
Abstract
Little is known about the extent and predictors of polymorphisms potentially influencing susceptibility to HIV integrase inhibitors. HIV-1 genetic diversity and drug resistance are major challenges in patient management globally. To evaluate HIV genetic diversity and drug resistance-associated mutations within a Nairobi cohort, genetic analysis of the HIV-1 pol-integrase gene regions was performed on samples collected from 42 subjects and 113 Kenyan intergrase sequences deposited in the Los Alamos HIV database. From the partial pol-integrase sequences analyzed phylogenetically, it was shown that 26 (61.9%) were subtype A1, 9 (21.4%) were subtype D, 5 (11.9%) were subtype C, 1 (2.4%) was subtype A2 and 1 (2.4%) was subtype CRF. Integrase-associated mutations were found in 12 patients, and in all 113 sequences already deposited in GenBank. One sample from this study and five from previous Kenyan integrase sequences had mutations at T97A, which is associated with reduced susceptibility to raltegravir.
HIV-1
Studies have shown that HIV-1 genetic diversity, including the existence of distinct genetic subtypes and the evolution of drug-resistant genotypes, can greatly affect the diagnosis and treatment of HIV-1 infection. 4 The continued development of novel antiretroviral agents for the treatment of HIV-1 has most recently culminated in the introduction of a new therapeutic class of drugs, the strand transfer integrase inhibitors (InSTI). The drug raltegravir (RAL; MK-0518) has demonstrated excellent efficacy in both treatment-experienced and drug-naive patients 5 and is the first InSTI to be approved for use in treatment-experienced patients in the developed world. 6 Elvitegravir (EVG; GS-9137) has also shown potent antiviral l activity both in vitro and in vivo, 7 and phase III clinical trials are underway. 6
InSTI act by targeting the integrase protein. Integrase has two catalytic functions; first it removes a dinucleotide from each 3′ end of viral DNA (the 3′ processing reaction), and second, it mediates the transfer of the proviral DNA strand in the host nucleus and covalently links the 3′ ends into the host DNA (the strand transfer reaction). 8 These steps create the provirus, a state that, at once, achieves transcriptional competency for the retrovirus and provides for the stable maintenance and integrity of the viral genome throughout the lifespan of the infected cell and for all subsequent daughter cells. 6 All current compounds in clinical development preferentially target the strand transfer reaction of integrase, despite representing distinct chemical classes. 7
Antiretroviral therapy (ART) has sharply reduced HIV transmission, morbidity, and mortality in developed countries, but it has created the long-term spectre of drug resistance. Widespread use of ART in these countries has resulted in an increased prevalence of drug-resistant variants, ranging from 10% to 20% among drug-naive patients. 9 However, new drugs continue to be developed for the treatment of HIV despite the threat of increasing drug resistance.
With the introduction of integrase inhibitors as viable clinical candidates, we sought to examine the integrase coding region from drug-experienced and drug-naive patients within a Nairobi cohort with the aim of determining the prevalence of mutations associated with InSTI.
Five-milliliter blood samples were collected from 42 subjects with informed consent, as well as with anonymous epidemiological data, including age, sex, ARVs status, and nationality, from different health facilities in the city of Nairobi, Kenya (Table 1). The samples were confirmed to be positive for HIV-1 antibodies using a rapid detection kit (Determine HIV1/2; Abbot, Japan) and Bioline HIV1/2 (Republic of Korea) and CD4 counts were determined.
HIV-1 DNA was extracted from PBMCs using the Qiagen proviral QIAamp DNA Mini kit following the manufacturer's instructions. A part of the HIV-1 pol gene covering 288 bp of the integrase (corresponding to 4493–4780 nt in HIV-1 HXB2) was amplified by nested PCR using primers Unipol5 (5′-TGGGTACCAGCACACAAAGGAATAGGAGGAAA-3′) and Unipol6 (5′-CCACAGCTGATCTCTGGCCTTCTCTGTAATAGACC-3′) in the first round and Unipol1 (5′-AGTGGATTCATAGAAAGCAGAAGT-3′) and Unipol2 (5′-CCCCTATTCCTTCCCCTTCTTTTAAAA-3′) in the second round. Amplification conditions were similar, with a hot start at 95°C followed by 35 cycles of denaturation at 95°C for 30 s, annealing at 45°C for 30 s, and extension at 72°C for 1 min, and a final extension of 72°C for 7 min. 10
Amplified products were confirmed by visualisation using ethidium bromide-stained agarose gels. The amplified products were sequenced directly with second PCR primers of the integrase region using an automated ABI 310 sequencer (Applied Biosystems, Foster City, CA) as per the manufacturer's instructions. 9
The 42 generated sequences from the study were pooled together with 113 Kenyan integrase sequences from the Los Alamos database accessed on June 1, 2011 (Supplementary Table S1; Supplementary Data are available online at
In addition, neighbour-joining phylogenetic trees, including reference sequences from the Los Alamos database, were constructed using Clustal W (version 1.83) and inferred using Tree View (version 1.6.6; Institute of Biochemical and Life Sciences, Scotland, United Kingdom). Bootstrap resampling (1000 data sets) of multiple alignments was performed to test the statistical robustness of the trees (see Fig. 1).
