
Abstract
There is an urgent global need for preventive strategies against HIV-1 infections. Llama heavy-chain antibody fragments (VHH) are a class of molecules recently described as potent cross-clade HIV-1 entry inhibitors. We studied the potential of a VHH-based microbicide in an application-oriented fashion. We show that VHH can be inexpensively produced in high amounts in the GRAS organism Saccharomyces cerevisiae, resulting in a very pure and endotoxin free product. VHH are very stable under conditions they might encounter during transport, storage, or use by women. We developed active formulations of VHH in aqueous gel and compressed and lyophilized tablets for controlled release from an intravaginal device. The release profile of the VHH from, e.g., a vaginal ring suggests sufficient bioavailability and protective concentration of the molecule at the mucosal site at the moment of the infection. The ex vivo penetration kinetics through human tissues show that the VHH diffuse into the mucosal layer and open the possibility to create a second defense layer either by blocking the HIV receptor binding sites or by blocking the receptors of immune cells in the mucosa. In conclusion, our data show that VHH have a high potential for HIV-1 microbicide application because of their low production costs, their high stability, and their favorable release and tissue penetration properties.
Introduction
With the HIV pandemic on the verge of entering its fourth decade, the need to develop effective and inexpensive preventive strategies remains an urgent global health priority. 1 Although there is promising progress in the development of a vaccine, 2 there is also growing evidence that vaginally administered HIV microbicides are effective in helping to reduce the incidence of sexually transmitted infections, which account for the majority of new infections in the developing world. 3,4 The recent positive outcome of the CAPRISA 004 trial has shown that it is possible to achieve a certain level of protection with a coitally dependent vaginal gel. 5 Most HIV microbicide candidates currently being evaluated in clinical studies are either nonspecific polymeric compounds formulated as vaginal gels and designed to prevent entry of the virus into host cells 6 –8 or potent small-molecule antiretroviral compounds that inhibit the viral replication cycle. 9 –11 However, the lack of specificity associated with polymeric microbicide candidates, coupled with clinical data suggesting a modest degree of protection at best, 12 and the concern over the potential for development of resistant strains with long-term use of a single antiretroviral agent have forced the field to consider alternative classes of microbicidal compounds.
A number of microbicide candidates are currently being evaluated for their potential as HIV inhibitors, including human antibodies b12 13 and a triple mix of monoclonal antibodies 2F5, 4E10, and 2G12. Although initial efficacy data in nonhuman primates has been encouraging, the high costs associated with production/synthesis, difficulties in developing stable formulations, and poor tissue permeability are major obstacles that need to be overcome in order to progress these biomolecular approaches as practical microbicidal strategies. 14
In addition to conventional immunoglobulins, the Camelidae family possesses a class of antibodies devoid of the light chain, named heavy-chain antibodies.
15
The variable domain of these antibodies (VHH) can be selected against almost any target molecule,
16
–18
cloned and efficiently produced by relatively easy and inexpensive methods.
19
–21
Previously, phage-displayed libraries of VHH isolated from llamas immunized with the glycoprotein gp120 from different HIV-1 clades (A,
The screening for clones with improved neutralization characteristics, targeting new epitopes, and the construction of multimeric and chimeric molecules continue and we now have a panel of more than 60 VHH with specific and characterized activity against different epitopes on gp120 and gp41. 23,24 Moreover, we are characterizing new families of VHH against the cell receptors CD4, CCR5, and DC-SIGN that, together with the recent discovery of CXCR4 binders, 25 open up the possibility of a double defensive layer based on virus binding and cell receptors binding. In this study, several important aspects that determine whether this class of molecules fulfills the requirements for topical HIV microbicides have been evaluated for the first time, including production, purification, thermal and pH stability, formulations, and mucosal permeability.
Materials and Methods
Production of VHH constructs by cell culture methods
Strains, vectors and cultivation media
Preparation of all the VHH constructs was carried out using Escherichia coli strain TG1 supE hsdΔ5 thi Δ(lac-proAB) F′[traD36 proAB+ lacIq lacZΔM15].
Saccharomyces cerevisiae strain VWk18 gal1 (CEN-PK102-3A, MATa, leu2-3, ura3, gal1::URA3, MAL-8, MAL3, SUC3) was used for the fermentations. Inocula, fermentation media and growth conditions were the same as previously described. 21 Plasmids pUR4547 and pUR4585 (Bio Affinity Company, Leiden, The Netherlands) were used as expression vectors for VHH genes in S. cerevisiae. pUR4547 is a 2-μm-based LEU2-marked plasmid suitable for GAL7 promoter-controlled expression of VHH fused to the yeast invertase signal peptide. pUR4585 is identical to it but encodes C-terminal myc- and his-tag. 26
Fermentations
Bioreactor Bioflo III (New Brunswick Scientific, NJ) was connected via an AFS Biocommand Interface to a personal computer, and data such as pH, temperature, agitation, and DOT (dissolved oxygen tension) were collected in real time. Exhaust ethanol was analyzed with a Xendos 2500 infrared analyzer (both instruments from Servomex, Zoetermeer, The Netherlands). Samples were taken and optical density (OD) at 600 nm was measured to determine the amount of biomass in the vessel. In addition, 1 ml samples from the culture were collected in preweighed Eppendorf tubes and cells were pelleted via centrifugation. Subsequently, cells were washed with 1 ml water and dried at 100°C for at least 24 h. The weight of the dried cells was calculated and used as dry-weight data.
