
Abstract
Our aim was to characterize for the first time the genetic diversity of HIV in Cape Verde Islands as well as the drug resistance profiles in treated and untreated patients. Blood specimens were collected from 41 HIV-1 and 14 HIV-2 patients living in Santiago Island. Half of the patients were on antiretroviral treatment (ART). Pol and env gene sequences were obtained using in-house methods. Phylogenetic analysis was used for viral subtyping and the Stanford Algorithm was used for resistance genotyping. For HIV-1, the amplification of pol and env was possible in 27 patients (66%). HIV-1 patients were infected with subtypes G (13, 48%), B (2, 7%), F1 (2, 7%), and CRF02_AG (2, 7%), and complex recombinant forms including a new C/G variant (n=8, 30%). Drug resistance mutations were detected in the PR and RT of three (10%) treated patients. M41L and K103N transmitted drug resistance mutations were found in 2 of 17 (12%) untreated patients. All 14 HIV-2 isolates belonged to group A. The origin of 12 strains was impossible to determine whereas two strains were closely related to the historic ROD strain. In conclusion, in Cape Verde there is a long-standing HIV-2 epidemic rooted in ROD-like strains and a more recent epidemic of unknown origin. The HIV-1 epidemic is caused by multiple subtypes and complex recombinant forms. Drug resistance HIV-1 strains are present at moderate levels in both treated and untreated patients. Close surveillance in these two populations is crucial to prevent further transmission of drug-resistant strains.
Introduction
C
Since the advent of highly active antiretroviral therapy in developed countries we have seen an improved prognosis of HIV-1-infected patients, with a significant decrease in morbidity and mortality rates associated with this infection. 4,5 Combined antiretroviral therapy (cART) has been introduced recently in resource-limited countries. 6 However, there are concerns about the low efficacy of cART in these countries, which could lead to the rapid and uncontrolled emergence and transmission of drug-resistant viruses. 7 Three factors that can lead to low cART efficacy are the limited HIV-RNA monitoring of patients undergoing therapy, the unregulated and unmonitored use of antiretroviral drugs bought on the black market or abroad, and the lack of drug resistance surveillance systems. 7 –10 Despite these limitations, transmitted HIV drug resistance is still residual (<5%) in most sub-Saharan countries. 11 –17
Worldwide strains of HIV-1 are phylogenetically classified into four very divergent groups, M, N, O, and P. 18 Amino acid sequences of Gag and Env can differ between groups up to 30% and 47%, respectively. HIV-1 group M, the only pandemic group, has diversified into nine divergent subtypes (A, B, C, D, F, G, H, K, and J), six subsubtypes (F1, F2, A1–A5), and multiple circulating recombinant forms. 19,20 Amino acid sequences of Env can differ by up to 20% within a particular subtype and over 35% between subtypes. As for HIV-2, which is mostly restricted to a few West African countries including Cape Verde, despite the existence of eight genetic groups named A to H only groups A and B seem to be spreading, with group A much more common than group B. 21 –23
The extraordinary HIV diversity is seen as a major hurdle to the development of a prophylactic vaccine. 24 HIV diversity and its rapid evolution may present significant problems to diagnosis and to antiviral therapy. For example, significant antigenic changes in HIV-1 group O viruses have recently been shown to hinder the diagnostic performance of several serodiagnostic assays. 25 Likewise, many comparative studies have shown that the sensitivity and specificity of viral load assays vary depending on HIV-1 group or subtype. 18,26 –29 Regarding antiviral therapy, there are major differences in the susceptibility of HIV-1 and HIV-2 to the current available drugs. Indeed, HIV-2 is naturally resistant to nonnucleoside reverse transcriptase inhibitors (NNRTIs) and it presents a diminished sensitivity to certain protease inhibitors (PI). 30 Moreover, most cART regimens used in HIV-1 patients are unable to fully suppress HIV-2 replication, to increase the number of CD4 cells, or to prevent the accumulation of drug-resistant mutations. 31 Diversity among HIV-1 isolates also affects antiviral therapy. For instance, like HIV-2, group O strains are resistant to NNRTIs. 32,33 Natural polymorphisms found in the protease and reverse transcriptase of some non-B subtypes may lower their genetic barrier to resistance to some protease and reverse transcriptase inhibitors. 17,34 Finally, certain drug-resistant mutations accumulate more frequently and rapidly in some non-B subtypes than in HIV-1 B (e.g., K65R, L90M, and V106M in subtype C and I54V/L in subtype G). 34 –36 Hence, HIV diversity has major implications for the development of resistance, for the genotypic evaluation of drug resistance, and for the selection of therapeutic regimens, at least in countries in which both HIV-1 and HIV-2 circulate such as Cape Verde.
