
Abstract
The aim of the study was to investigate the prevalence and persistence of syncytium-inducing (SI) strains in HIV-1-infected children along time of infection and to evaluate the influence of antiretroviral therapy and host factors on viral tropism. This is a retrospective analysis carried out in 267 HIV-1 vertically infected children from an Argentinean cohort. The viral phenotype was screened in MT-2 cells and coreceptor usage confirmed by the GHOST cell assay. Also, CD4+ T cell count, viral load, antiretroviral therapy, and human CCR5-Δ32 and CCR2-64I genotypes were analyzed. A high frequency of HIV-1 SI/CXCR4-using variants (22%) was found among children within the first trimester of life, reaching 46% after 10 years of infection. At acute infection, zidovudine prophylaxis did not significantly affect the proportions of SI HIV-1 strains, while their presence was favored by the CCR5+/Δ32 genotype. Interestingly, the majority of the early SI strains did not persist over time, probably due to a higher susceptibility to antiretroviral (ARV) treatment or immunologic pressure. At the chronic stage, SI variants emerged even in the presence of HAART reaching 36% at 120 months of infection. Also the HIV-1 SI phenotype was associated with lower CD4+ T cell counts all along the course of infection. These findings highlight the need to evaluate the presence of SI/CXCR4 variants early at primary infection. This will make it possible to optimize the use of CCR5 inhibitors in children who are apparently carriers of the R5 virus preventing early therapeutic failure due to the reemergence of SI strains from reservoirs.
Introduction
V
HIV-1 natural infection in children differs from that of adults. In infants, time to AIDS has a bimodal distribution. Around one-fourth to one-third of infected children show a more aggressive infection, which progresses to AIDS within the first 2 years of life, probably due to an immature immune system at the time of infection. 7,8 The other three-fourths of infected children have a disease progression similar to that of adults. Shortly after transmission, most infants are infected with NSI/R5 variants. 9 –12
We have previously observed that the presence of HIV-1 SI variants at acute infection, with rapid replication capability, is indicative of a poor prognosis favoring a rapid onset of AIDS in vertically infected children. 13 The presence of SI/CXCR4-using viruses at an early stage of vertical transmission may be particularly relevant in pediatric immunopathogenesis, specially at birth due to an extremely active thymus with an unusual high number of potential target cells. 14 –16
Other factors such as antiretroviral (ARV) therapy and host genotype related to coreceptor usage may affect viral tropism. To date, their influence on the viral phenotype in infected children and adults remains controversial and information is scarce. 17 –21
The aims of our study were to investigate the prevalence and persistence of SI strains in HIV-1-infected children along the time of infection, and to evaluate the effect of ARV therapy and host genotype on HIV-1 variants with different tropism.
Materials and Methods
Study population
A retrospective evaluation of HIV-1 tropism was performed in 267 HIV-1 vertically infected children, born between 1986 and 2002, admitted at the “Garrahan Pediatric Hospital,” Buenos Aires, Argentina. A total of 107 infants were studied at primary infection (≤12 months of age) and 160 at the chronic stage (>12 months old). No information was available to distinguish between intrauterine, during delivery, and/or breast-feeding transmission.
At acute infection, information on zidovudine (ZDV) prophylaxis according to the AIDS Clinical Trials Group (ACTG) 076 protocol 22 was available in 88 of the 107 children. Forty-three infants received ZDV prophylaxis without breast-feeding as follows: (1) in 16 mother–infant pairs, the mothers received ZDV at pregnancy (last trimester) and at delivery, and the baby for the first 6 weeks of life; (2) in 10 pairs, the mother received ZDV at delivery and the infant during the first 6 weeks of life; and (3) in 17 cases, only the baby received ZDV for 6 weeks after birth. Forty-five infants did not receive prophylactic ARV therapy, and of them, 18 were breast-fed. The first HIV-1 phenotype determination was performed before starting HAART in 88 infants. At the chronic stage (>12 months old), 160 HIV-1-infected children were studied, and at the time of HIV-1 phenotype determination, 35 were naïve and 125 were receiving HAART [median time of 31 months, inter quartile range (IQR)=13–56 months].
