
Abstract
Many HIV-infected infants progress to AIDS during the first year of life when antiretroviral therapy (ART) is not given. The immune determinants of progression to AIDS are not known. We hypothesized that distinct HIV-specific T cell responses correlate with viral load and survival over the first year of life. Whole blood of infants at 3, 6, 9, and 12 months of age was incubated with HIV antigens Gag and Env. The frequency of specific T cells producing interferon (IFN)-γ was then measured by flow cytometry. Viral load and CD4% in HIV+ infants were determined at each time point. ART was not available for this population at the time of sample collection. Those infants who survived to 12 months of age (n=12) had lower viral loads and higher Gag-specific CD8+ T cell responses at 3 months, compared with infants who died (n=8). Furthermore, the frequency of Gag-specific CD4+ T cells correlated inversely with viral load at 3 and 6 months of age. Together these data indicate that the early presence of quantitatively higher Gag-specific T cell responses in HIV-infected infants is associated with lower viral loads and decreased mortality in the first year of life. Our data support the design of a vaccine that preferentially elicits Gag responses, which may result in lower levels of viremia and possibly improve outcome.
W
Participants were enrolled in a study of the effect of HIV infection and exposure on the BCG-induced immune response. 12 Briefly, infants born to mothers in public health care facilities in the Western Cape, South Africa, were enrolled within the first 3 months of life, and followed up at 3, 6, 9, and 12 months of age. Mothers of the infants provided written informed consent before participation in the study, which was approved by participating institutional review boards. The groups of infants enrolled were composed of HIV-infected infants (HIV+), HIV-exposed, uninfected infants (HIV-EU), and HIV-unexposed, uninfected infants (HIV−). This study took place prior to the routine use of ART in South Africa.
At each study visit, blood was collected for viral load (Roche Amplicor) and CD4 T cell count in HIV+ infants. Immediately after venipuncture, 250 μl of heparinized whole blood was incubated with recombinant vaccinia viruses expressing HIV subtype C Gag or Env proteins (consensus sequences of rVV-gag1 and rVV-env, NIH AIDS Research and Reference Reagent Programme, 1.96 pfu/ml––a dose found optimal in preliminary experiments), together with the costimulatory antibodies anti-CD28 and anti-CD49d (0.5 μg/ml each, BD Biosciences). An empty vaccinia vector (rVV-C1) served as a negative control. Blood from all infants was also incubated with staphylococcal enterotoxin B (10 μg/ml) as a positive control; this induced an interferon (IFN)-γ response in all infants. 12 Blood was incubated at 37°C and 5% CO2 for 9 h, a duration found optimal in preliminary experiments. Brefeldin A (10 μg/ml, Sigma Aldrich) was added for the last 5 h of incubation. Red blood cells were then lysed and white cells fixed (FACS Lysing Solution, BD Biosciences). Fixed white blood cells were cryopreserved and stored in liquid nitrogen until use.
Fixed, stimulated cells were thawed and stained with the following antibodies for 1 h at 4°C in Perm/Wash Buffer (BD Biosciences): CD3-QDot605 (UCHT1, Invitrogen), CD8-PerCP-Cy5.5 (SK1, BD Biosciences), CD4-Pacific Blue (RPA-T4, BD Biosciences), and IFN-γ-Alexa Fluor 700 (B27, BD Biosciences). Acquisition was performed using a LSRII flow cytometer (BD Biosciences).
Flow cytometry data were analyzed using FlowJo (v8.86, Treestar). The frequency of cytokine-positive cells in the negative control sample (empty vaccinia construct) was subtracted from the frequency in the Gag- and Env-stimulated samples. A response was considered positive if it was greater than the median plus three times the median absolute deviation in the negative control samples (0.004% IFN-γ+ for CD4+ T cells and 0.04% IFN-γ+ for CD8+ T cells), and there were 20 more cytokine-positive cells in the positive sample than in the negative control sample. A minimum of 180,000 lymphocytes was acquired for each HIV+ infant (median 1,920,000, range 182,000–8,150,000). Data analysis was performed using Prism software (GraphPad, version 4.0). The nonparametric Mann–Whitney U test and Kruskal–Wallace test were used to assess differences between groups. Correlations were assessed using the nonparametric Spearman rank test. Data were considered significant if p<0.05.