Cape Verde, like most other resource-limited countries, adopted the World Health Organization (WHO) public health approach in antiretroviral therapy (ART) utilization. 37 cART has been available free of charge since December 2004 and was initially given to a group of 148 patients. The preferential first line regime adopted includes a combination of lamivudine (3TC), stavudine (d4T), and nevirapine (NVP). Presently, there are no data concerning the genetic diversity of HIV in Cape Verde. Moreover, there is no information about the prevalence and nature of primary and secondary drug resistance in this country. The aim of the present work was to determine, for the first time, the HIV-1 and HIV-2 genetic forms present in Cape Verde and the drug resistance profiles in treated and untreated patients.
Materials and Methods
Patients
Blood samples were collected during 2005 (n=37), 2006 (n=14), and 2007 (n=4) from 41 HIV-1- and 14 HIV-2-infected patients living in Santiago Island and attending the Delegacia de Saúde, Cidade da Praia, Cape Verde. Nineteen of the HIV-1 individuals had received cART, 18 were drug naïve, and for four there were no indication on the drug regimen. Half of the HIV-2 patients were on cART. The epidemiological characteristics of the HIV-1 and HIV-2 patients are described in Table 1. Plasma samples were preserved at −20°C in Cidade da Praia and thereafter at −80°C in Lisbon, Portugal. Serologic diagnosis of HIV infection and CD4+ T cell counts were done in Cape Verde; plasma viral load was determined with the Cobas Amplicor HIV-1 Monitor test version 1.5 (Roche) in Lisbon, Portugal. Written informed consent was obtained from all participants and the study was approved by the ethics committees of the participating institutions.
Fisher exact test.
Fisher exact test comparing the number of patients below or above 40 years old.
Fisher exact test comparing sexual transmission with other routes of transmission.
Mann–Whitney test.
Fisher exact test comparing the number of patients on ART with untreated patients.
ART, antiretroviral therapy; nd, not done.
Viral RNA extraction, PCR amplification, and sequencing
HIV-1 viral RNA was extracted from 140 μl of plasma using the QIAamp Viral RNA miniKit (QIAGEN). Reverse transcriptase polymerase chain reaction was performed with the TitanOne Tube reverse transcriptase polymerase chain reaction System (Roche). Nested polymerase chain reaction was done to obtain a 409-bp fragment from the C2V3C3 env region, a 532-bp fragment containing the part of the gag gene and all the PR region, and a 1026-bp fragment from the RT region. Thermal cycling conditions and primers sequence and position were described elsewhere.
38,39
HIV-2 viral RNA was extracted from plasma as described above. Nested polymerase chain reaction was done to obtain a 373
Phylogenetic and recombination analysis
Sequences were aligned with sequences from reference strains collected from the Los Alamos Sequence Database (
Resistance mutation analysis
Genotypic analysis of HIV-1 drug resistance was performed using the Stanford Genotypic Resistance Interpretation Algorithm (
Statistical analysis
The Mann–Whitney test was used to compare groups. The frequencies of drug resistance mutations of viruses from Cape Verde were compared with those of all subtype B and G sequences available at the Stanford HIV Drug Resistance Database using the Fisher exact test. p values <0.05 were considered significant.
Results
We analyzed samples from 55 patients, 41 (75%) infected with HIV-1 and 14 (25%) infected with HIV-2. There were no significant epidemiological or immunological differences between HIV-1- and HIV-2-infected patients (Table 1). Most patients (60%) were female and were aged 40–49 years. The infection was mostly transmitted by the sexual route. Most patients had less than 500 CD4+ T cells, which is suggestive of long-term infection. As expected, viral load was significantly lower in treated HIV-1 patients (N=18) than in untreated patients (N=19) (median HIV RNA copies per ml of plasma; range, 400; 400–93,000 vs. 43,900; 400–513,000; p<0.0001).
For HIV-1, the amplification of the PR, RT, and/or C2V3C3 genomic regions was possible in 27 HIV-1 samples (66%). Ten of these patients (37%) were on cART and 17 (63%) were drug-naive individuals. PR and RT sequences were obtained in 26 (63%) and 27 (66%) patients, respectively. C2V3C3 env gene sequences were obtained in 24 (59%) patients. All sequences were checked using the program Hypermut and none showed evidence of hypermutation.