For phenotypic switch studies, 10 patients with SI variants at acute infection were prospectively evaluated, with a median follow up time of 25 months (IQR:13.5–37 months). Informed consent from the parents or legal guardians was obtained. The study was reviewed and approved by the Ethics Committee of the Hospital.
Isolation of HIV-1 from peripheral blood mononuclear cell cocultures
HIV-1 was isolated by cocultivation of cells as previously described by the AIDS Clinical Trials Group. 23 Briefly, peripheral blood mononuclear cells (PBMCs) from both the patient and HIV-1-seronegative blood donors prestimulated for 24–72 hours with 5 μg/ml of phytohemagglutinin (PHA) (Difco Laboratories) were cocultured at a final concentration of 2×106 cells/ml. Cocultures were maintained for 28 days in RPMI 1640 medium (Gibco BRL, Invitrogen) supplemented with 20% heat inactivated fetal bovine serum (FBS), 5 U/ml interleukin (IL)-2 (Sigma Aldrich), and 10 μg/ml gentamicin (Gibco BRL Invitrogen). Measurement of HIV-1 p24 Ag of coculture supernatants was performed with a commercial assay kit (Vironostika HIV-1 Antigen, BioMérieux).
Determination of syncytium-inducing viral isolates using MT-2 cells
The SI assay was performed as described by Japour et al. 24 In 96-well plates, supernatants from HIV-1 positive cocultures were added to four wells containing 5×104 MT-2 cells/well in RPMI 1640 supplemented with 10% FBS and 10 μg/ml gentamicin. SI primary viral isolates and a CXCR4-tropic HIV-1LAI/PBMC laboratory adapted strain (from NIH AIDS Reference and Reagent Program) were used as positive controls and wells without virus and with NSI isolates were considered as negative controls. Syncytium formation of MT-2 cells was examined under a microscope every 3 days for 28 days. To confirm the infection of MT-2 cells, at the end of the culture, HIV-1 proviral DNA of MT-2 cell lysates was amplified directly, as described by Albert and Fenyo. 25 To ensure infectivity of the viral inoculum, a parallel control assay was performed adding the viral inoculum in wells containing 2×105 PHA-stimulated PBMCs from HIV-1-seronegative donors. Supernatants from all the wells were measured for HIV-1 p24 Ag. When the viral inoculum infected PBMCs but was unable to induce syncytium in MT-2 cells, the viral isolate was scored as NSI.
Determination of coreceptor usage
Viral infection was performed as described by Vodrös and Fenyo. 26 GHOST cell lines transfected with CD4+ and viral coreceptors (kindly provided by Dr. H. Salomon, Centro Nacional de Referencia de SIDA) were maintained in DMEM medium (Hyclone, Biodynamics) containing 10% FBS and antibiotics. One day before infection, 24-well plates were prepared with 1.5×105 cells/well, followed by infection with virus standardized for TCID50. Cultures were observed under a fluorescent microscope and p24 Ag was determined. In all cases, CCR5-tropic (HIV-1BaL) and CXCR4-tropic (HIV-1LAI/PBMC) laboratory adapted strains were used as control of coreceptor usage. CXCR4 using was confirmed by the inhibition of viral infection with AMD3100 (Sigma Aldrich), 10 μg/ml, a CXCR4 inhibitor.
CCR5 and CCR2 genotyping
Of the 267 patients, 259 were genotyped for CCR5-Δ32 and 246 for CCR2-64I by RFLP-PCR, as previously described. 27
Clinical parameters
In 240 children, CD4+ T cell counts were measured using flow cytometry (FACSort; Becton Dickinson, CA). The data were collected close (<6 months) to the time of the phenotypic study. Clinical and immunological stages were determined according to the 1994 criteria of the U.S. Centers for Disease Control and Prevention (CDC) classification for children. 28
Viral load measurements
Plasma viral load was determined by the HIV-1 RNA QT Nuclisens (Organon Teknika, Boxtel, Netherlands), Amplicor HIV-1 Monitor test, or COBAS AmpliPrep/COBAS TaqMan HIV-1 Test (Roche Diagnostic Systems, Branchburg, USA), depending on the kit available at the time of measurement.