Twenty HIV+ infants were enrolled in the study (Table 1). Eight of these infants died during the first year of life: four before 6 months of age and four within 9 months of age. Whole blood ICS results from one infant were excluded due to a missing negative control condition. All infants were ART-naive, except for three infants who started treatment during the course of the study: one at 8 months and two after 9 months of age.
HIV-infected infants.
HIV-exposed, uninfected.
HIV-unexposed, uninfected.
Frequencies of Gag- and Env-specific IFN-γ-producing T cells were determined with a whole blood flow cytometry-based ICS assay (Fig. 1A). We confirmed the specificity of responses by comparing the IFN-γ responses among the HIV+, HIV-EU, and HIV− groups of infants (Supplementary Fig. S1; Supplementary Data are available online at
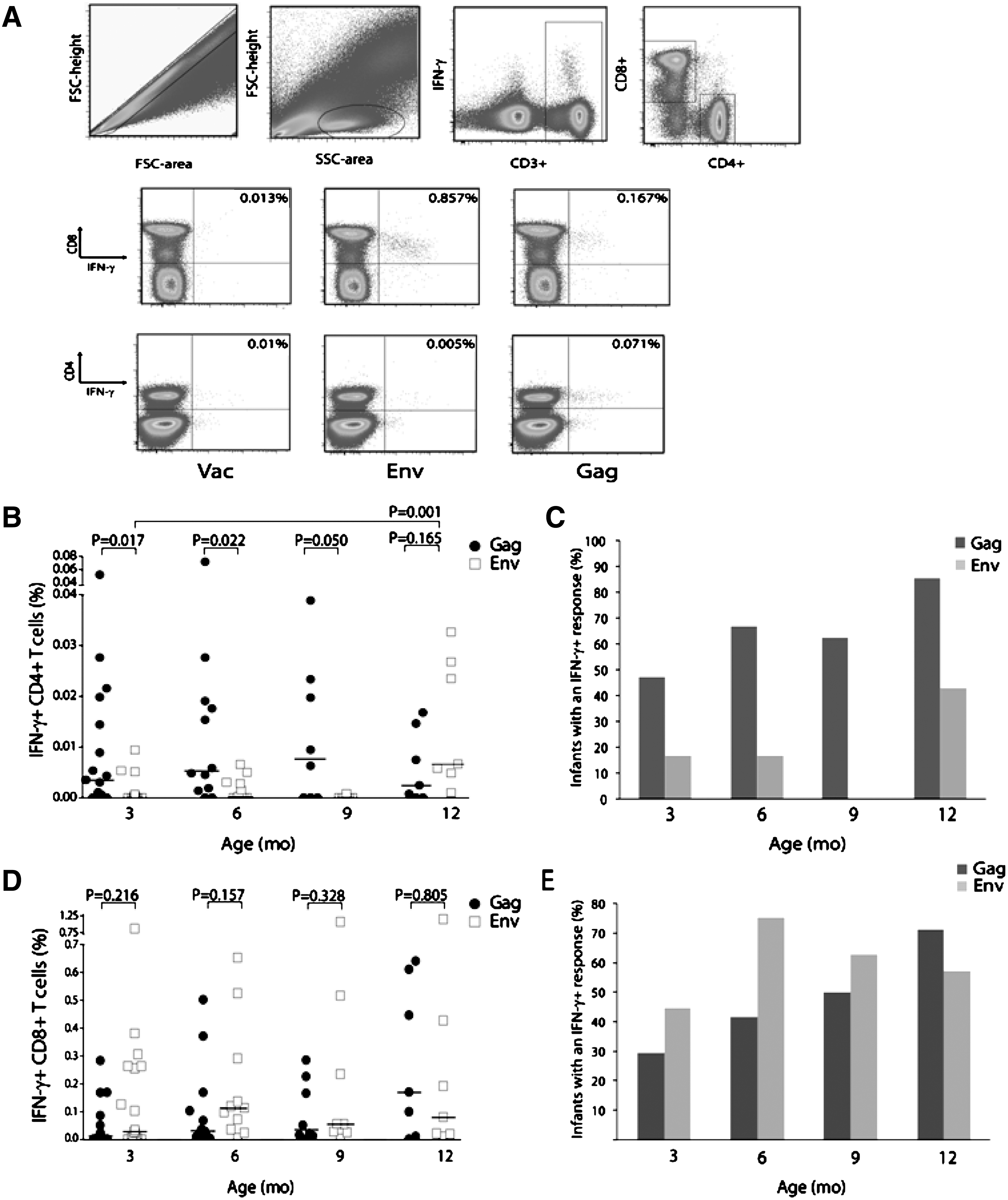
Gag- and Env-specific T cell responses at 3, 6, 9, and 12 months of age. Whole blood was incubated with Gag- or Env-expressing vaccinia constructs; intracellular interferon (IFN)-γ was measured by flow cytometry.
