
Abstract
Human immunodeficiency virus (HIV)-1 infection compromises the natural killer (NK) cell function and leads to defective control on virus multiplication. One of the major features of HIV-1 infection is the expansion of a functionally compromised defective NK cell subset (CD56−CD16+). We analyzed the NK cell subsets in early HIV infection to determine the effect of NK cell perturbation on the viral load set point, a marker of disease progression. We report that the defective NK cells are expanded in early HIV infection within 6–8 months of acquiring infection and are correlated with a higher plasma viral load set point, suggesting its utility as a predictive marker for disease progression. The expression of CD161, a molecular marker responsible for proliferation and differentiation of NK cells, was significantly down-regulated in the defective NK cells as compared to slow progressors (p=0.0009) and healthy controls (p=0.0003) and was correlated with a higher viral load set point in early HIV-1 infection (r=−0.6154, p=0.03), suggesting the probable role of CD161 expression in the impaired proliferation and differentiation of defective NK cells into the functional NK cells in early HIV infection. The reduction in CD161 expression on the defective NK cells in early HIV infection is thus indicative of the role of innate immune cells in early control of HIV infection.
T
NK cell activity has been shown to be governed by two different subsets of NK cells, cytotoxic (CD56+16+), responsible for cytolytic activity and comprising 90% of the circulating NK cells, 3,4 and regulatory (CD56++16−/dim), (10–15% of circulating NK cells) responsible for the abundant production of cytokines and chemokines mediating suppression of viral infection, prevention of viral entry, and also recruitment and priming of cells of innate and adaptive immunity. 1,5
HIV infection is known to compromise the innate as well as adaptive immune function of the host leading to functional impairment of several hematopoietic cell lineages including NK cells and dendritic cells. 6 Numerous studies have shown that the total NK cell number is decreased in acute and chronic HIV infection. 7 –9 The NK cells are found to be deficient in controlling the spread of HIV-1 and are defective in the clearance of HIV-1-infected autologous CD4+ T cells. 10,11 The reduction in NK cells was found to correlate with low CD4+ T cell count 12 and increasing viral load. 13 –15
Recently, it has been observed that the chronic phase of HIV infection showed the presence of dysfunctional/defective NK cells (CD56−CD16+) that correlated with high viral load. 13,16 –18 Sequential impairment of NK cell function has been seen in patients with acute HIV infection having persistent viral replication resulting in an increase of CD56−CD16+ defective NK cells. 19
The various actions of NK cells are a result of a fine balance between signal transduction executed by various activatory and inhibitory receptors after ligation of their cognate ligands. It has been shown that the expression of natural cytotoxicity receptors (NCRs), NKp46, NKp30, and NKp44, is markedly decreased among viremic individuals along with a concomitant decrease in NK cytolytic activity. 16,20 Additionally, NK cells also express CD161, C-type lectin-like receptors, and the expression of this receptor was found to correlate with the cytotoxic function of NK cells. 21,22 CD161 is a marker involved in proliferation, differentiation, and apoptosis, depending on the pulse of ceramide production, 23 and is present on NK cells, subsets of thymocytes, and mature CD3+ T cells including CD4+ and CD8+ cells. The role of CD161 expression on NK cells has not been explored in HIV infection.
We hypothesized that CD161 expression might play a significant role in the impairment of NK cell function and the presence of defective NK cells in HIV infection. We have chosen the early phase of HIV infection to study the influence of defective NK cells as well as CD161 expression on NK cells on the rate of disease progression using the plasma viral load (PVL) set point as a surrogate marker of disease progression.
From the patients visiting the out-patient clinics of the National AIDS Research Institute, Pune, India, 18 HIV-infected patients with a recent history of exposure and with less than 7 months (range 3–7 months) duration between the last HIV-negative and first HIV-positive test were enrolled and followed up for 2 years to obtain the plasma viral load set point. The blood samples collected at the enrollment visit were processed and the data were used for the analysis. Nine individuals with slow disease progression who were HIV-1 positive, asymptomatic, and antiretroviral treatment (ART) naive with a stable absolute CD4 count above 500 cells/μl for the past 3 years were also enrolled. Additionally, 18 HIV-seronegative healthy individuals were also included in the study. The study has been approved by the institutional Ethics Committee and the whole blood samples were obtained after obtaining written informed consent.