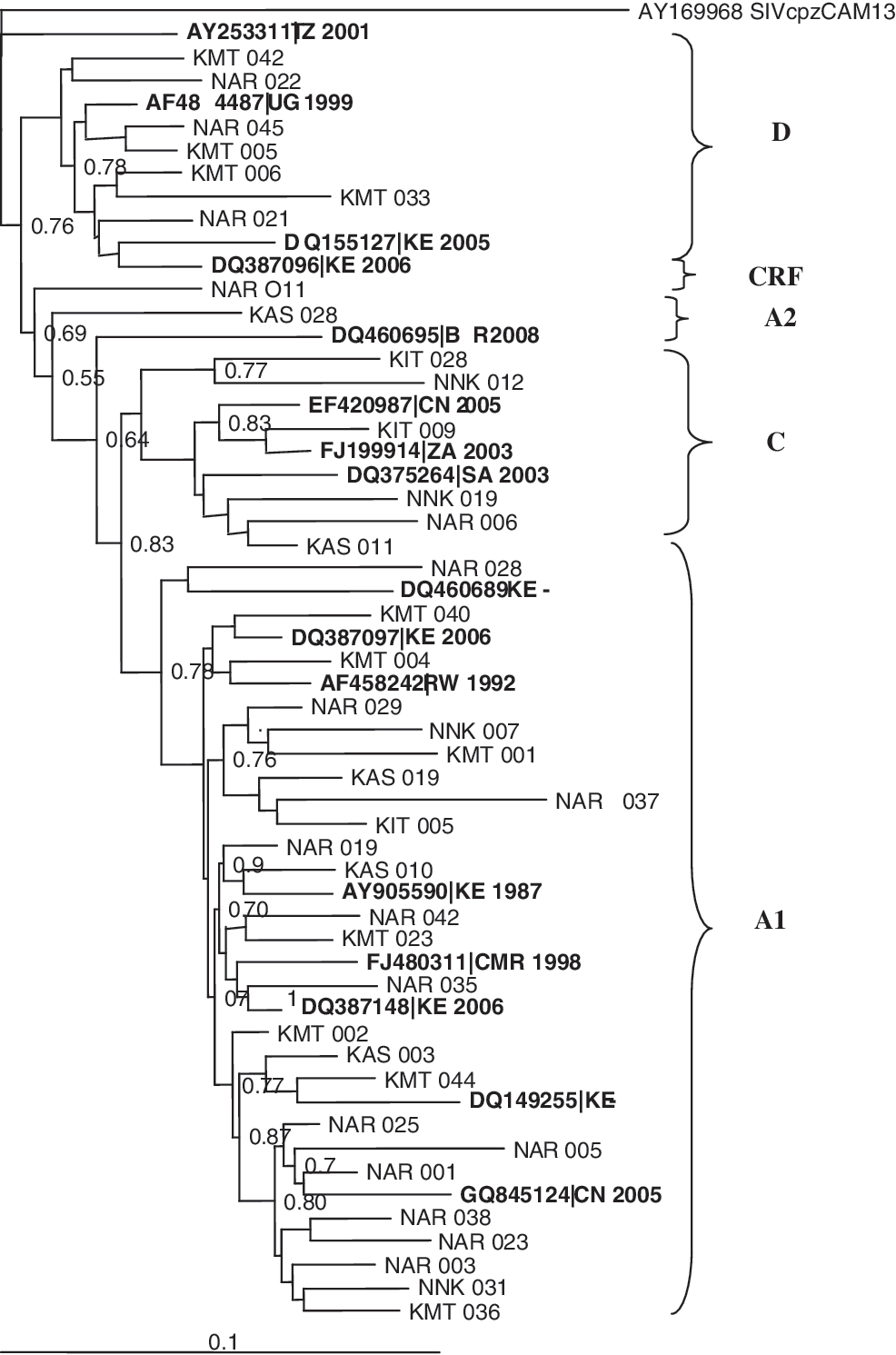
Phylogenetic tree of the HIV-1 pol-integrase region in relation to reference sequences from the Los Alamos HIV database (boldface).