Gag-specific CD4+ T cell responses in the HIV+ group were relatively low in magnitude (Fig. 1B), although the proportion of infants with a detectable response increased over the first year of life (Fig. 1C). HIV-specific CD4+ T cell responses preferentially targeted Gag, compared with Env, although responses to the latter antigen were detectable in a significant proportion at 12 months of age (Fig. 1B and C).
In contrast to CD4+ T cell responses, the frequency of Env-specific CD8+ T cells appeared higher than that of Gag-specific CD8+ T cells at 3, 6, and 9 months of age, although this did not reach statistical significance (Fig. 1D). The proportion of infants who responded to Gag increased over time (Fig. 1E), whereas the proportion of infants responding to Env remained relatively similar over the first year of life.
To determine if early HIV-specific T cell responses were associated with clinical outcome, we compared these responses at 3 months of age in infants who survived and those who did not survive the first year of life. Higher frequencies of Gag-specific CD8+ T cells were present in survivors compared with nonsurvivors; there was no difference in Gag-specific CD4+ T cells between these groups (Fig. 2A and B). Furthermore, infants with detectable Gag-specific CD4+ and CD8+ T cell responses had lower viral loads at 3 months of age, compared with infants with no detectable responses (Fig. 2C and D). No differences were observed for Env-specific CD4+ or CD8+ T cell responses between survivors and nonsurvivors (data not shown).

Association between HIV-specific T cell responses and survival and surrogate markers of disease progression.
Comparing clinical outcome between the groups, we found that at 3 months of age, survivors had lower viral loads and tended to have a higher CD4% than nonsurvivors (Fig. 2E and F). Moreover, there was an inverse correlation between the frequency of Gag-specific CD4+IFN-γ+ T cells and viral load at 3 and 6 months of age (Fig. 2G and H). Although we observed lower viral loads in the infants with detectable Gag responses, we did not find a significant correlation between viral load and CD8+IFN-γ+ T cell frequencies. Altogether, these data suggest that the presence of early Gag-specific T cell responses by 3 months of age is associated with lower viral load and improved clinical outcome.
Previous studies have indicated that HIV-specific CD4+ T cell responses are of low magnitude in early infancy, especially in the first months of life. 4,11 Our data agree with these studies; however, we go on to show that there is an inverse correlation between the magnitude of early Gag-specific CD4+ T cell responses and viremia. Although an association between Gag-specific CD4+ T cell frequency and viral load has not been described in infants to date, Gag-induced proliferative capacity of CD4+ T cells has been shown to correlate inversely with viremia in adults. 13 –15 Our data suggest that early targeting of Gag by CD4+ T cells is associated with better virological control, although it is not possible from these results to determine the precise cause and effect relationship between HIV-specific T cell responses and control of viremia. An important limitation of our study is that we did not measure immune responses to all the HIV proteins, therefore we cannot exclude the possibility that the correlation we observed could be influenced by an immune response to another HIV antigen. Further studies comparing the full range of HIV proteins are needed in order to assess the precise contribution of Gag responses to control of viremia. In addition, functional studies that demonstrate the mechanisms by which these T cells contain viremia would further support our data. Due to limited blood samples, we were unable to do additional functional assays in this study.
