
Abstract
Human immunodeficiency virus 1 (HIV-1) infection is characterized by genetic diversity such that specific viral subtypes are predominant in specific geographic areas. To determine circulating subtypes of HIV-1 in different parts of central Kenya, a cross-sectional study was carried out on HIV-1-positive blood samples collected from consenting individuals in eight hospitals of Kenya's central province. Proviral DNA was extracted from peripheral blood mononuclear cells. Polymerase chain reaction and direct sequencing using primers generated from a highly conserved region of HIV-1 env gp41 were carried out. Ninety-six samples were successfully amplified and sequenced. Analysis of the sequences showed that a majority of them belonged to subtype A1 (67/96, 69.8%), followed by subtypes D (18, 18.7%) and C (11/96, 11.5%). Consistent with findings in other parts of Kenya, HIV-1 subtype A1 was the most dominant virus in circulation. Continued surveillance of circulating subtypes of HIV-1 in Kenya is important in determining the evolution of the HIV/AIDS epidemic in Kenya.
The HIV/AIDS epidemic is a major global public health crisis with an estimated 33 million people worldwide living with HIV infection. 1 The majority of HIV cases (67%) are in sub-Saharan Africa. 1,2 HIV-1 has evolved into different subtypes and subsubtypes that differ in geographic distribution across the world. 3 The large genomic diversity of HIV-1 in different geographic regions is a consequence of the high mismatch error rate of the HIV reverse transcriptase (RT) enzyme coupled with the absence of an exonuclease proofreading activity. 4 As a result of this, three groups of HIV-1 have evolved across the globe: M (major), O (outlying), and N (new). 3 Among the M group, which accounts for more than 90% of reported HIV/AIDS cases, viral envelopes have diversified so greatly that this group has been further subclassified into nine major subtypes including A–D, F–H, J, and K, as well as 49 circulating recombinant forms.
Genetic differences of the different groups and subtypes of HIV-1 are based on differences in the envelope protein nucleotide sequence. The env proteins of groups M and O differ by as much as 30–50%. The N subtype, in turn, appears to be phylogenetically equidistant from groups M and O. 5 Within the M subgroup, intersubtype env variations differ by 20–30%, whereas intrasubtype variation of 10–15% is observed. 6 Variation in the env gene may play an important role in viral entry and pathogenicity, both directly and indirectly. Variations in the pol gene, although less divergent, are particularly important in determining susceptibility of the virus to antiretroviral therapy. 4
Several studies have suggested that AIDS progression differs as a function of the infecting subtype and viral tropism. 7 –9 The differences in subtypes may also be a subject of concern when choosing assays to use for viral load testing. 10
Kenya is among the countries of sub-Saharan Africa in which the HIV pandemic has had an overwhelming effect. Estimates suggest that by the end of 2008, about 1.5 million Kenyans were infected with HIV. 11 HIV-1 subtype surveillance has shown that the majority of the HIV cases in Kenya belong to HIV-1 subtype A1. Despite this, there is an increasing prevalence of other subtypes and recombinants due to the nature of the virus and continued interactions of infected people from different regions of the world.
Most subtype epidemiological studies carried out in Kenya have been done in urban areas. 12 –14 Little is known about circulating subtypes of HIV-1 in rural areas, which are the home to many of the infected people. In this study, we sought to determine the circulating subtypes of HIV-1 in eight districts of central Kenya in which HIV-1 prevalence has been known to be high. Ethical clearance to carry out this study was obtained from the Kenya Medical Research Institute (KEMRI)/National Ethics Committee. The study was approved under protocol no. SSC1374.