Downstream processing and protein purification
Following fermentation, cultures were harvested and the supernatant separated from the biomass by centrifugation (Sorvall RC-5B plus centrifuge, 5000 rpm, 15 min) and subsequently filtered (0.22 μm, Millipore, Billerica, MA). Sample volume was reduced by tangential flow ultrafiltration (5000 MWCO Vivaflow membrane, Vivascience-Sartorius, Goettingen, Germany). VHH were captured and purified from the supernatant with an AKTA Xpress chromatographic system (GE Healthcare, UK). His-tagged proteins were affinity purified with a His-Trap 1 ml column (GE Healthcare, UK). Untagged proteins were purified by affinity chromatography using protein A (MabSelect SuRe, 5 ml column). Binding and elution buffers, specific for each column type, were the same as indicated by the manufacturer. For desalting, a 2× HiTrap 5 ml or a 1× HiPrep 26/10 desalting column (GE Healthcare, UK) was used. The proteins were eluted in phosphate-buffered saline (PBS) pH 7.0 and stored at −20°C.
Enzyme-linked immunosorbent assay
Enzyme-linked immunosorbent assays (ELISA) were used to test the activity of VHH following formulations, stress treatment, and in vitro release tests. MaxiSorp 96-well plates (Nunc, Roskilde, Denmark) were coated overnight at 4°C with 50 μl rgp140 ZM96, rgp140 UG37, or gp120 IIIB (4 mg/liter). Plates were blocked (200 μl 4% skimmed milk in PBS) and washed three times with PBST. VHH in 50 μl of milk 1%/PBS were added in 4-fold serial dilutions ranging from 40 to 0.01 mg/liter and incubated 1 h at room temperature. After washing with PBST, detection was carried out with 50 μl antillama heavy-chain rabbit serum 1:2000 for 1 h at room temperature and washed three times in PBS. Subsequently, plates were incubated with 50 μl peroxidase-conjugated goat antirabbit monoclonal antibody 1:5000 for 1 h at room temperature. Peroxidase activity was developed by adding 50 μl o-phenylenediamine in the presence of H2O2. The reaction was stopped after 30 min with 25 μl H2SO4 1 M. Absorptions at 490 nm were measured using a microtiter plate reader.
Preparation and characterization of VHH A12 gel formulations
An aqueous solution composed of sorbic acid (0.1% w/w), citric acid monohydrate (1.05% w/w), and sodium citrate dihydrate (1.48% w/w) was prepared and adjusted to pH 4.5. Lyophilized VHH A12 powder (0.1–1% w/w) was added to the solution followed by the gelling agent hydroxyethyl cellulose (HEC, Natrosol 250 HHX, 1.5% w/w). Clear homogeneous gels were formed upon mixing. In addition to these citrate-buffered gels, unbuffered gels were prepared by replacing citric acid and sodium citrate in the formulation with NaCl (0.85%) to maintain osmolality. The gels were characterized with respect to pH, osmolality (The Advanced Osmometer 3250, Advanced Instruments, Norwood, MS), and viscosity (37°C; SV-10 vibrational viscometer, A&D Company, Tokyo, Japan). VHH precipitation in gel formulations (0, 1, 5, or 10 mg/g VHH) was investigated by optical density measurement (OD600, Tecan Infinite M200 microplate reader, Tecan Benelux, Mechelen, Belgium) 60 min after dilution of the gels (50 μl) with vaginal fluid simulant (VFS, 20 μl) and semen simulant (SS, 70 μl). These volumes correspond to the application of 2.5 ml of gel (a typical dose volume in man) in 1 ml of vaginal fluid and 3.5 ml of seminal fluid. 27,28
Manufacture and characterization of vaginal rings containing VHH A12
Silicone elastomer (MED-6382, Nusil Technology, Carpinteria, US) vaginal ring devices, having 54 mm in overall diameter, 7.6 mm in cross-sectional diameter, and comprising three open-ended 3.0-mm-diameter channels for insertion of solid dosage forms (tablets) of appropriate dimensions, were prepared by reaction injection molding (80°C, 2 min) using a laboratory-scale ring-making machine fitted with stainless-steel ring injection molds. Solid dosage inserts containing VHH A12 were prepared by two different methods: direct compression of a powdered mixture of VHH and hydroxypropylmethylcellulose (HPMC) and lyophilization of an aqueous HPMC gel containing VHH A12. For the directly compressed tablet inserts, 50.0 mg of lyophilized VHH A12 was mixed with 950.0 mg of HPMC (100,000 cps, Sigma Aldrich, Dorset, England) in a Speed Mixer DAC 15FVZ-K (Synergy Devices Ltd.) for 1 min at 3500 rpm. Approximately 75.0 mg quantities of this active mix were compressed using a single punch tablet press (Riva Minipress) fitted with tablet die (3.2 mm deep, 3.0 mm wide, and 7.6 mm long). A single insert was then located within each of two channels within the vaginal ring device. For the lyophilized inserts, 20.0 mg of lyophilized VHH A12 and 180.0 mg of HPMC were added to 1.8 ml of HPLC grade water and mixed at 3500 rpm for 1 min in the SpeedMixer before allowing hydrating overnight at 4°C to form a clear homogeneous gel. The gel was then injected into silicone elastomer tubing (SFM3-3650, SF Medical, 3.3 mm internal diameter, 6.35 mm outer diameter). The tubing was cut into 2.5 cm lengths and subsequently lyophilized (the protocol is described in Table 1). The lyophilized rods were removed from the silicone tubing, cut to length, and inserted into the channels of the vaginal ring device. Four such rings were prepared, each containing two VHH-loaded lyophilized inserts.