Statistical analysis
The effects of age, ZDV prophylaxis, HAART, CCR5-Δ32, and CCR2-64I on the HIV-1 phenotype were tested with logistic regression analysis. Group contrasts for CD4+ T cells percentage/absolute counts and viral load were evaluated with the Mann–Whitney test. In all cases, two-tailed tests were performed with a significance level of 0.05.
Results
Proportion of SI and NSI HIV-1 variants in children at different times of infection
HIV-1 isolates from 267 vertically infected patients, ranging from 1 to 220 months of age, were evaluated for SI capability in MT-2 cells. The viral phenotype was determined once in 170 children, and more than once (2–9 times) in the other 97 infants. Children were stratified according to age (time of infection). The frequency of SI HIV-1 isolates varied with time of infection, being 22% within the first 3 months after birth, with a sustained drop until 12 months of age (Fig. 1). Afterward, it was followed by an increase, reaching 46% of SI variants after 10 years of infection.
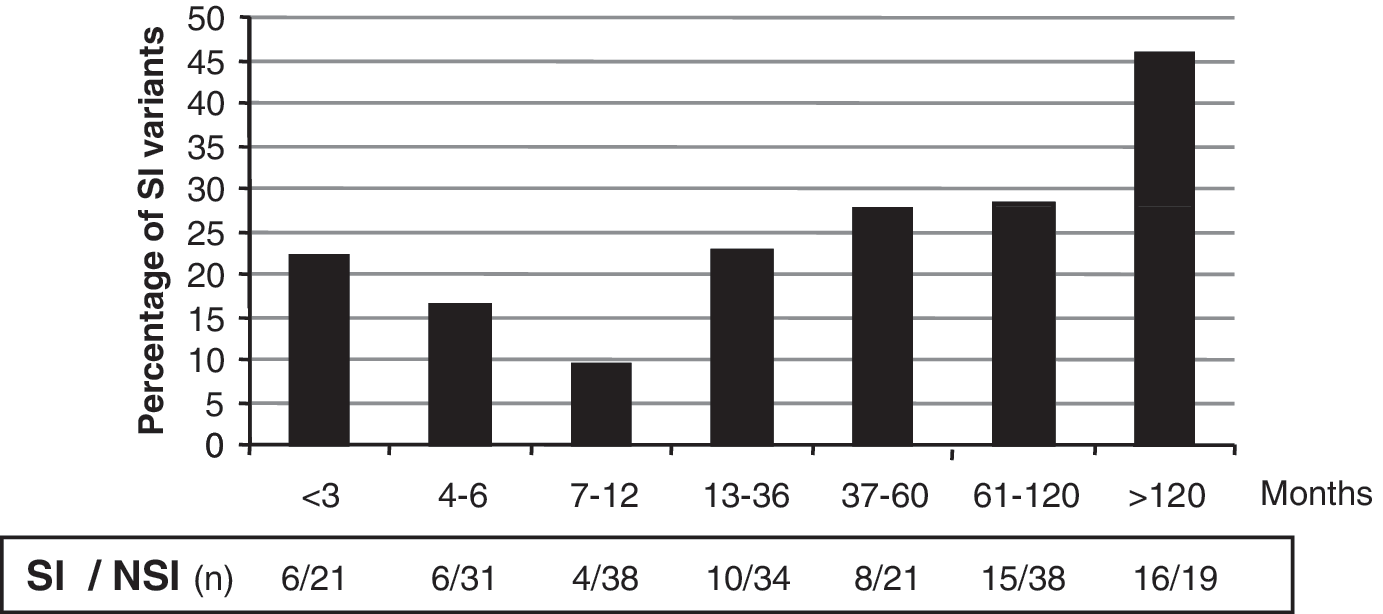
Distribution of HIV-1 phenotype from vertically infected children at different ages or times of infection. Filled columns are syncytium-inducing (SI) variants and n=the number of patients SI/non-syncytium-inducing (NSI). A patient with SI viral isolates was plotted as SI at the age observed; when a patient had more phenotypic characterizations as NSI during follow-up, we considered the first one.