Phylogenetic analyses revealed that 13 (48%) viral isolates were subtype G, six (22%) were non-G [two (7%) subtype B, two (7%) subtype F1, and two (7%) CRF02_AG], four (15%) were recombinants between subtype G and other subtypes or U strains [1 G (PR)/G (RT)/H (C2V3C3); 1 G/G/B; 1 C/C/G; 1 U/G/-], two (7%) were recombinants between CRF02_AG and subtype B and/or U strains [1 U/B/CRF02_AG; 1 U/CRF02_AG/CRF02_AG], and two (7%) were recombinants between the A subtype and U strains (U/A/U) (Fig. 1).
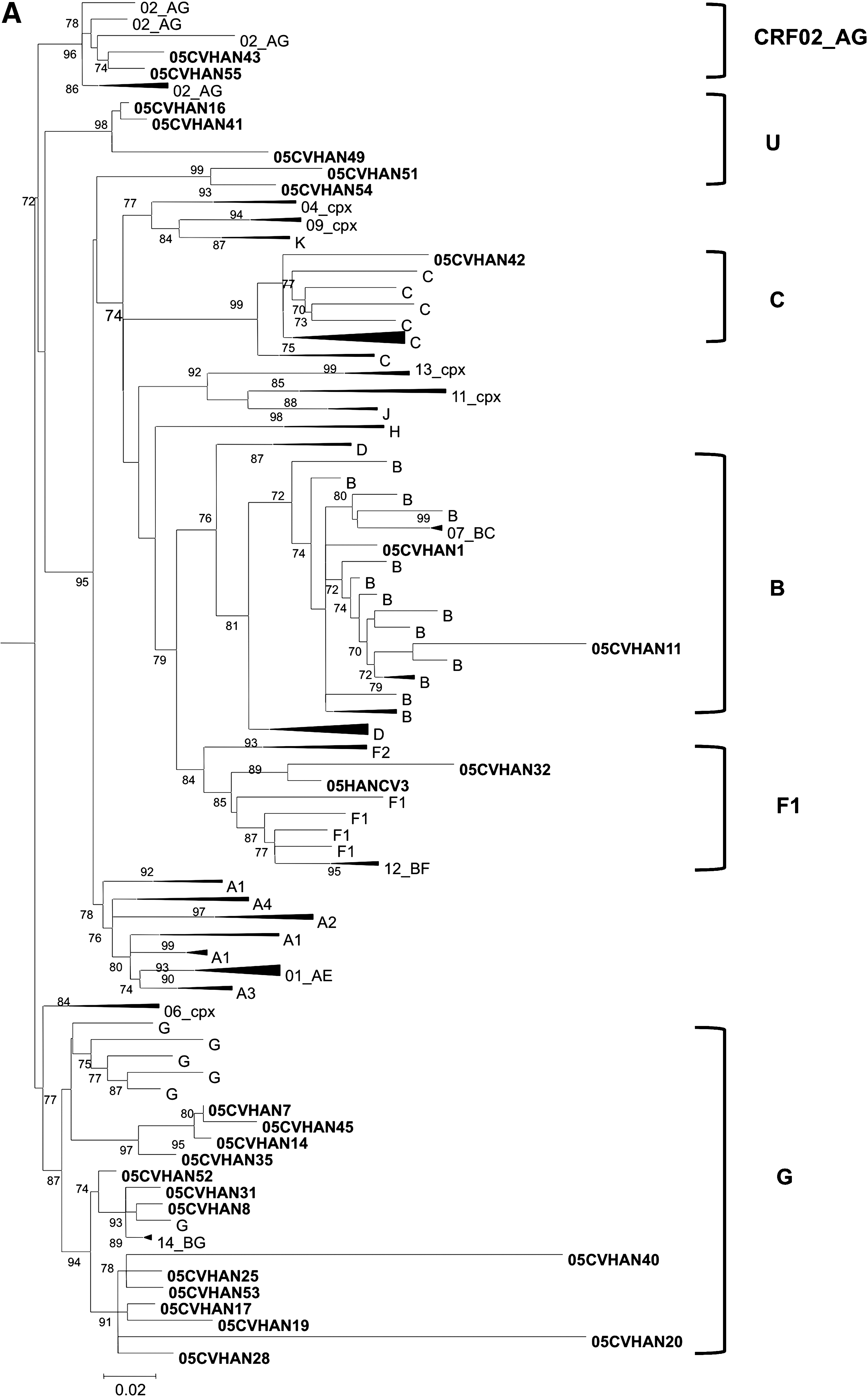
Genetic subtypes and evolutionary relationships of the HIV-1 viruses sequenced in this study based on maximum likelihood phylogenetic trees of the PR
There were no major mutations associated with PI resistance in the naive population. The minor mutations associated with PI resistance, L10I (6%) and A71V (6%), were detected in two unrelated drug-naive patients (Table 2). The M41L, K103N, and Y318F mutations conferring resistance to NRTIs or NNRTIs were found in 3 of 17 untreated patients. Only M41L and K103N are considered transmitted drug resistance mutations. 48 –50 Hence, our results reveal a 12% (2 out of 17 patients) rate of transmitted drug resistance in Cape Verde.