Freeze Drying Protocol
Individual inserts (n=4) and vaginal rings with inserts (n=4 for each formulation) were individually placed into sealed flasks containing 10.0 ml and 30.0 ml of HPLC grade water, respectively. Each flask was placed in a temperature-controlled orbital shaking incubator (Unitron Infors, 37°C, 60 rpm, throw diameter 2.5 cm). The release medium was sampled daily for 7 days with complete replacement of the release medium. Quantitation of VHH release was performed by SEC-HPLC with UV detection (PerkinElmer series 200 HPLC system) and employing a 50 mM ammonium acetate mobile phase adjusted to pH 5.5.
Permeability of VHH through human vaginal tissue
Healthy human vaginal tissue was obtained from five postmenopausal patients, ages 49–70 years (mean age 59±9 years SD) following vaginal hysterectomies at the Louis Leipoldt Hospital, Bellville, South Africa.
No specimens were obtained where there was clinical evidence of any disease that might have influenced the permeability characteristics of the different specimens. The Ethics Committee of Stellenbosch University and the Tygerberg Academic Hospital approved the study. Specimens were treated and stored as previously described. 29,30 VHH labeled with fluorescein isothiocyanate (FITC) were prepared according to the vendor's instructions (Molecular Probes, Inc., Eugene, OR). The tissue specimens were carefully cut into sections (4 mm diameter) and mounted in flow-through diffusion cells (exposed areas 0.039 cm2) with the epithelial surfaces facing upward. The tissue disks were then equilibrated for 10 min with PBS (pH 7.4) at 37°C in both the donor and acceptor compartments of the diffusion cells. The PBS was removed from the donor compartment and replaced with 1.0 ml of 0.5 mM solution of FITC-labeled VHH in PBS. PBS at 37°C was pumped through the acceptor cell at a rate of 1.5 ml/h and collected by means of a fraction collector at 2 h intervals for 24 h. FITC-labeled VHH was detected using a fluorometric method (520 nm emission and 497 nm excitation, Perkin-Elmer spectrophotometer, Perkin-Elmer, MA).
Permeation studies were performed on seven tissue replicates for each patient. Additional permeation studies were performed using deepithelialized tissue specimens; deepithelialization was achieved by submerging the thawed vaginal tissue in water at 80°C for 30 s, followed by removal of the epithelial layer with tweezers. For all experiments, flux (J) values of FITC-labeled VHH across the vaginal membrane were calculated by means of the relationship J=Q/A·t, where Q=the quantity of compound crossing the membrane (pmol), A=the membrane area exposed (cm2), and t=the time of exposure (min). Steady-state permeation was assumed for a particular vaginal mucosal specimen and VHH when no statistically significant differences (p<0.05, ANOVA and Duncan's multiple range test) between flux values were obtained over at least two consecutive time intervals.
Results
Production of VHH by fermentation
After selection from phage libraries and characterization of specific activity, genes encoding for neutralizing VHH 22,24 were cloned into vectors pUR4547 and pUR4585 and subsequently transformed into S. cerevisiae. Transformant cells were screened and the colonies with the highest secretion level were selected and used for fermentive production (data not shown). Feed rate was set to a fixed value: 1 liter feed per 12 h. Taking into account the constant increase in volume due to the addition of medium, the dilution rate at t=0 h was 0.055 h−1, at t=6 h was 0.041 h−1, and at t=12 h was 0.033 h−1. This was close to the optimal dilution rate of 0.06 h−1 described previously for VHH production. 21 After 12 h of feeding, 1 liter of fermentation fluid was harvested from the vessel in sterile conditions; then a second round of feed was started when the culture parameters stabilized. Three rounds of feed–harvest were performed for each fermentation run.
Two types of fermentations were carried out: one based on glucose as the carbon source and the other one based on ethanol. We previously recorded a lower yield of purified protein in the fed-batch process based on glucose as the sole carbon source compared to the ethanol-based fermentation (58.9 mg/liter vs. 85.6 mg/liter). Therefore, ethanol was selected as the preferred carbon source for the feed phase, whereas the batch phase employed glucose. In ethanol-fed fermentations we obtained biomass yields up to 71±11 g biomass (dry weight) per liter of culture. The yield of purified product obtained ranged between 90.7 and 109.6 mg/liter of fermentation fluids.
Downstream processing and purification
At the end of each run, the recovery of secreted VHH from approximately 4.5 liters of fermentation fluids included four steps. Cells were pelleted by centrifugation, the supernatant was filtered (0.22-μm pores membrane), and an ultrafiltration device (2× Vivaflow 200 membranes, MWCO 5 kDa) was used to reduce the volume to a few hundred milliliters. The processed supernatant was then loaded on a His-Trap column (GE Healthcare, UK) several times. High purity was obtained with all VHH tested with essentially no loss in the unbound fraction.
As an alternative to tagged proteins, VHH lacking tags were produced and purified using a protein A column (also from GE Healthcare). Protein A purification proved efficient and highly selective for these VHH. Protein purity in the eluted fraction was >95%.