Peripheral blood mononuclear cells (PBMCs) were isolated from whole blood using Ficoll density gradient centrifugation and stored at −196°C until use. The cryopreserved PBMCs of the study participants were revived and incubated in RPMI 10% fetal calf serum (FCS) in a humidified 5% CO2 incubator overnight. The next day, the PBMCs were stained with anti-CD3 PerCP, anti-CD16 FITC, anti-CD56 APC, and anti-CD161PE (BD Biosciences, Singapore). After staining, cells were washed twice with phosphate-buffered saline (PBS) containing 0.5% bovine serum albumin. The cells were then fixed with 2% paraformaldehyde in PBS, pH 7.2, acquired on a FACSCalibur (Becton Dickinson, USA) and analyzed using Cell Quest Pro Software. The lymphocyte population was gated using forward and side scatter (Fig. 1A) while acquiring the sample and 50,000 gated events were acquired. The NK cells were identified as (CD3−CD16+CD56+) cells (Fig. 1B) and further categorized into different NK cell subsets depending on the expression of CD16 and CD56 molecules as regulatory (CD56++ 16−), cytotoxic (CD56+16+), and defective (CD56−16+) (Fig. 1 C).
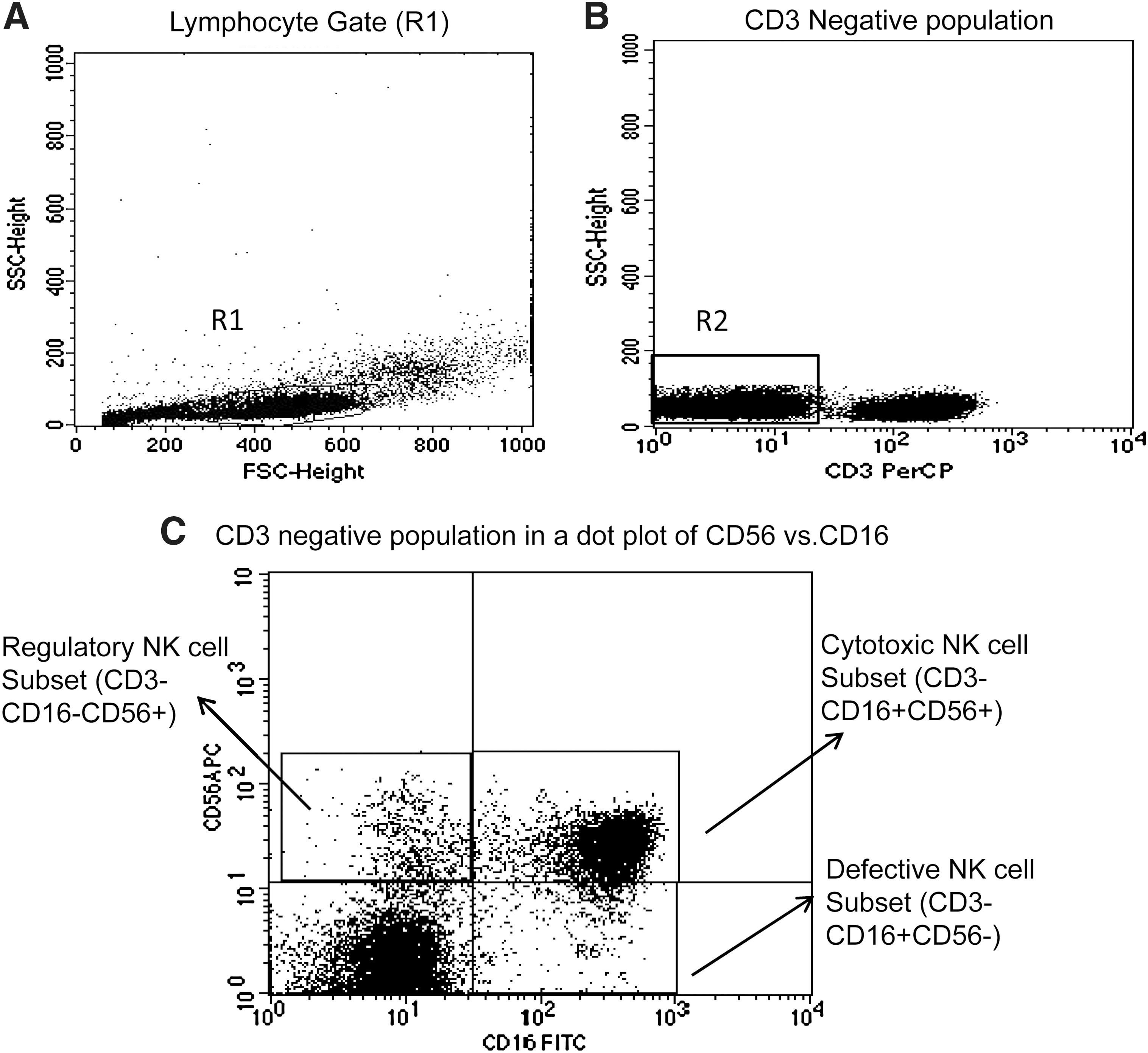
Gating strategy used to assess the NK cells and the NK cell subsets. The lymphocytes were live gated during acquisition using the side and forward scatter dot plot display
CD161 expression on the NK cells was expressed as mean florescence intensity (MFI). The PVL set point was calculated by determining the geometric mean titer (GMT) of viral load results obtained between 6 and 15 months after the estimated date of infection.