PR, protease; RT, reverse transcriptase; PI, protease inhibitor; NRTI, nucleoside reverse transcriptase inhibitor; NNRTI, nonnucleoside reverse transcriptase inhibitor. S, susceptible; I, intermediate resistance; R, high-level resistance; LLR, low-level resistance; PLLR, potential low-level resistance; 3TC, lamivudine; ABC, abacavir; AZT, zidovudine; d4T, stavudine; ddI, didanosine; FTC, emtricitabine; TDF, tenofovir; EFV, efavirenz; NVP, nevirapine.
Resistance mutations were detected in 3 of 10 (30%) patients undergoing cART (Table 3). No major mutations associated with PI resistance were found in this population. The minor mutations L10I/V and V11I, associated with PI resistance, were found in two patients infected with complex recombinant isolates. The K65R, T69d, K103N, and Y181C mutations, which confer resistance to the NRTIs and NNRTIs, were all detected in patient 05HANCV55, who was infected with a CRF02_AG strain.
U, untypable; S, susceptible; I, intermediate resistance; R, high-level resistance; LLR, low-level resistance; 3TC, lamivudine; ABC, abacavir; AZT, zidovudine; d4T, stavudine; ddI, didanosine; FTC, emtricitabine; TDF, tenofovir; EFV, efavirenz; NVP, nevirapine.
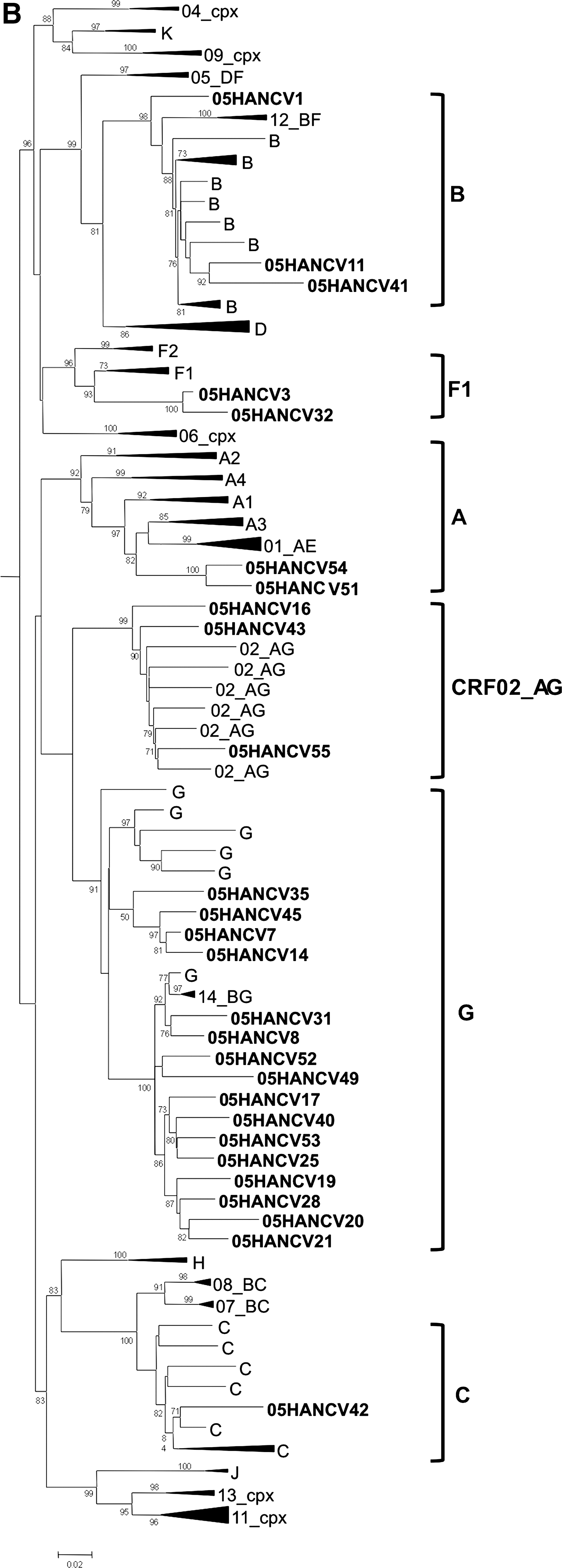
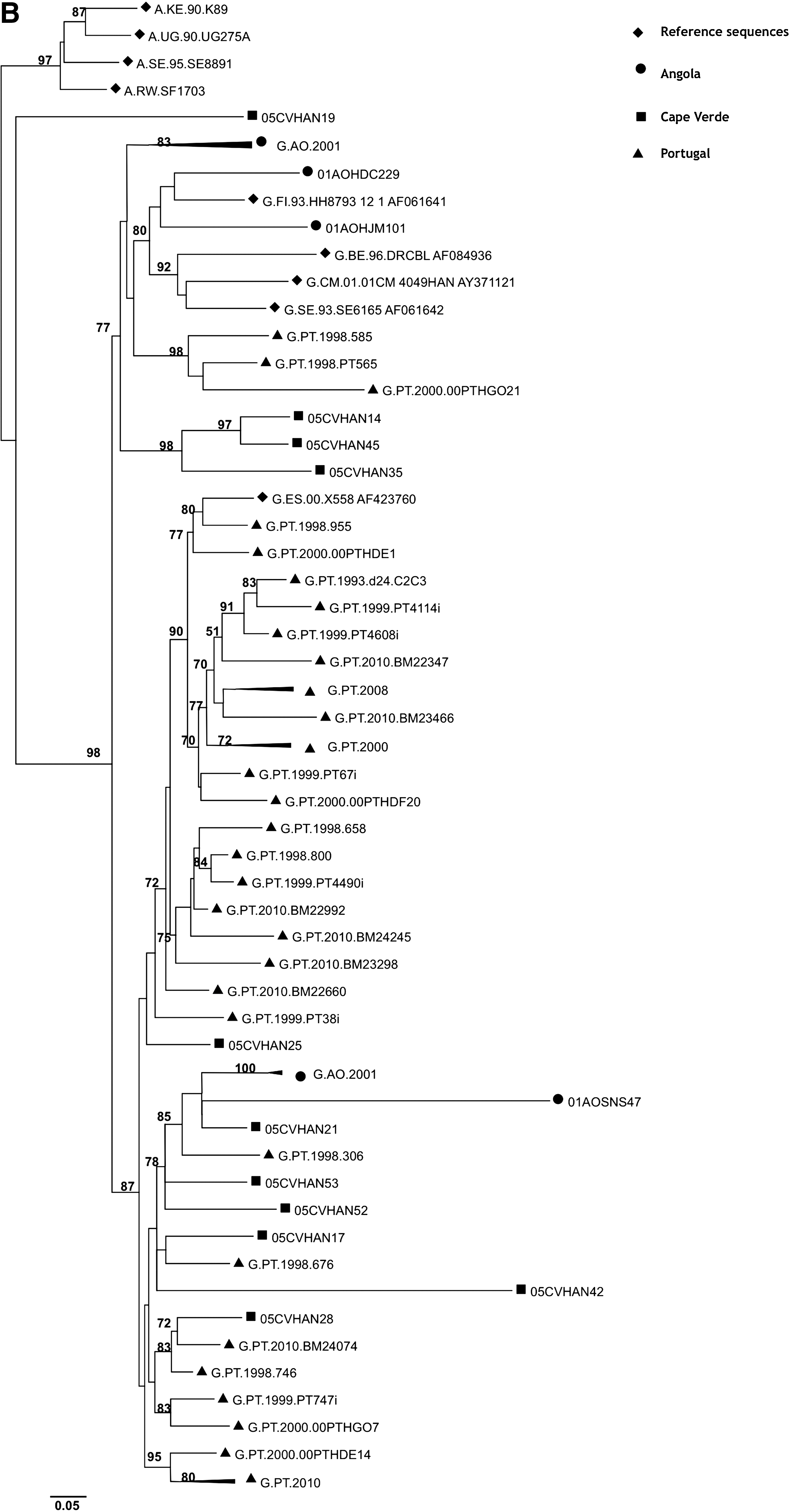
The frequency of some previously known PR polymorphisms detected in subtype G isolates (29% of polymorphisms) was significantly different (p<0.05) when compared to isolates of similar subtype found throughout the world (Fig. 2). Likewise, the frequency of the RT polymorphisms found in the G subtype (38% of polymorphisms) was significantly different (p<0.05) when compared to isolates from worldwide treatment-naive patients infected with the same subtype (Fig. 3). These results are a clear indication that the G strains circulating in