Formulations
Gel formulation
A semisolid gel formulation for vaginal administration of VHH A12 was developed and characterized. Preliminary experiments in which various vehicles were tested demonstrated increased solubility of A12 (pI=7.96) at acidic pH and relatively high ionic strengths. Based on these observations, a citrate-buffered vehicle (pH 4.5) with an ionic strength of 278 mM was selected for formulation of VHH A12 up to concentrations of 10 mg/g. Sorbic acid and the nonionic polymer HEC were included as preservative and viscosity enhancer, respectively. The basic characteristics of the gels were assessed in three independent batches: pH 4.6±0.1, osmolality 253±3 mOsm/kg, and viscosity 1.34±0.09 Pa·s. The binding activities of formulated (1 mg/g) and native VHH A12 were similar: the activity ratio formulated/native amounted to 0.97±0.22 and it was consistent for the three clades used in ELISA. Upon storage of the gels for 2 months at 4°C, the ratio amounted to 0.93±0.11; storage at 40°C, however, resulted in a decrease of activity to 0.56±0.05 compared to freshly prepared gels (n=3).
In clinical use, gels will ultimately be diluted by vaginal fluid and, upon coitus, semen. Due to changes in the chemical environment, this dilution may alter the availability of VHH. To simulate this phenomenon, the gels (both buffered and unbuffered, 0–10 mg/g) were diluted with biorelevant volumes of vaginal fluid simulant (VFS) and semen simulant (SS). After 60 min, the optical density at 600 nm (OD600) was determined as a measure of possible precipitation of the VHH. Upon dilution with VFS alone, no precipitation was observed (data not shown). Similarly, dilution with both VFS and SS did not cause precipitation in gel formulations containing A12 at concentrations up to 5 mg/g. The high-dose gels (10 mg A12/g), however, showed an increased OD600, suggesting precipitation of VHH A12 (Fig. 1) in the presence of VFS and SS. The increase in OD600 was about 4-fold higher for the unbuffered high-dose gel compared to the buffered one. The observed precipitation may be related to decreased solubility of VHH A12 in the relatively high pH of SS: in fact, upon dilution with SS, pH increased from 4.6 to 7.7 in the unbuffered gels, but only from 4.6 to 5.8 in the buffered gels.
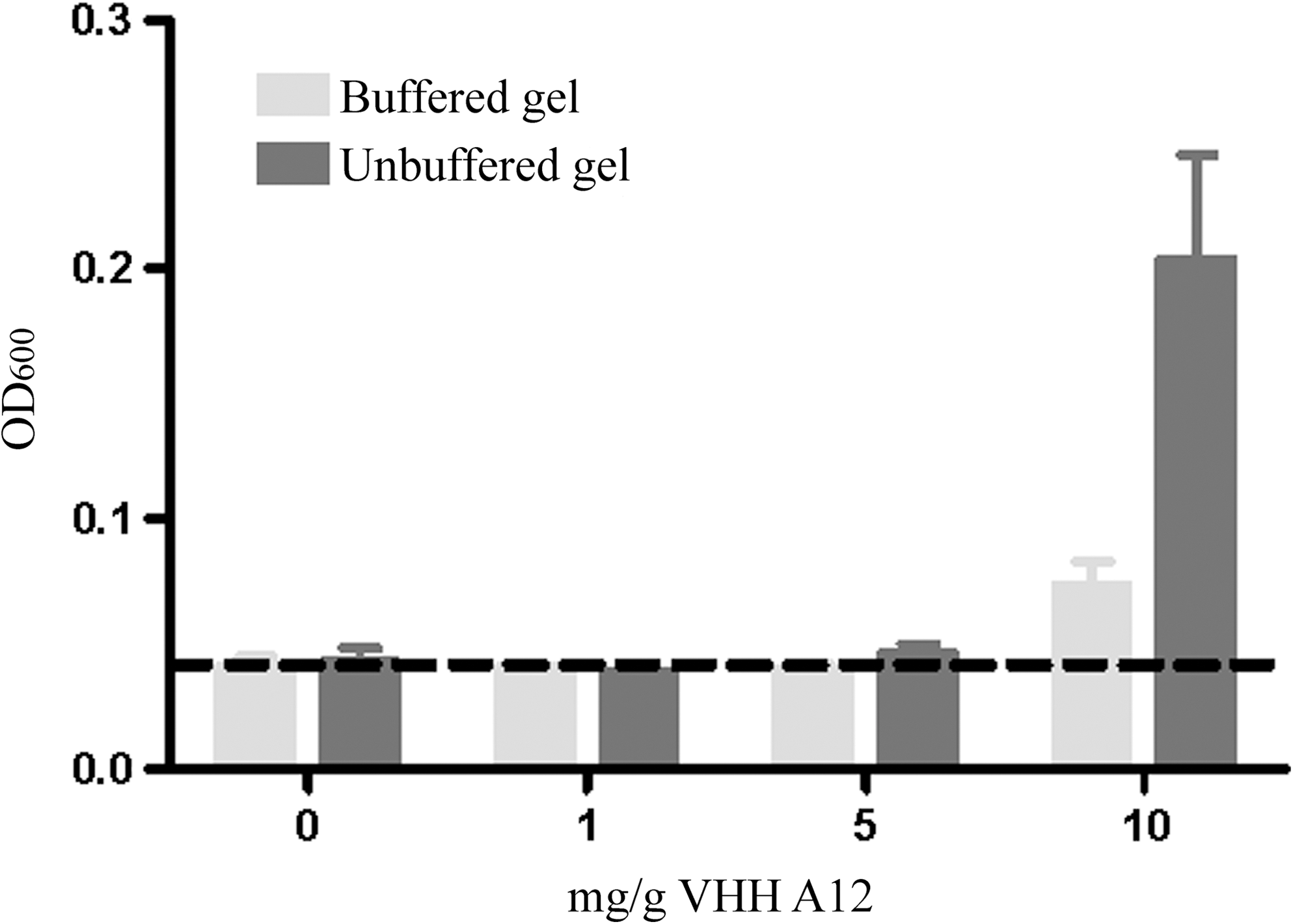
Increase in optical density (600 nm) upon dilution of gels with biorelevant media as a function of VHH A12 concentration. Buffered and unbuffered gels (50 μl) were diluted with 20 μl vaginal fluid simulant (VFS) and 70 μl semen simulant (SS). OD600 was measured after 60 min. The dashed line indicates the background signal (measured in the absence of A12). Data represent mean±SD, n=3.