In this cross-sectional study, 5 ml of whole blood was collected in EDTA vacutainer tubes from consenting participants who were 18 years or older. The samples were collected from different health centers in central Kenya and anonymized by giving them unique identifiers that were not linked back to the donors. The health centers were chosen based on their location, the number of patients that visit the hospital or are referred from elsewhere within Kenya's Central Province, and the logistics of sending the samples to the central laboratory in Nairobi for analysis. Collected samples were tested for anti-HIV-1 antibodies using Unigold and Determine rapid tests following the manufacturer's instructions, and the positive samples selected. Peripheral blood mononuclear cells (PBMCs) were extracted from whole blood by density gradient centrifugation using Ficoll-Paque Plus (Pharmacia) and Proviral DNA extracted using DNAzol (Gibco BRL) and ethanol precipitation as previously described. 12,13
A segment of the env gp41 (corresponding to nucleotide 7850–8310 of HIV-1HXB2) was amplified with primers, gp40F1 5'-TCTTAGGAGCAGCAGGAAGCACTATGGG-3′ and gp41R1 5'-AACGACAAAGGTGAGTATCCCTGCCTAA-3' in the first round and gp46F2 5'-ACAATTATTGTCTGGTATAGTGCAACAGCA-3' and gp47R2 5'-TTAAACCTATCAAGCCTCCTACTATCATTA-3' for nesting. Amplification was done with one cycle of 95°C for 10 min and 35 cycles of 95°C for 30 s, 55°C for 30 s, and 72°C for 1 min, with a final extension of 72°C for 10 min. The amplicons were further labeled using Big Dye with a set of nested primers and sequenced using the ABI 310 genetic analyzer. Generated sequences were aligned using Gene cutter and phylogenetic analysis was done using phyML.
15
Trees were visualized using the FigTree software (
Eight health centers in the central province participated in this study (Table 1). From each center, 12 samples were successfully analyzed to determine prevalent HIV-1 subtypes. Overall, 96 samples were sequenced. Analysis of these sequences showed that the majority belonged to subtype A1 (67/96, 69.8%) followed by subtypes D (18/96, 17.7%) and C (11/96, 11.5%). Figure 1 shows a phylogenetic tree of all the 96 samples that were analyzed together with the HIV-1 reference sequences from the Los Alamos HIV database.
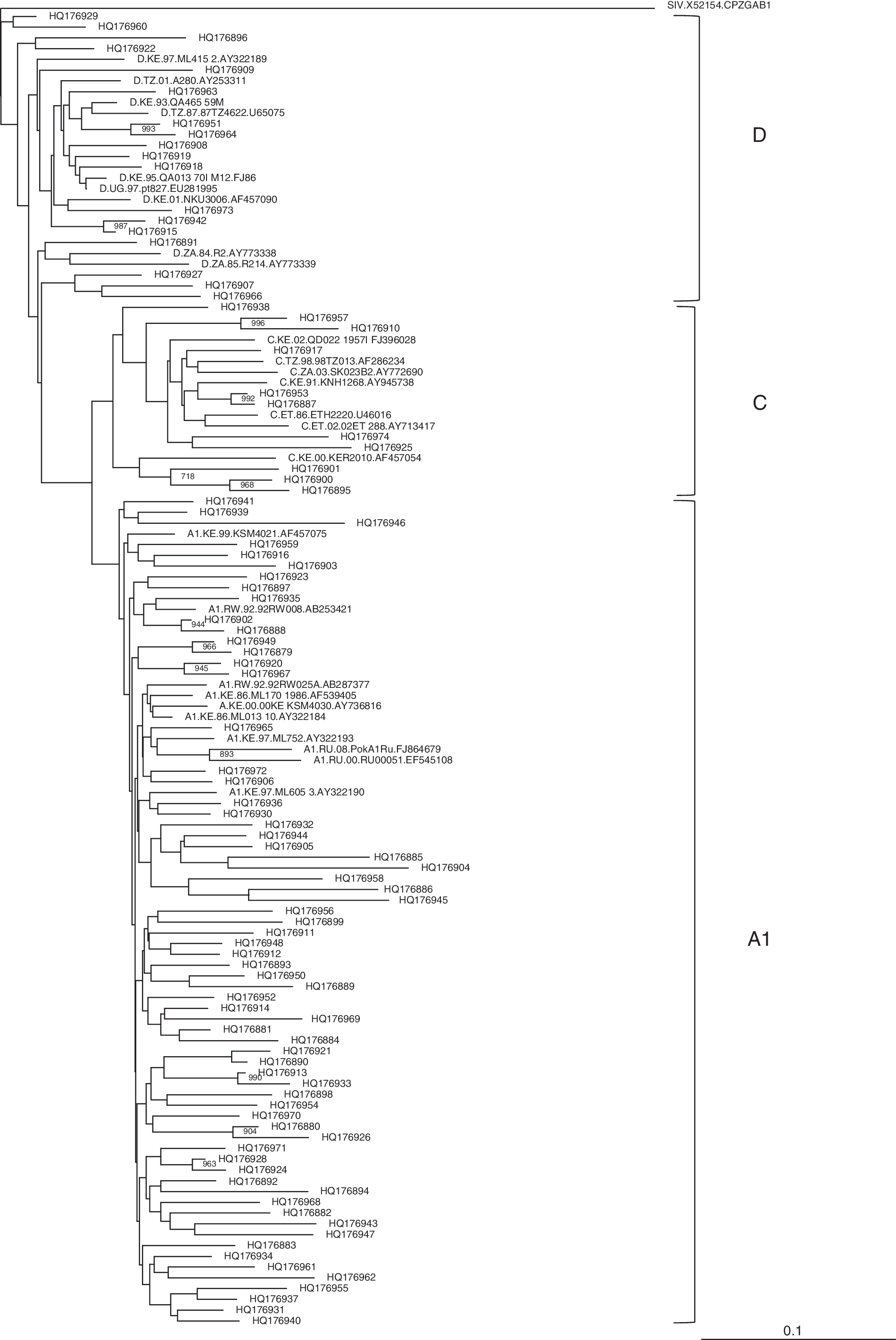
Phylogenetic tree of the HIV-1 subtypes in eight major health facilities in central Kenya. Patient sample sequences (indicated by accession numbers) were aligned and compared with reference sequences from the Los Alamos HIV database. Phylogenetic relationships were constructed using the neighbor-joining method and rooted with SIVcpzGAB. The bootstrap values of 1000 replicates are indicated next to the node. Bootstrap values of >70% (of 1000 replicates) are indicated.