To confirm CXCR4 coreceptor usage, 35 SI isolates derived from MT-2 cultures were tested in GHOST transfected cell lines. Of them, 33 were R5X4, and the remaining 2 SI MT-2 isolates were able to infect the GHOST cells only through the CCR5 coreceptor. The absence of infection in CXCR4 GHOST cells of the two latter SI isolates could be due to a higher sensitivity of the MT-2 assay (which expresses only the CXCR4 coreceptor) for detection of low levels of SI strains, 29 based on different levels of expression of coreceptors between MT-2 and GHOST transfected cell lines. 30
In addition, viral isolates from 10 children of our cohort were simultaneously tested by Trofile and MT-2 assays with a full concordance of both methods (all 10 SI strains were X4R5 dual/mix by Trofile).
Effect of ARV drugs on viral tropism
We next analyzed whether ARV prophylaxis/treatment might have influenced the selection of SI variants at different stages of infection. Thus, we divided the patients into acute and chronic stages. Viral isolates from a total of 88 infected infants under 12 months old at acute infection were studied comparing those who had received ZDV as preventive treatment (n=43) and those who had not (n=45). During the first 6 months of life, the percentage of infants with SI variants did not differ regardless of having received preventive therapy or not, indicating that, at early stages of infection, preventive ZDV did not affect the viral phenotype (Table 1). Between 6 and 12 months of age, SI variants were absent in infants who had previously received ZDV prophylaxis, although the association was not statistically significant (p=0.9).
Patients ≤12 months old include infants who had received zidovudine exclusively for 6 weeks, and in patients older than 12 months, at a chronic stage of infection, children receiving HAART.
Age-stratified logistic regression.
ARV, antiretroviral; NSI, non-syncytium-inducing; SI, syncytium-inducing.
We further evaluated the impact of HAART on the viral phenotype in a total of 160 children older than 1 year. At the moment of study, 125 children were receiving HAART and 35 were naïve of ARV (due to a delay in HIV diagnosis). Because age can be a confounding factor the children were subdivided into two groups: ≤60 and >60 months old, following PAHO guidelines, 31 which consider children older than 60 months immunologically similar to adults. The proportion of SI variants was significantly higher in patients under therapy than in those who did not receive any ARV (p=0.015), independently of the age stratum (Table 1). The marked reduction of SI variants in naïve children older than 12 months old may be due to a shorter asymptomatic period than those carrying NSI variants.
Likewise, the association between HAART and SI phenotype in children at the chronic stage was significant after correcting for a monotone effect of age, immunological stage, and viral load (p=0.019, data not shown).
SI to NSI phenotype switch at early stages of infection
We further investigated the effect of ARV therapy on the HIV-1 SI to NSI phenotype switch in the acute stage. Consecutive viral tropism was evaluated in 10 patients before and close to initiation of ARV treatment (Table 2). Two viral tropism determinations were performed in infants #354 and #579 prior to the start of therapy. In both cases, the viral phenotype spontaneously switched from SI to NSI at 3 and 29 months, respectively. Moreover, infant #579 preserved the same NSI tropism after 34 months of ARV therapy with one log10 reduction of viral load. In the other six infants, the initial SI variants switched to NSI as early as 3 months after initiation of HAART, with a mean of 12.3 months under ARV. In the remaining two infants (#75 and #545), SI variants persisted despite the treatment. Infant #75 remained SI and died at 18 months of age, with a severe immunodeficiency. Paradoxically, in patient #545, the SI variants detected at 5 months of age did not switch after initiation of HAART, with persistent high viral loads, normal CD4+ T cells, and free of AIDS up to 121 months. These observations indicate that the majority of the SI variants are susceptible to phenotype switch either spontaneously or by HAART during acute pediatric infection.
Patient 75 died at 18 months of age.
ZDV, zidovudine; ARV, antiretroviral; NSI, non-syncytium-inducing; SI, syncytium-inducing; ND, not determined; ddI, didanosine; NFV, nelfinavir; 3TC, lamivudine; d4T, stavudine; EFV, efavirenz; RTV, ritonavir; ABC, abacavir; KLT, lopinavir-ritonivir.