Vaginal ring formulations
Sustained release of VHH A12 was achieved over 7 days for both directly compressed and lyophilized tablets inserted within a vaginal ring device. For the system having two lyophilized tablets inserted into the ring body, the total quantity of VHH released over 24 h and 7 days was 289 and 1276 μg, respectively, equivalent to 17% and 76% of the initial VHH loading within the ring. By comparison, a single lyophilized tablet placed directly into the release medium (i.e., not inserted into a ring body) provided in vitro release of 626 μg (75%) over 24 h. These results demonstrate that reduced surface area exposure of the tablet insert within the ring significantly reduces the daily release rate and extends the duration of release of the VHH.
The mean cumulative release versus time profile observed for the ring devices comprising two directly compressed tablet inserts (5% w/w VHH loading per tablet) was very similar to that observed for the lyophilized tablet insert device except that the amount of VHH released was significantly greater (5.2 mg over 7 days, equivalent to 67% of the initial VHH loading in the ring), a consequence of the higher absolute VHH loadings in these tablet inserts (Fig. 2). A single directly compressed VHH tablet placed in the release medium achieved 54% release over 24 h, although, unlike the lyophilized system, the tablet had not completely dissolved within this period. Binding data show that A12 formulated in the compressed tablet, inserted in the silicone holder preserves higher activity than when it is included in lyophilized tablets (99% vs. 92% activity, respectively, compared to A12 control). The activity of VHH A12 released after 7 days in both the compressed and lyophilized tablet insert experiments was higher than 90% of the A12 control on all the clades tested.
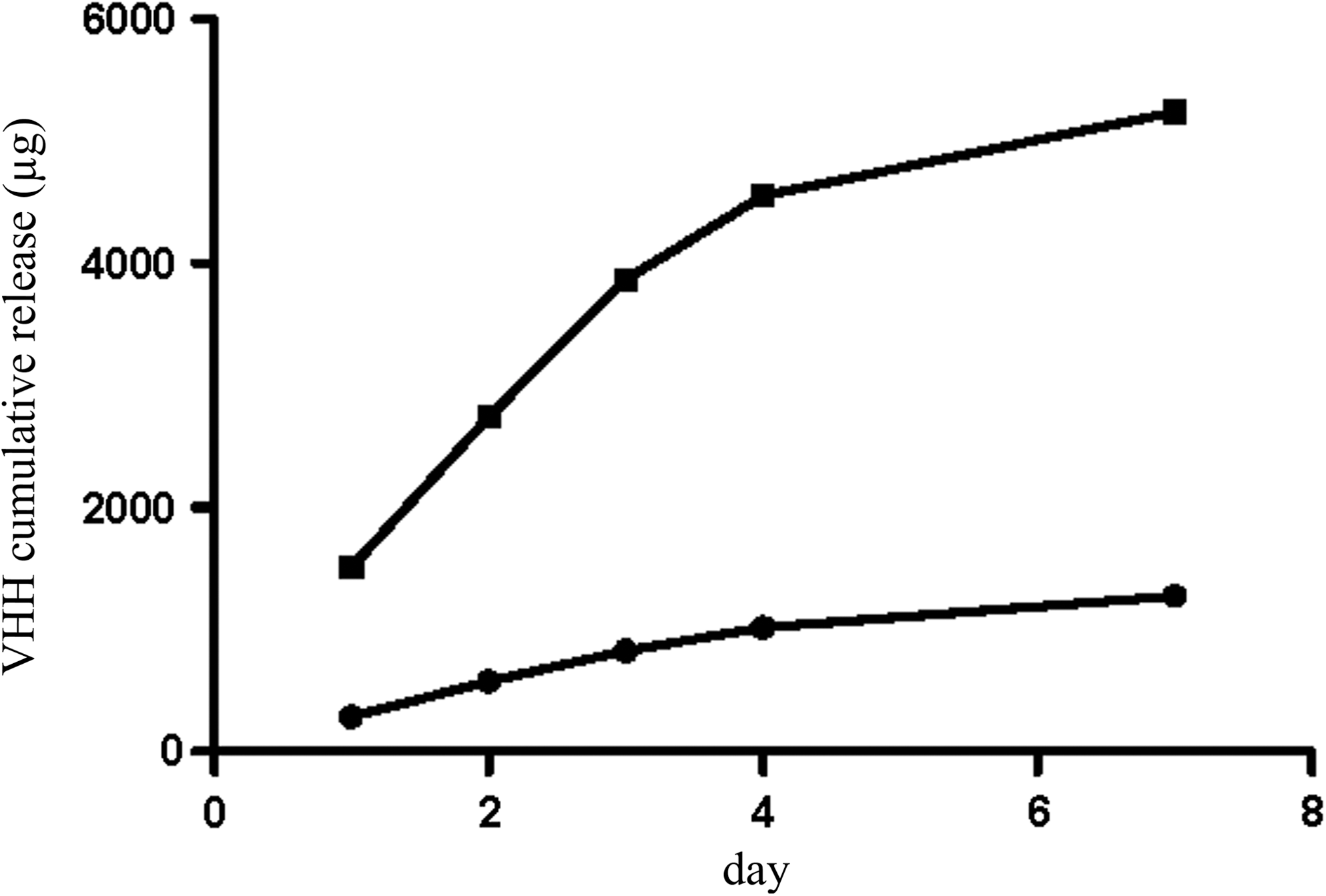
Mean in vitro cumulative release of VHH A12 from a vaginal ring device containing two compressed tablet inserts each loaded with 5% w/w VHH (square) and from a vaginal ring device containing two freeze-dried tablet inserts each loaded with 10% w/w VHH (circle). Cumulative release was measured at days 1, 2, 3, 4, and 7.