Cape Verde are highly divergent and genetically complex. The origin of the G strains from Cape Verde was investigated by phylogenetic analysis using RT and C2V3C3 env sequences from reference G sequences collected worldwide as well as sequences from Angola and Portugal deposited in the Los Alamos database. We found that the majority of the sequences from Cape Verde were closely related to sequences from Portugal and/or Angola (100% in the RT and 91% in env) (Fig. 4). These results indicate that the HIV-1 variants circulating in Cape Verde have their origin in one of these two countries.
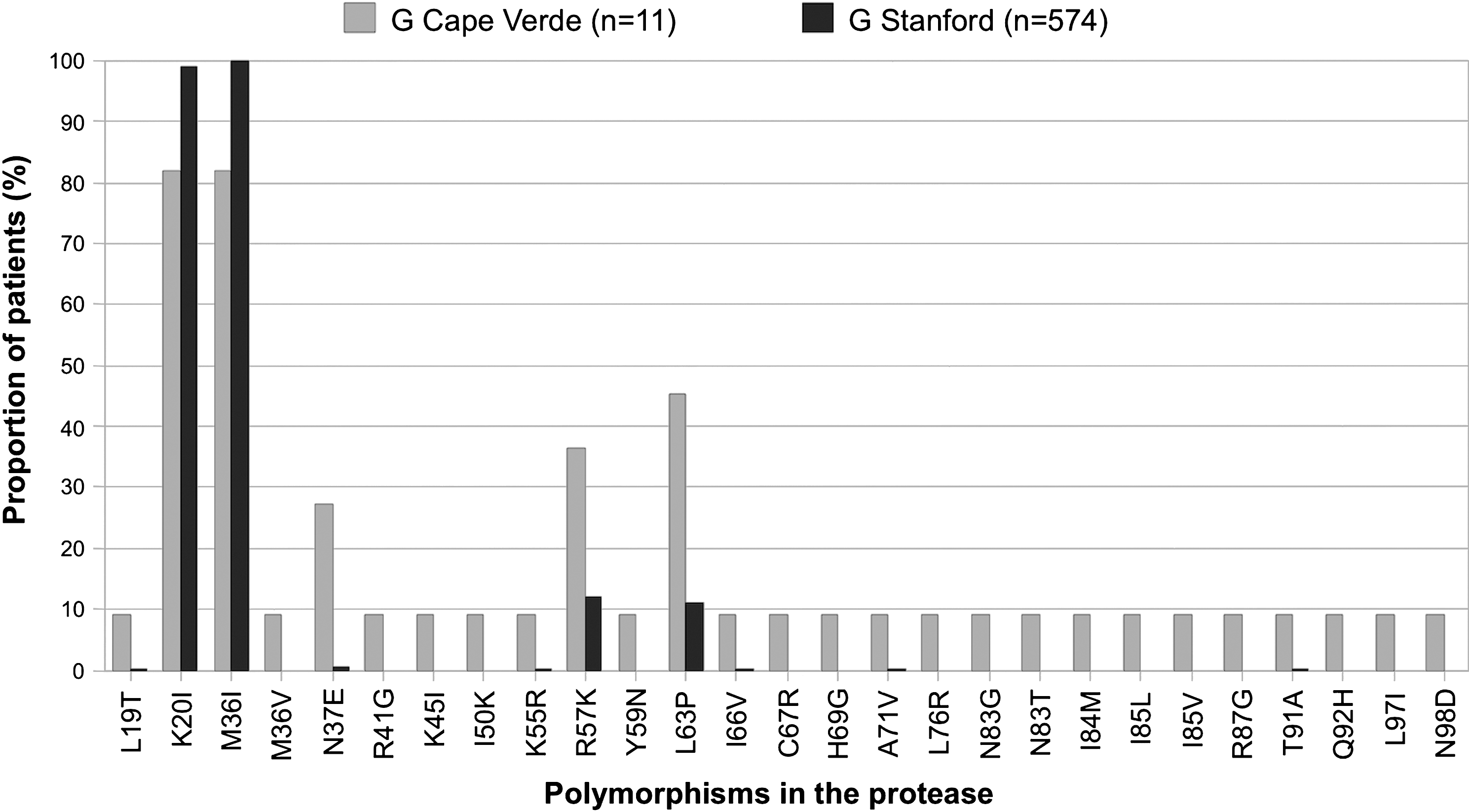
Minor mutations and natural polymorphisms detected in the protease of drug-naive patients from Cape Verde. Only the mutations whose frequency is significantly different (p<0.05, Fisher's exact test) than that found in the Stanford database for untreated patients are shown.