Association of HIV-1 tropism and clinical parameters
We next explored whether the HIV-1 phenotype is associated with clinical parameters, and whether it may differ between the acute and chronic stages of infection. In infants younger than 12 months of age, viral loads were extremely high regardless of the viral phenotype. The median for patients with SI viruses was 5.55 log10 copies/ml and for NSI variants was 5.64 log10 copies/ml. However, at chronic infection, the viral loads were higher in children with SI variants than in those with NSI isolates (p=0.026) (Table 3). Evaluation of the viral phenotype in association with immunological parameters showed that children under 12 months harboring NSI strains had higher CD4+ T cells in percentage and absolute counts than those carrying the SI variants.
Therefore, at early and chronic stages of infection, the SI phenotype was significantly associated with lower CD4+ T cell counts (p=0.010). Furthermore, different stages of immunodeficiency were evaluated by a logistic regression analysis confirming that the SI phenotype is strongly correlated with severe immunodeficiency (p≤0.001). These findings indicate that along the whole course of infection the viral tropism is associated with the immunodeficiency of the patients.
Association between CCR5-Δ32, CCR2-V64I polymorphisms, and HIV-1 tropism
Since CCR5-Δ32 heterozygosity may contribute to a selective pressure for NSI/R5 viruses to alternatively use CXCR4 as a coreceptor, we evaluated its influence on the frequency of HIV-1 SI isolates at different stages of infection (Table 3). At primary infection, we observed that SI isolates were significantly more frequent in CCR5+/Δ32 heterozygous children than in CCR5+/+ homozygotes (60% vs. 14%, respectively, p=0.028). However, this association was not observed in children at chronic infection.
The evaluation of CCR2-V64I polymorphism revealed no association with viral tropism at either stage of infection (Table 3). These results indicate that at early stages of HIV-1 infection acquired by perinatal transmission, children heterozygous for the 32-bp deletion in the CCR5 allele are more prone to harbor SI variants.
Discussion
In the present study, we observed a high proportion of SI/CXCR4-using variants at primary infection in HIV-1 vertically infected children, apparently not affected by ZDV prophylaxis. In acute infection, SI variants can switch to NSI spontaneously or after initiation of HAART. However, HAART was not able to prevent the late reemergence of SI strains. In addition, the CCR5+/Δ32 genotype favored the early presence of SI variants, but had no influence on the appearance of SI strains at the late stage of infection.
Several studies have indicated that most infants at the acute stage are infected with NSI/R5 viruses. 9 –12 Notably, in our study, a high proportion of vertically infected children carried SI variants within the first months of life. The presence of SI variants, at as early as 1 month of age, suggests a rapid switch from NSI to SI viruses after transmission, 32 or a direct perinatal transmission of SI/CXCR4-using variants. Huang et al. 33 and Salvatori et al. 10 have documented that viruses with different phenotypes, including CXCR4-using viruses, are potentially transmissible from the mother to the infant. Unfortunately, in our study, samples from the mothers were not available.
Therefore, we could not determine whether SI variants were transmitted or suffered a premature phenotypic switch. However, these two possible explanations are not mutually exclusive. The infection with SI variants can also be a common event for other routes of transmission. In recent studies in adult cohorts, including HIV-1 seroconverters and naïve patients, CXCR4-using variants were found in proportions as high as 20%. 34 –36 The high proportion of SI viruses at the early stage of infection might be due to a preferential selection of CXCR4-using variants by trophoblast cells, favoring their passage in utero. 10,37 Likewise, in the newborn, other conditions, such as the predominant expression of the CXCR4 coreceptor in thymocytes, and detectable CXCR4 in neonatal PBMCs and monocytes, with very low levels of CCR5, may promote the selection of SI variants. 38 –40 Furthermore, CCR5 expression in immune cells varies with age, being undetectable at birth, 41,42 creating a potential beneficial scenario for the replication of SI variants.
Another relevant finding was that the SI viral phenotype at the early stage of infection was more common among CCR5+/Δ32 heterozygotes than among CCR5+/+ homozygotes. However, this association was not found at the chronic stage. These observations suggest that CCR5+/Δ32 heterozygosity could act as a selective pressure at primary infection, favoring transmission of CXCR4-using variants. 43
In contrast to our results, different investigators did not find SI variants during the first year of infection, even in symptomatic children. 44,45 These discrepancies can be attributable to regional HIV-1 subtype bias and/or to the sensitivity of the methods used. It has been suggested that different HIV-1 subtypes can vary in their ability to use the CCR5 or CXCR4 coreceptors. Among HIV-1 group M viruses, subtype D and CRF01_AE have shown a higher prevalence of CXCR4-using variants when compared to other subtypes, 46,47 although these differences are controversial. 48,49 In our population, BF recombinants represent almost 85% of the circulating strains, 50 but we found no evidence of a preferential coreceptor usage between BF and B subtypes (unpublished data). Therefore, the higher frequency of SI variants at acute stages of infection seems to be independent of HIV-1 subtype in our group of vertically infected children.