HIV-1 Subtype Prevalence Among Infected Patients Who Seek Treatment in Major Health Facilities of Central Kenya
Previous studies have shown that HIV-1 subtype A1 is the most dominant virus in circulation in Kenya. 11 This study showed that HIV-1 subtype A1 was the dominant subtype in circulation in central Kenya, involving more than 69% of all the samples analyzed. This information shows that the HIV-1 epidemiology in central Kenya is similar to that of many other parts of the country where HIV-1 subtype A1 is the dominant virus in circulation. It is sufficient to say that the HIV/AIDS epidemic in Kenya is driven mainly by HIV-1 subtype A1. However, as seen from clustering in the phylogenetic tree in Fig. 1, there was a high diversity even within subtypes as demonstrated by low bootstrap values.
As is the case in other cross-sectional studies, this study had limitations. One, the samples were anonymously collected with no demographic information such as gender and age of participants. This would not enable us to do an analysis based on gender disparities among the studied population, though it is known that women are disproportionately affected by the HIV pandemic worldwide. 1 Second, only positive samples were referred for further analysis on subtypes in circulation; therefore we could not determine the site prevalence of HIV in the studied population. However, the prevalence may be extrapolated from the Kenya AIDS indicator survey report, which indicated that the prevalence of HIV in this region was 3.8%. 11 Third, these patients were referred to the indicated centers for ultimate treatment and care, yet no data were available as to their immune status, i.e., CD4 cell counts and viral loads. As such, we could not ascertain drug susceptibility levels and resistance to available regimens. Four, the sample size was too small to give a generally acceptable picture of the prevailing status of subtypes circulating in central Kenya. Nonetheless, this provides a picture of HIV-1 subtypes circulating in central Kenya.
Sequence Data
All the 96 sequences generated from this study were deposited in GenBank under accession numbers HQ176879–HQ176974.
Footnotes
Acknowledgments
We acknowledge The Kenya Medical Research Institute for enabling us to carry out this research at its facility in Nairobi. This study was funded in its entirety by funds generated at the Centre for Virus Research, Kenya Medical Research Institute.
Author Disclosure Statement
No competing financial interests exist.