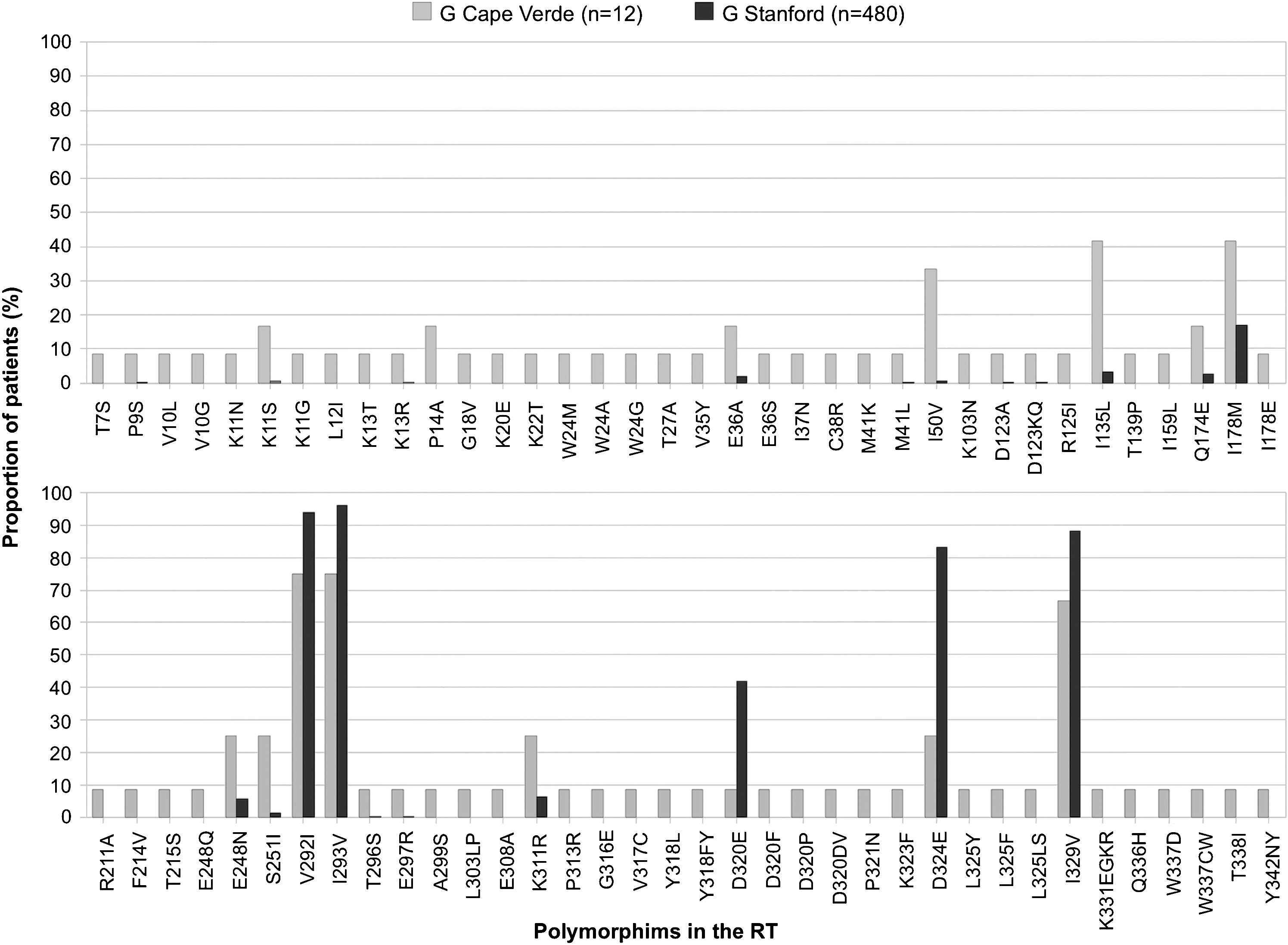
Resistance mutations and natural polymorphisms detected in the RT of drug-naive patients from Cape Verde. Only the mutations whose frequency is significantly different (p<0.05, Fisher's exact test) than that found in the Stanford database for untreated patients are shown.