On the other hand, different methods for phenotype determinations show broad sensitivity ranges. Current assays of viral tropism include two versions of the MT-2 assay (PBMC and MT-2 coculture or MT-2 infected with viral stocks), transfected cell lines, and the algorithm from the V3 loop of the env gene. 51 –53 It has been reported that the MT-2 assay used (with viral stocks) has a 98% concordance with the Trofile assay, 29 which is the only tropism test approved for screening the phenotypic variants prior to administration of CCR5 antagonists. In agreement, a complete concordance between the Trofile and MT-2 assays was observed in our group of patients tested.
Furthermore, we found that ZDV as preventive therapy did not affect the presence of SI variants after birth. Nevertheless, the absence of SI variants after 6 months of age in children with ARV prophylaxis was noteworthy. Although we found no statistical differences as compared with children without ZDV prophylaxis, the small number of patients of this age stratum did not make it possible to discard a difference with statistical power above 0.8 (the desired sample should have n>79 for the observed side effect). The fact that early circulating SI/CXCR4-using variants can spontaneously disappear suggests that they have a higher susceptibility than NSI/R5 strains to immune control, allowing NSI/R5 variants to emerge and predominate in the infection. 54,55
It should be noted that at chronic stages of infection, only 35 of 160 children persisted without treatment because of a delayed diagnosis. The majority of these asymptomatic naïve patients carried NSI strains. Probably, children who are carriers of SI variants soon after birth are more frequently symptomatic and thus diagnosed and treated at an earlier stage of infection. A different scenario was observed in children under HAART with a higher prevalence of SI variants consistent with data on adult cohorts. 17,56 indicating that HAART did not prevent the appearance of SI strains at late stages of infection. The SI emergence may be related to a reduced expression of CCR5 in patients under treatment, 57,58 and/or to an increase in naïve CD4+ T cell subsets, with high levels of CXCR4. 59
As previously described, 1,60 we observed a strong association between SI tropism and immunodeficiency regardless of the time of infection. However, according to different authors, the immune damage by SI/CXCR4-using viruses may differ with the time of infection. At primary infection, SI strains can directly affect thymic function 16 with a decrease in the number of CD4+ T cells, while at late stages the impairment can be mainly at the level of secondary lymphoid tissue. Further studies are needed to elucidate this issue.
These findings highlight the need to evaluate the presence of SI/CXCR4 variants at primary infection. These observations may have important clinical and therapeutic implications, especially during early HIV-1 infection, since the phenotypic determination at the acute stage is crucial to provide a more effective and accurate treatment. This will make it possible to optimize the use of CCR5 inhibitors and prevent early therapeutic failure in children who are apparently carriers of the R5 virus due to the reemergence of SI strains from reservoirs.
Footnotes
Acknowledgments
We thank Ms. Natalia Beltramone, Mrs. Carmen Gálvez, and Mrs. Silvia Marino for their technical assistance. Cintia Crudeli executed the laboratory work; Paula Aulicino, Carlos Rocco, and Andrea Mangano actively participated in discussions and critical reading; Carlos Rocco performed the statistical analyses; Rosa Bologna is the infectious disease specialist and performed the follow-up of the pediatric cohort; and Luisa Sen directed the investigations. All authors contributed to the writing and revision of the manuscript. We acknowledge the NIH AIDS Research and Reference Reagent Program for providing the HIV-1LAI/PBMC strain used in this work. This study was partially supported by the Argentine Awards from Agencia Nacional de Promoción Científica y Tecnológica (PICT 2004-25830) and from Consejo Nacional de Investigaciones Científicas y Técnicas (PIP 6057).
Author Disclosure Statement
No competing financial interests exist.