Stability
Exposure to high temperatures due to lack of cold storage facilities and acidic pH in the cervicovaginal environment are important factors to be considered in the development of HIV microbicide products. Therefore, the ability of VHH to withstand thermal and pH stress was investigated. Solutions of VHH A12 of 1 mg/ml in PBS preincubated at 60°C, 70°C, or 80°C for 10 min were more than 88% active compared to an untreated control (Fig. 3A). The thermal denaturation of VHH A12 and that of four other recently selected α-HIV VHH were compared and they were very similar, indicating that the remarkable heat tolerance is not unique to VHH A12. 24
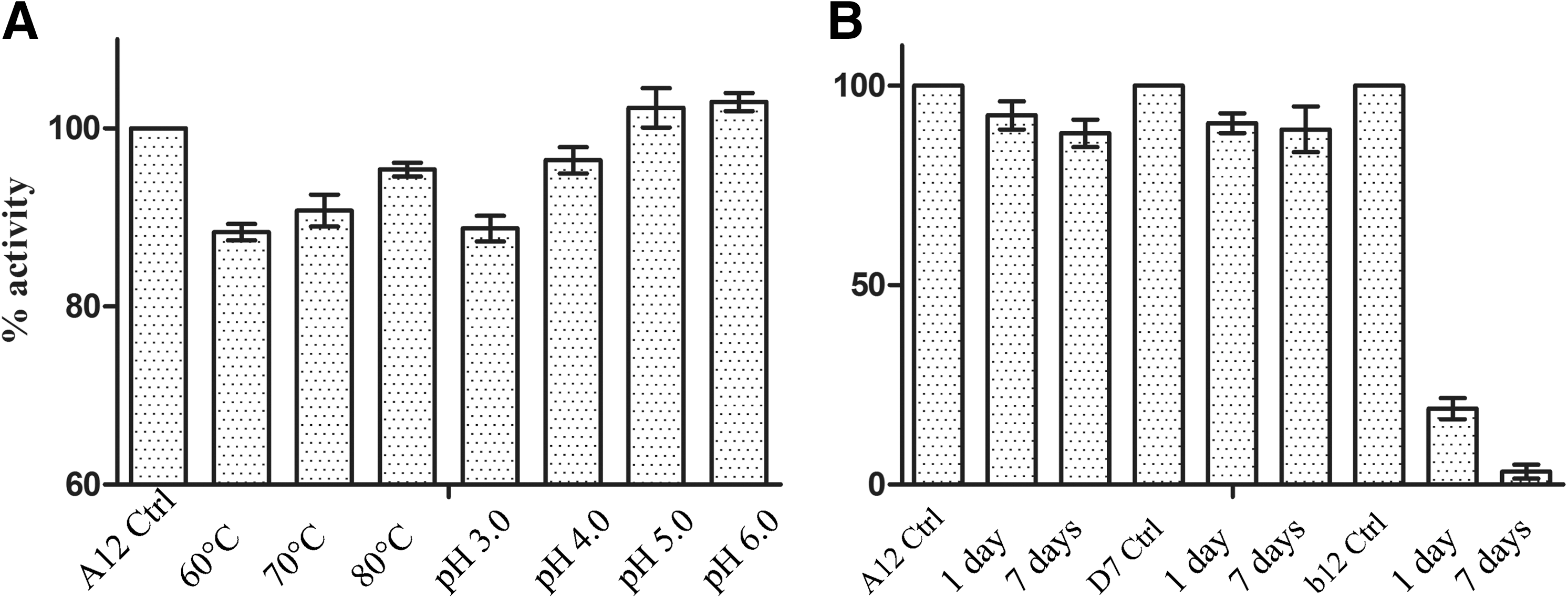
Incubation of VHH A12 solutions in PBS at various pH (3.0, 4.0, 5.0, and 6.0; vaginal pH is<4.5 31 ) for 48 h at 4°C demonstrated at least 87% of the original binding activity (Fig. 3A). In comparison, loss of binding was observed in the conventional antibody b12 starting from 24 h incubation at all tested pH. Moreover, after 1 week the activity of A12 and D7 was substantially unaltered (<10% loss) at any pH, whereas incubation at pH 4.0, 5.0 and even 6.0 was detrimental for b12 (1% of residual activity) and incubation at pH 3.0 caused a complete loss of signal in ELISA (Fig. 3B)
Permeability
Since HIV primary target cells (Langerhans cells and CD4+ T cells) are localized in the vaginal mucosa, 32 it is of critical importance for an entry inhibitor microbicide based on virus binding as well as on cell receptors binding to be present in the tissues where the infection may occur. We investigated the diffusion kinetics of FITC-labeled VHH through human intact and deepithelialized vaginal mucosa. Deepithelialized mucosa mimics the situation in which the integrity of the vaginal epithelium has been compromised, due to ulcerations associated with a variety of pathological conditions or sexual intercourse, even consensual.