The data were analyzed using Graph Pad prism (version-5). The Mann–Whitney U test was used to analyze continuous variables in all three study groups. The association between different NK cell subsets, the CD161 expression estimated at the enrollment visit (i.e., within 7 months of acquiring HIV infection), and the PVL set point was determined using the Spearman correlation coefficient. A p value ≤0.05 was considered significant for all analyses.
Total NK cell percentage was significantly reduced in HIV infection both in recent phase (mean 8.75%±1.2, range 7.2–11.2%, p=0.0001) and patients with slower disease progression (mean 10.8%±3.9, range 4.2–16.3%, p=0.015) as compared to HIV-seronegative healthy controls (mean 14.34%±3.3) (Fig. 2A).
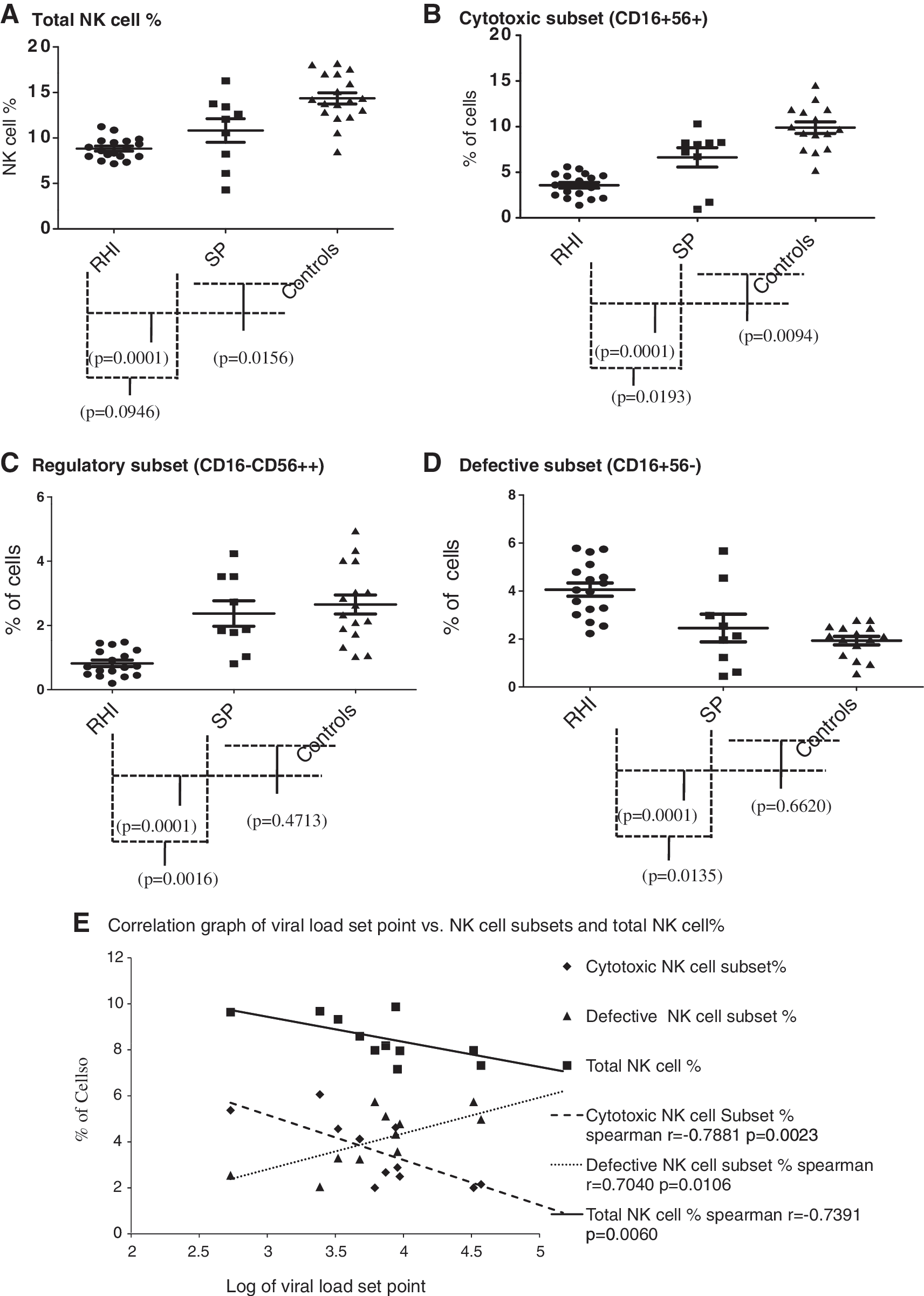
The percentages of total cells and subsets in the study groups. The total NK cell percentages in patients with recent HIV infection (RHI), slow progressors (SP), and healthy HIV-seronegative individuals (Controls) are shown in
The percentages of cytotoxic (CD16+56+) as well as regulatory (CD56++16−) NK cell subsets were also reduced significantly in recent HIV infection (cytotoxic NK cell subset mean 3.72%±1.4, range 1.4–6.1%, regulatory NK cell subset mean 0.95%±0.5, range 0.2–1.73%) as compared to seronegative controls [cytotoxic NK cell subset mean 10.06%±2.4, range 5.2–14.6%, (p=0.0001); regulatory NK cell subset mean 2.67%±1.1, range 1–4.9%, (p=0.0001)] and individuals with slow disease progression [cytotoxic NK cell subset mean 6.62%±3.2, range 0.9–10.3% (p=0.0193); regulatory NK cell subset mean 2.37%±1.2, range 0.8–4.2% (p=0.0016)] (Fig. 2B and C). The percentages of cytotoxic NK cells were significantly reduced in slow progressors as compared to healthy controls, however the regulatory NK cell percentages in both these groups were similar, indicating the reduction in cytotoxic NK cells in early HIV infection might be permanent, although the findings reflect the percentages from the cross-sectional study. The reduction in cytotoxic NK cell percentages has also been reported in other studies in acute HIV infection 12,14,15,19 and chronic HIV infection. 9,13,15
