios could widely vary within an individual animal (e.g., 96P025). Further analysis of our data revealed that the levels of lactate in the vaginal fluids of these animals significantly correlated with their levels of glycogen (Spearman r=0.32, two-tailed p-value=0.0086).
To learn the relationship between sex hormones and lactate or glycogen levels in these macaques, we also compared lactate:protein and glycogen:protein ratios at three time points: 1 week before peak, at peak, and 1 week postpeak plasma progesterone. We found that the differences among the medians of lactate:protein (Fig. 4a) and glycogen:protein levels (Fig. 4b) at these time points were small and failed to reach statistical significance.
Discussion
In the current study we report that similar to women with BV, both rhesus and pigtail macaques have lactate:protein levels that are significantly lower than what is measured in women without BV (Fig. 1). These findings agree with our previously reported results that macaques' genital tract microbiota was similar to women with BV and had relatively lower levels of lactobacilli 10 and help explain the reduced vaginal acidity in both BV+ women 5 –8 and macaques. 10 –12 Additionally, we found no differences in lactate levels between the rhesus and the pigtail macaques. Also, both types of macaques had significantly lower levels of lactate than women regardless of whether the women's samples were or were not stratified by Nugent score (analysis not shown)
Vaginal acidity plays an important role in maintaining the health of the lower genital tract in women. 5 –8 It is believed that the lactic acid produced by lactobacilli is by and large responsible for the acidic pH of a healthy vagina. 7,8 Indeed BV, a condition in which the vaginal pH is perturbed (pH>4.5), is strongly associated with a myriad of sexually transmitted diseases including HIV infection. 1 –4 Similar to women with BV, macaques' lower genital fluid has also been reported to be polymicrobial 10 and less acidic. 10 –12 Given these results, macaques may be less protected against vaginal infections with SIV or SHIV.
Although there is currently some debate about the relative frequency and predominance of lactobacilli in macaques, some studies have shown that macaques' lower genital tract is polymicrobial, sharing many of the same bacteria types with BV+ women. 10 –12,14 –16 Interestingly, we found that the prevalence of other bacteria capable of producing lactic acid such as Streptococcus and Staphylococcus in some of the rhesus monkeys 10 resulted in lactic acid concentrations at levels seen in women without BV (Fig. 1). Despite this, however, several studies have reported that the vaginal pH in macaques is higher than what has been measured in women with or without BV. 10 –12 This suggests that in the dynamic ecosystem of the vagina, the acidity afforded by lactic acid may be neutralized by other bacterial products such as amines in these animals, as known to be the case in women with BV. 17 This will effectively render the vaginal environment less acidic and perhaps more prone to infections. Clearly, a careful analysis of the types of bacteria that reside in the lower genital tract and their metabolites will be pivotal when using these animals to study the interrelationship between the microbial flora, SIV/SHIV infection, and the presumed protective effects of lactic acid. Moreover, in light of our data, a scoring system in macaques, similar to the Nugent system used in humans, may reveal important information about these animals' vaginal environment and how it compares to that of women. Because bacteria can utilize the glycogen available in the vaginal epithelial cells as an energy source, 6 –9 and because glycogen is present in vaginal epithelia in both humans 18 and macaques, 19,20 we also compared glycogen levels in the genital fluid of macaques to that of women. We found that both macaques and women with BV had significantly lower levels of glycogen:protein than women without BV (Fig. 2). We also observed some small, albeit significant, differences in glycogen levels in rhesus versus pigtail macaques (analysis not show).
Interestingly, macaques' glycogen:protein levels were even lower than what was measured in BV+ women (p<0.001). While the explanation for this is not known, we might postulate that the synthesis, deposition, and/or the shedding of glycogen in macaques' vagina might differ from that in humans. Alternatively, macaques' lower genital tract microbiome might be such that the available glycogen is consumed differently than in humans. Regardless of what causes the low concentrations of glycogen in macaques' lower genital tract, our data suggest that lower glycogen levels in these animals can affect lactobacilli's frequencies and lactate levels in the macaques' CVL. This, in turn, may lead to reduced amounts of lactic acid present in the genital fluid of macaques and, to a lesser extent, women with BV, resulting in pH measurements that are higher than normal. Indeed, statistical analysis of our data collected longitudinally from cycling pigtail monkeys revealed that there was a significant, albeit small, correlation between lactate and glycogen concentrations in these animals (Spearman r=0.32, two-tailed p-value=0.0086). Interestingly, a strong correlation between lactate and glycogen levels was also observed in CVL samples collected from women (r=0.7, two-tailed p-value=0.0002), but not rhesus monkeys (r=0.23, two-tailed p-value=0.19).
Unexpectedly, we also found that neither lactate:protein nor glycogen:protein levels were significantly associated with changing progesterone levels in cycling pigtail monkeys (Fig. 4). While glycogen concentrations are generally thought to be the highest when estrogen levels peak during the menstrual cycle, 6 –8 our data are in agreement with some reports that glycogen levels are independent of hormonal changes associated with the menstrual cycle. 20,21
Our data help further characterize the environment of the macaques' lower genital tract and support a model in which lower glycogen levels in the vaginal epithelium may limit the growth of lactobacilli, which in turn leads to lower concentrations of lactic acid and a higher vaginal pH. As studies have shown that the inflammatory milieu created by BV increases HIV expression, 22,23 further analysis of the macaques' lower genital tract may be helpful in understanding how bacterial microbiota influences retrovirus infection and pathogenesis in humans and primate models.
Footnotes
Acknowledgments
We would like to thank Drs. Ellen Kersh, Nattawan Promadej-Lanier, Ron Otten, and Ms. Patricia Guenthner [Centers for Disease Control and Prevention (CDC)] for providing macaque samples and Drs. Toni E. Ziegler and Daniel Wittwer (University of Wisconsin National Primate Research Center) for progesterone measurements. This research was funded by NIH Grants U19 AI076981 and P30 AI082151.
Author Disclosure Statement
No competing financial interests exist.