The antibody fragment diffused through intact and deepithelialized vaginal mucosa at 37°C reaching steady-state flux values after 4 h of 10.24±1.70 and 9.90±1.74 pmol cm2 min−1, respectively. Contrary to what we expected, the difference in permeability was quite small, and was likely related to the removal of negatively charged epithelium. VHH A12 acquires a net positive charge at the physiological pH conditions under which the permeability experiments were conducted. The cationic antibody fragment will therefore be attracted to negatively charged epithelium, facilitating its partitioning from the aqueous buffer into the barrier domains of the intact mucosal membrane. Deepithelialization, with the consequent loss of negative charge, decreases the attractive forces between the positively charged VHH A12 and the mucosa, thereby lowering its penetration potential into the mucosa. Figure 4 shows the mean estimated steady state (20–24 h) flux values for five peptides/proteins across frozen/thawed human vaginal mucosa vs. molecular weight.
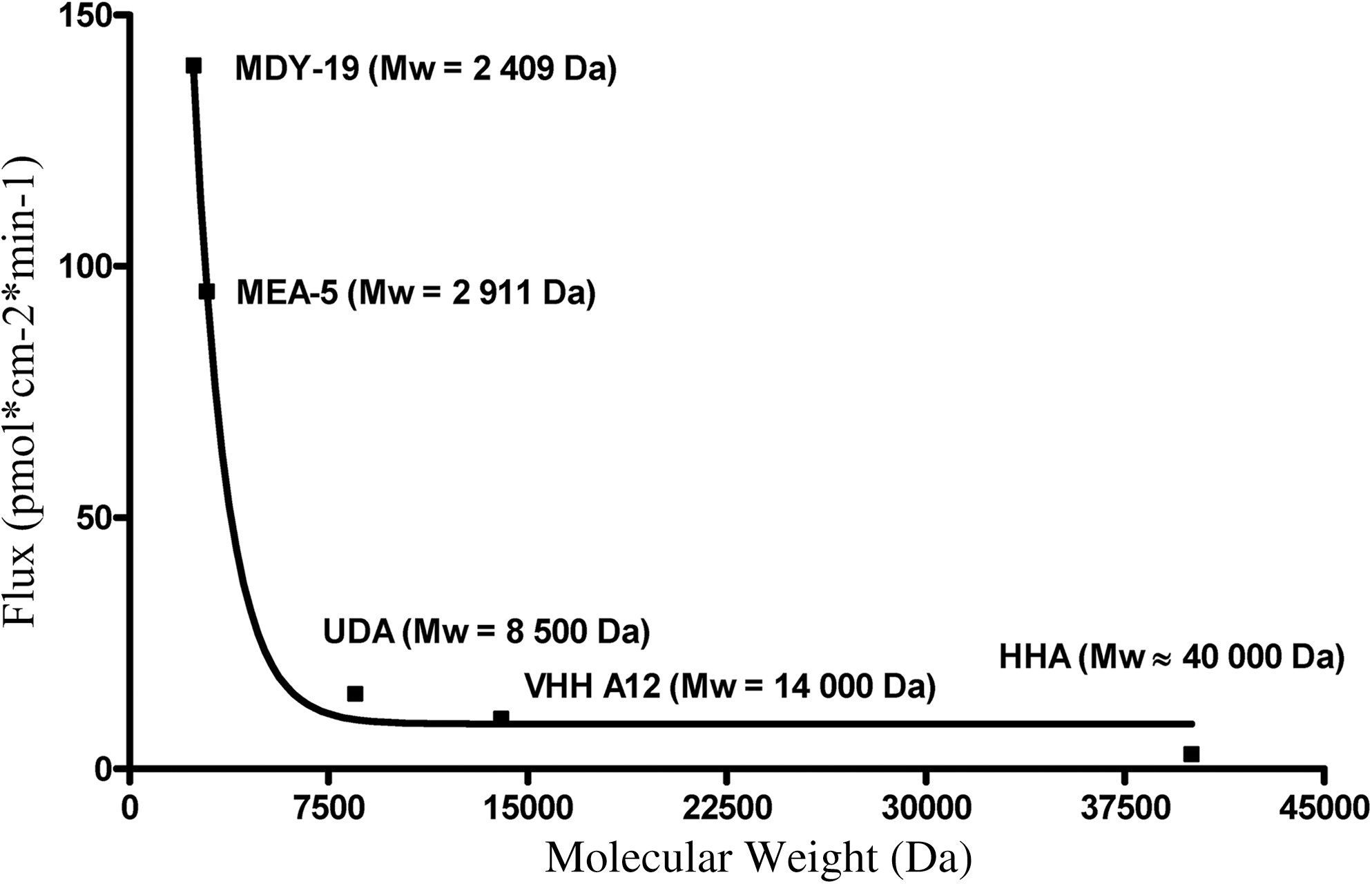
Mean estimated steady state (20–24 h) values for five peptides/proteins across frozen/thawed human vaginal mucosa vs. molecular weight. VHH A12 flux fits in the model curve at its actual MW value.