Gag-specific CD8+ T cell responses have rarely been detectable in infants younger than 12 months of age, but have been readily detectable in older children. 11,16 These observations suggest that Gag-specific CD8+ T cell responses may be associated with better viral control and survival. Our findings support this hypothesis, as we showed that infants who survived the first year of life had a significantly higher frequency of Gag-specific CD8+ T cells at 3 months of age, compared with nonsurvivors; moreover, infants with a detectable Gag-specific CD8+ T cell response had lower viral loads than nonresponders. Preferential targeting of Gag by CD8+ T cells has been linked to better viral control in adults and in older children with HIV infection. 4,9,11,16,17 We did not find a correlation between the magnitude of Gag-specific CD8+ T cell responses and viral load, although the magnitude of a response is not the only indicator of viral control. Studies of HIV-infected adults have indicated that the ability of antigen-specific T cells to perform multiple functions simultaneously (“polyfunctional”) is associated with control of viremia. 18 Huang at al. showed that CD8+ T cell polyfunctionality is impaired in HIV-infected infants. 11 This could be why we do not see a correlation between the frequency of CD8+IFN-γ+ T cells with viral load in our study. Further studies are required to assess the relationship between the functional capacity of CD8+ T cell responses and disease progression in HIV-infected infants.
Our results provide additional evidence that Gag-specific responses are beneficial in infant HIV infection. Data from models of simian immunodeficiency virus (SIV) infection indicate that Gag epitopes are uniquely presented earlier than any other viral protein, suggesting that early recognition of Gag epitopes by SIV-specific CD8+ T cells may lead to rapid clearance of infected CD4+ T cells. 19 Recent data further indicate that Gag-specific CD8+ T cell responses may be more effective at neutralizing virus than Env-specific CD8+ T cells. 20 Additionally, immune escape mutations in CD8+ T cell epitopes in Gag can decrease viral fitness. 21,22
We found no association of Env-specific T cell responses with outcome, even though these responses are more dominant in CD8+ T cells than Gag-specific responses in the first year of life. In HIV-infected adults, the relative dominance of Env-specific responses over Gag-specific responses has been associated with poor virologic control. 9 Our results therefore appear to support the hypothesis that Env-specific responses contribute less to control of viremia, compared with Gag-specific responses.
Lohman et al. examined the association between HIV-specific T cell responses and viral load or mortality in infants. 3 In their study, IFN-γ responses were measured at 1 month of age, following incubation of peripheral blood mononuclear cells (PBMCs) with peptides from various HIV proteins, using an ELISPOT assay. No association between the strength and breadth of IFN-γ responses at 1 month and peak plasma viral loads, rates of decline of viral load, and mortality risk was shown. The differences observed in our study may be due to differences in the time of evaluation (1 month versus 3 months) and the methods used to detect HIV-specific T cells. Additionally, we analyzed Gag and Env responses separately and found significant associations only with Gag. Lohman et al. analyzed their data using combined responses to HIV epitopes, which may have masked the independent effects of Gag on viral control. Furthermore, in contrast to Lohman et al., our small sample size precluded us from applying analytical models to evaluate the possible independent effects of HIV-specific immune responses and early low viral loads on clinical outcome.
Similarly, Huang et al. measured HIV-specific T cell responses using an IFN-γ ELISPOT assay in their cohort of children, 5 weeks to 10 years old, and found an inverse correlation between the breadth and magnitude of HIV-specific responses and viral load after adjusting for age. 11 Our study is unique in that we used flow cytometry, which allowed us to differentiate CD4+ and CD8+ T cell populations and determine their associations with viral load. A caveat of this study was the use of vaccinia-based antigens used to stimulate cells in our assay system; overlapping peptide pools were not available to us at the time the samples were being collected.
In summary, our results indicate that the absence of early Gag-specific T cell responses in infancy may be predictive of disease progression and HIV-related infant mortality. Our data support the hypothesis that HIV vaccine strategies should aim to elicit strong and broad Gag-specific responses in recipients. Such a vaccine could curb the high viral loads typical of infant HIV infection in the first few years of life, and may result in an improved prognosis.
Footnotes
Acknowledgments
We would like to thank all the mothers and infants who participated in this study, as well as the clinical team at our field site. We would also like to acknowledge our funders, the Tuberculosis Research Unit of the National Institutes of Health (NO1-AI70022) and the Elizabeth Glaser Pediatric AIDS Foundation. B. Nqoko was also supported, in part, by the National Research Foundation of South Africa, and the Duncan Baxter Scholarship and an Equity Scholarship from the University of Cape Town.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.