The percentage of defective NK cell subsets (CD56−CD16+) was increased significantly in recent HIV infection (mean 4.07%±1.2, range 2–5.7%, p=0.0001) as compared to seronegative controls (mean 1.97%±0.7, range 0.6–3.2%) and slow progressors (mean 2.45%±1.73, range 0.45–5.67%, p=0.0135) (Fig. 2D). The slow progressors also showed increased percentages of defective NK cells, but the difference was not significant when compared with those obtained from healthy controls (p=0.662) (Fig. 2D).
Furthermore, the total number of NK cells (spearman r=−0.7391, p=0.006) and the cytotoxic NK cell subset (Spearman r=−0.7881, p=0.002) showed a significant negative correlation with PVL set point, whereas the defective NK cell subset showed a significant positive correlation with PVL set point (r=0.7040, p=0.010) (Fig. 2D). The PVL set point is the marker of disease progression. Hence, the failure of defective NK cells in the early clearance of HIV-infected cells might be responsible for the increase in PVL set point, and, in addition, for faster disease progression. Previously it has been shown that the defective NK cells appear in chronic HIV infection and are a consequence of high viral load. Alter et al. 19 showed that the defective NK cells are expanded after the period of acute HIV infection and the CD56dimCD16− cells are depleted during the further course of disease. We confirmed this observation and showed that the presence of defective NK cells is seen within 6 months after acquiring HIV infection and could be responsible for the higher PVL set point and disease progression. The estimation of the defective NK cell percentage might be important for better characterization of the clinical stage of HIV infection and also to identify the patients with faster disease progression.