In this study, no primary mutations (Y143R=C=H, Q148H=R=K, and N155H=S) associated with reduced susceptibility to the integrase inhibitors raltegravir and elvitegravir were detected. However, 4.5% of the Kenyan strains had the T97A mutation, which is associated with a reduced susceptibility to integrase inhibitors, raltegravir and elvitegravir. In addition, 80.6% of the studied sequences had secondary mutations (see Table 2). The secondary mutations might have been due to natural polymorphisms that occur in the HIV genome.
The majority of the sample sequences analyzed were found to belong to subtype A1: 26 (61.9%), followed by D: 9 (21.4%), C: 5 (11.9%), A2: 1 (2.4%), and CRF: 1 (2.4%) (Fig. 1). This study agrees with previous molecular epidemiological studies on the predominance of HIV-1 subtype A1 in Kenya. 10,13 –15 The HIV-1 subtype A1 samples clustered with reference sequences mainly from Kenya showing that the viruses were probably of Kenyan origin. The samples that were HIV-1 subtype C clustered with reference sequences from South Africa, Zambia, and Kenya. The HIV-1 subtype D sequences clustered with reference sequences from Uganda, Tanzania, and Kenya. 9,10 This study not only described the distribution of HIV-1 strains in a Nairobi cohort in Kenya, but also evaluated drug resistance mutations on the new class of InSTI prior to treatment.
The Kenya AIDS indicator survey of 2007 showed that of the estimated 392,000 Kenyan adults in need of ART, 138,000 (35%) had received treatment by September 2007. 11 This figure increased to 212,000 (54%) by June 2008. 11 The increase in ART coverage is expected to lead to an increase in drug-resistant strains among drug-naive patients. In addition, stigma and cultural practises by different communities in Kenya may affect ART compliance resulting in an accelerated appearance of drug-resistant mutants, which are a potential source of transmitted drug resistance. 9
There is an evolving need for continuous development of antiretroviral drugs targeting different stages of the HIV replication cycle and infection. Development of antiretroviral drugs has been going on, an initiative supported by many governments worldwide, including Kenyan. Though new classes of drugs are being developed, development of drug resistance by the virus has always limited effective management of HIV patients. Nevertheless, the addition of integrase inhibitors to the armamentarium of antiviral agents is a considerable advance in the management of patients infected with HIV-1. 6
Despite documenting substantial numbers of polymorphisms in the integrase coding regions similar to that seen in other analyses, 6 this study did not identify any amino acid substitutions associated with high level resistance to the clinically relevant integrase inhibitors raltegravir and elvitegravir. 6 Although none of the major mutations associated with primary resistance to InSTI in clinical use was identified (T66I, E92Q, Y143R/C, Q148K/R/H, and N155H), the T97A mutation was detected. The prevalence of natural polymorphism of T97A mutations known to confer reduced susceptibility to raltegravir, an InSTI, appeared in 4.5% of the total sequences analyzed. This could be an indication that with an increase in drug pressure, the virus is bound to develop primary mutations known to lead to drug resistance to this class of drugs.
Secondary mutations associated with resistance to RAL, EGV, and/or other inhibitors, T97A, V201, and I203L, were detected in more than 5.3% of samples. The mutation 201I was detected in most samples (71%). In addition, other secondary mutations were detected (see Table 2). The detection of these natural polymorphisms suggested a possible adaptation of the virus due to immune or drug pressures, which may eventually occur even on the positions linked to drug resistance to raltegravir. This study confirms previous studies on HIV-1 integrase that have revealed that several secondary mutations associated with resistance to integrase inhibitors were frequently detected in drug-naive patients from many countries including Belgium, Brazil, the United States, and Thailand. 12 This suggests the need for phenotypic studies on integrase drug resistance studies.
This study shows that it is possible that the new class of integrase inhibitors can be used together with HAART in the management of HIV/AIDS patients. However, drug interaction studies need to be carried out to determine this. Taken together, we believe that these data support the current argument that integrase inhibitor-naive subjects are likely to benefit fully from the incorporation of the current clinically relevant agents into a treatment regimen, without concern of inherent reduced susceptibility.
It would appear that the integrase coding region contains as much natural variation as that of protease, but that mutations associated with high-level resistance to existing InSTI are rare, even if present in integrase-naive patients. Most of the highly prevalent polymorphisms have little effect on InSTI susceptibility in the absence of specific primary mutations. It must be emphasized that as the use of these potent agents expands, the potential for the transmission of InSTI-resistant viruses will increase and it is therefore critical to include integrase genotypic and phenotypic analyses in studies that focus on transmission of drug-resistant HIV-1 variants.
Sequence Data
The pol integrase sequences were deposited in GenBank under accession numbers HQ177046–HQ177063, HQ177068– HQ177069, HQ177071–HQ177088, HQ177090, and HQ177093.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.