Discussion
In this study we assessed the feasibility of an HIV-1 microbicide based on llama heavy-chain antibody fragments (VHH) capable of cross-clade neutralization. A few clones including A12 and D7 were previously characterized with a high potency of neutralization over a broad spectrum of HIV-1 isolates 22 and the structure of D7 and its mode of interaction with gp120 were described. 33 Currently our panel of VHH has increased to about 60 clones recognizing several epitopes on both gp41 and gp120 with some clones displaying breadth of neutralization better than the human monoclonal antibody b12. 24
A number of relevant properties for developing VHH as HIV microbicides are independent of molecular affinity of a specific clone, and can be associated more generally with the wider class of llama antibody fragments. An effective HIV microbicide will be impractical if the cost per dose is not affordable. We have developed methods for producing VHH using fed-batch fermentations based on ethanol as the sole carbon source. Fed batch processes are routinely used by the biotech industry for production of biopharmaceuticals in large scale reactors up to 100 m3. 34 Using a bioreactor with a working volume of 2 liters we could affinity purify about 100 mg of VHH per liter of fermentation fluids, and this yield can be increased by at least a factor of 10 in industrial settings, as it was achieved in the production of antirotavirus VHH that is currently being tested in a feeding trial of children in India (L. Hammarstrom, personal communication).
Several formulation approaches are being actively evaluated for vaginal administration of HIV microbicides, including gels, films, and rings. 35 The preferred choice of formulation will ultimately depend upon many factors. A preferred formulation strategy would involve either once-a-day administration of a microbicide product (e.g., through use of a mucoadhesive, site-retentive gel formulation), or sustained release from a single vaginally administered device over a period of many days or weeks. The feasibility of formulating VHH in a gel was evaluated. VHH A12 was formulated in a citrate-buffered gel with pH and osmolality falling within the physiological range of vaginal fluids, at concentrations up to 10 mg/g without losing binding activity. Considering the use of microbicides in tropical climates, the observed drop in binding activity of VHH A12 upon storage of the gel formulation at elevated temperature (40°C) requires further investigation. VHH were also formulated as solid tablets, using two different methods of manufacture (i.e., direct compression and lyophilization), and inserted into a vaginal ring device for prolonged retention within the vaginal cavity. It is important to note that although the in vitro release results presented here specifically relate to a ring comprising two VHH-loaded tablet inserts, there is scope to produce ring devices with a greater number of inserts, resulting in a proportionate increase in the amount of VHH released.
Binding assays performed on VHH after gel incubation and tablet formulation showed that these molecules retain more than 90% of their original activity. Subjecting VHH to temperatures up to 80°C and to acidic conditions (pH 3.0) caused activity to retain 88% and 87%, respectively. More than 75% of original activity was also maintained after incubating VHH for a month at 37°C. Taken together, these data show that VHH are robust molecules able to maintain a high activity level in the harsh conditions met during storage and use as a vaginal microbicide. No other conventional, full-chain immunoglobulin has so far been shown to withstand such conditions.
Studies describing the correlation between in vitro and in vivo activity of the monoclonal antibody b12 have indicated that effective concentrations in vivo are typically up to 103-fold higher than in vitro 36 and the difference is even more accentuated in the case of other microbicide candidates. 37 Assuming a vaginal fluid volume of 1 ml, the application of a single dose of 2.5 g gel containing 10 mg A12/g would result in vaginal concentrations that are about 103 to 2×106-fold higher than the IC50 of VHH A12 measured on several tier 1 clade C viruses. 22 Given an average vaginal fluid production of 6 ml per day, the concentration of VHH A12 released every day over 1 week by compressed tablets inserted in a vaginal ring would be about 2×102 to 3×105-fold higher than the IC50 of A12 in vitro against some viruses. These data are promising and, provided VHH are selected against viruses used in current animal models, suggest that low doses are sufficient for full protection.
VHH A12 readily traversed both intact and damaged vaginal tissue at rates that are similar to those of similarly sized molecules. This suggests that the vaginal epithelium does not work as such a tight barrier for the small antibody fragments. So VHH permeate into the submucosal tissue, where interstitial dendritic cells and Langerhans cells reside. The ability to traverse the vaginal mucosa and enter the systemic circulation offers the possibility of creating a second barrier either by blocking the receptor binding sites on the virus or by blocking the relevant receptors of the immune cells present in the vaginal mucosa. However, the presence of VHH raises the issue of immunogenicity. Even though there is evidence that VHH do not elicit immune response or create adverse effects because of their clearance via the kidneys, further studies need to be performed. 38,39
In conclusion, we suggest that a coitally related application of gel loaded with VHH or the formulation of VHH in IVR has the potential to perform as well as the successful 1% tenofovir gel formulation in terms of drug thermal and proteolytical stability, bioavailability, and cost per dose. A number of VHH were cross-reactive for the in vitro neutralization of SHIV (unpublished results), therefore suggesting that primate models can be readily used to test the effectiveness of these molecules in vivo.
Footnotes
Acknowledgments
This study was supported by EU grants Europrise and EMPRO. The authors thank Dr. Lucy Rutten for critical reading of the manuscript.
Author Disclosure Statement
No competing financial interests exist.