Since the defective NK cells were found to be the major population in the early phase of HIV infection, we tried to explore the role of CD161 expression on defective NK cells in early HIV infection. CD161 expression was estimated on all three NK cell subsets separately. Representative histograms of CD161 expression on the cytotoxic NK cell subset (CD3−CD16+CD56+), regulatory NK cell subset (CD3−CD16−CD56+), and defective NK cell subset (CD3-CD16+CD56−) from recent HIV infection, slow progressor, and seronegative control are shown in Fig. 3A, B, and C, respectively.

Representative histograms of CD161 expression level in NK cell subsets. CD161 expression in terms of MFI on cytotoxic NK cell subsets
The mean CD161 expression in cytotoxic and regulatory NK cell subsets did not differ significantly in all three groups; however, cytotoxic NK cells from healthy controls showed a higher expression of CD161 as compared to slow progressors and patients with recent HIV infection (Fig. 4A). The mean MFI of CD161 in the cytotoxic NK cell subset in recent HIV infection, slow progressors, and controls was 11.17 (range 3.0–29.3), 19.16 (range 4.2–32.6), and 18.95 (range 4.9–70.2), respectively. Whereas the mean of MFI of CD161 in the regulatory NK cell subset in recent HIV infection, slow progressors, and controls was 25.33 (range 12.7–57.9), 31.49 (range 13.4–68.2), and 33.02 (range 13.6–50.7), respectively. The defective NK cells from patients with recent HIV infection expressed significantly reduced expression of the CD161 molecule (mean MFI=13.90, range 6.7–25.8) as compared to the healthy controls (mean MFI=32.81, range 12.8–61.8) (p=0.0003) and slow progressor (mean MFI=27.45, range 15.1–55) (p=0.0009) (Fig. 3A). This reduced expression of CD161 on the defective NK cell subset showed a significant negative association with the PVL set point (r=−0.6154, p=0.0332) (Fig. 4B).
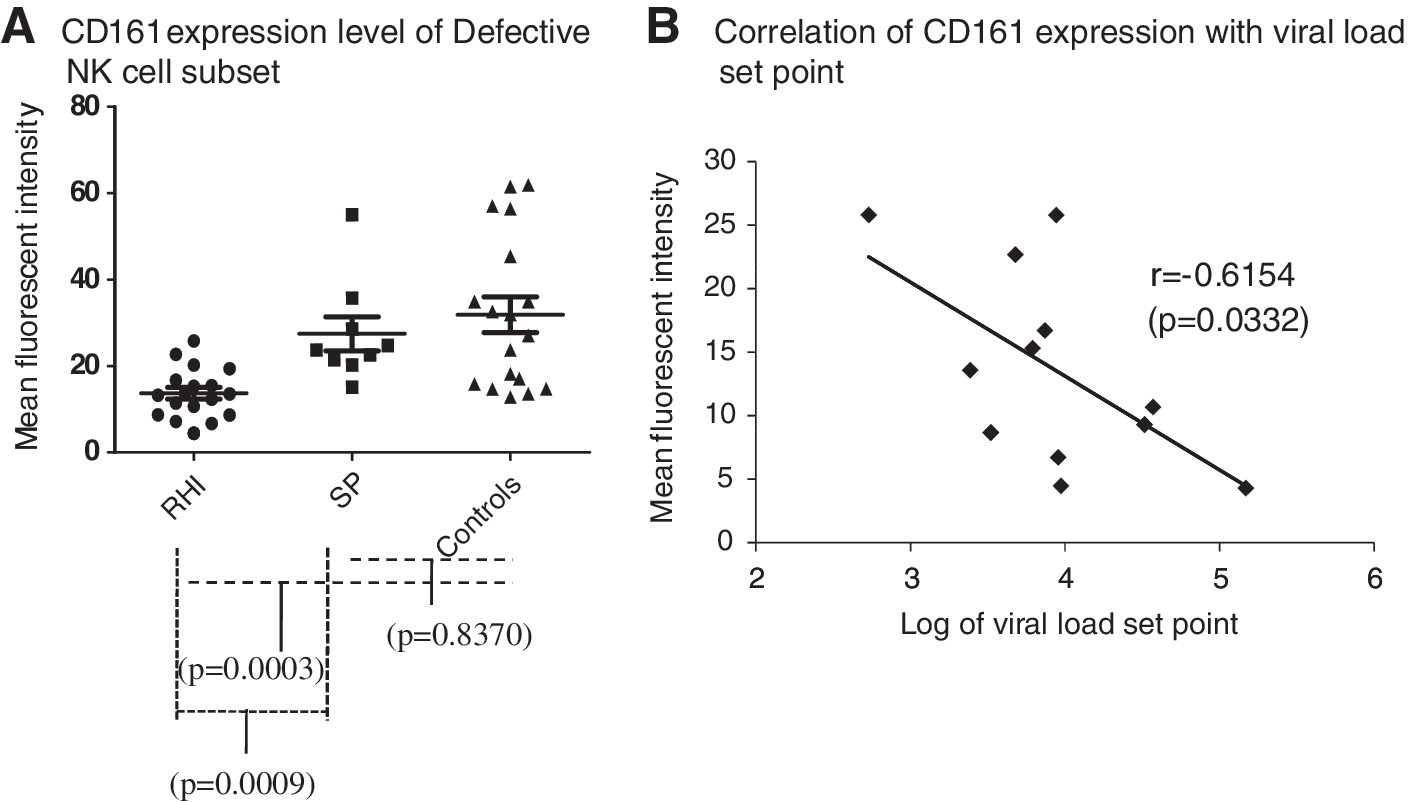
CD161 expression on defective NK cell subsets in the study groups. The CD161 expression, measured as mean fluorescence intensity (MFI) (y-axis) on defective NK cell subsets from patients with recent HIV infection (RHI), slow progressors (SP), and healthy HIV-seronegative individuals (controls), is shown in
CD161 has been found to be a major cell surface marker on NK T cells. 24 The signaling via CD161 involves alteration in sphingomyelin metabolism that could lead to cell apoptosis, differentiation, and proliferation. 23 Pozo et al. 23 showed that CD161 ligation in NK cells activates the acid sphingomyelinase pathway, which leads to ceramide production. Ceramide is known to act as a secondary messenger for various pathways for differentiation, proliferation, as well as apoptosis. The reduced expression of CD161 on the defective NK cells might be responsible for inhibiting the proliferation and maturation of these cells into cytotoxic/regulatory NK cells, in turn reducing the functional capacity of NK cells to lyse HIV-infected autologous CD4+ T cells and to secrete various cytokines. The reduced expression of CD161 in the early phase of HIV infection thus might be responsible for the failure of early virus control and the occurrence of high viral load set point. Indeed, CD161 expression was shown to influence the cytotoxicity of NK cells. 21,22 The intensity of CD161 staining on NK cells was higher in exposed but uninfected individuals than in HIV-infected individuals and controls. 25 Furthermore, following stimulation, CD161 was preferentially expressed by the activated CD107a+ NK cell subset. It has been shown that the binding of LLT1 (lectin like transcript 1), the cognate ligand of CD161, with CD161 reduces the cytolytic as well as IFN-γ-secreting ability of NK cells. CD161 expression can be unregulated by interleukin (IL)-12 supplement in vitro.
The interaction between CD161+ NK cells and LLT1 expressed by T lymphocytes in the early course of HIV infection might be responsible for the down-regulation of CD161 expression and thus further proliferation and differentiation of NK cells into cytotoxic and regulatory NK cells resulting in NK cell dysfunction and loss of control over viral multiplication.
To conclude, early HIV infection showed the presence of defective NK cells associated with a high PVL set point, indicating that the maker has utility in predicting disease progression and better clinical staging of HIV infection. The down-regulation of CD161, the proliferating and differentiating marker in the defective NK cells, might be suggestive of impaired differentiation of these cells into functional NK cells.
Footnotes
Acknowledgments
This study was supported primarily by a research grant from the Department of Biotechnology, Government of India. The authors thank the clinicians, counselors, and staff of NARI Clinics for primary screening and enrollment of patients. The authors also thank Dr. Smita Kulkarni and the staff of the Department of Virology for providing viral load, the staff of the Department of Immunology for providing CD4 T cell counts, and Dr. Arun Risbud and the staff of the Department of Serology for providing HIV diagnosis for the study subjects. Archana is supported by a Junior Research Fellowship from the Indian Council of Medical Research, Government of India.
Author Disclosure Statement
No competing financial interests exist.