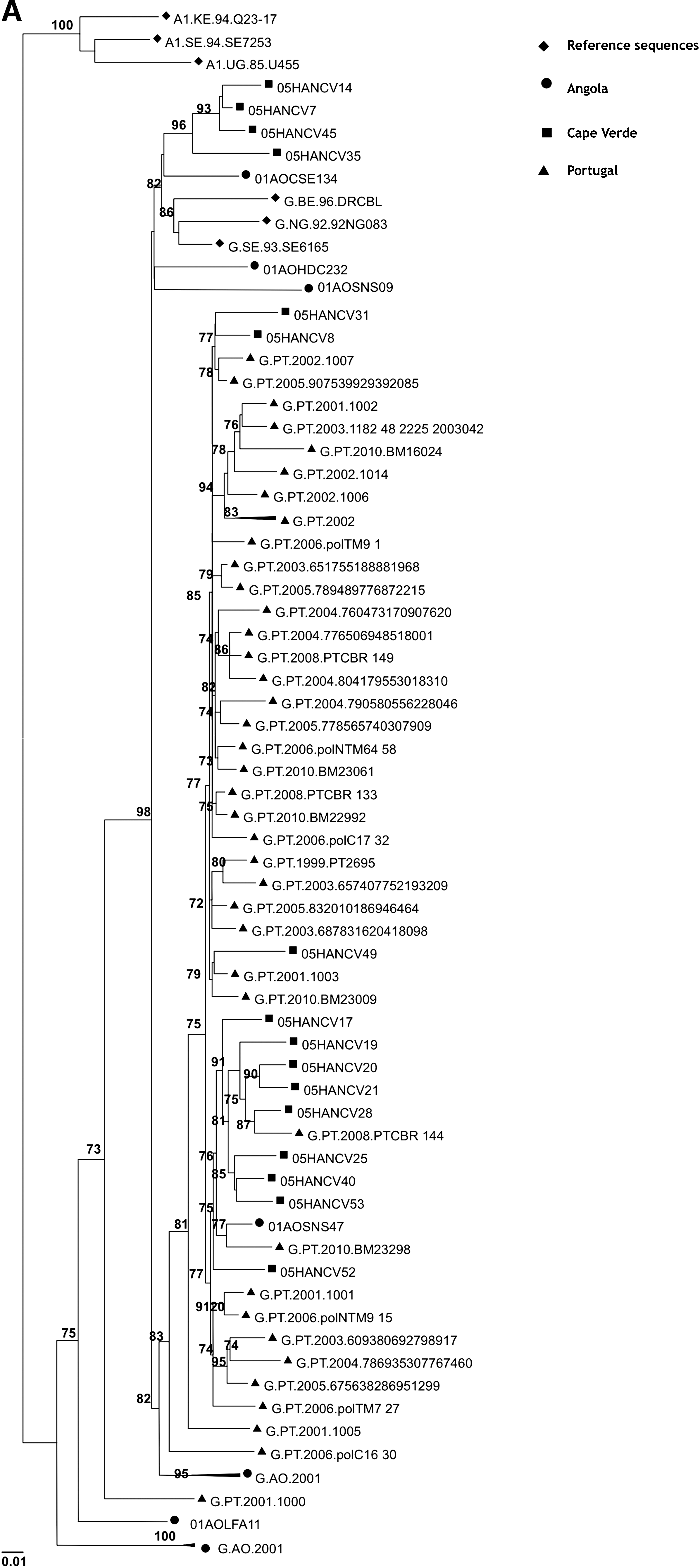
Origin of the HIV-1 subtype G isolates from Cape Verde. Phylogenetic trees were built using RT
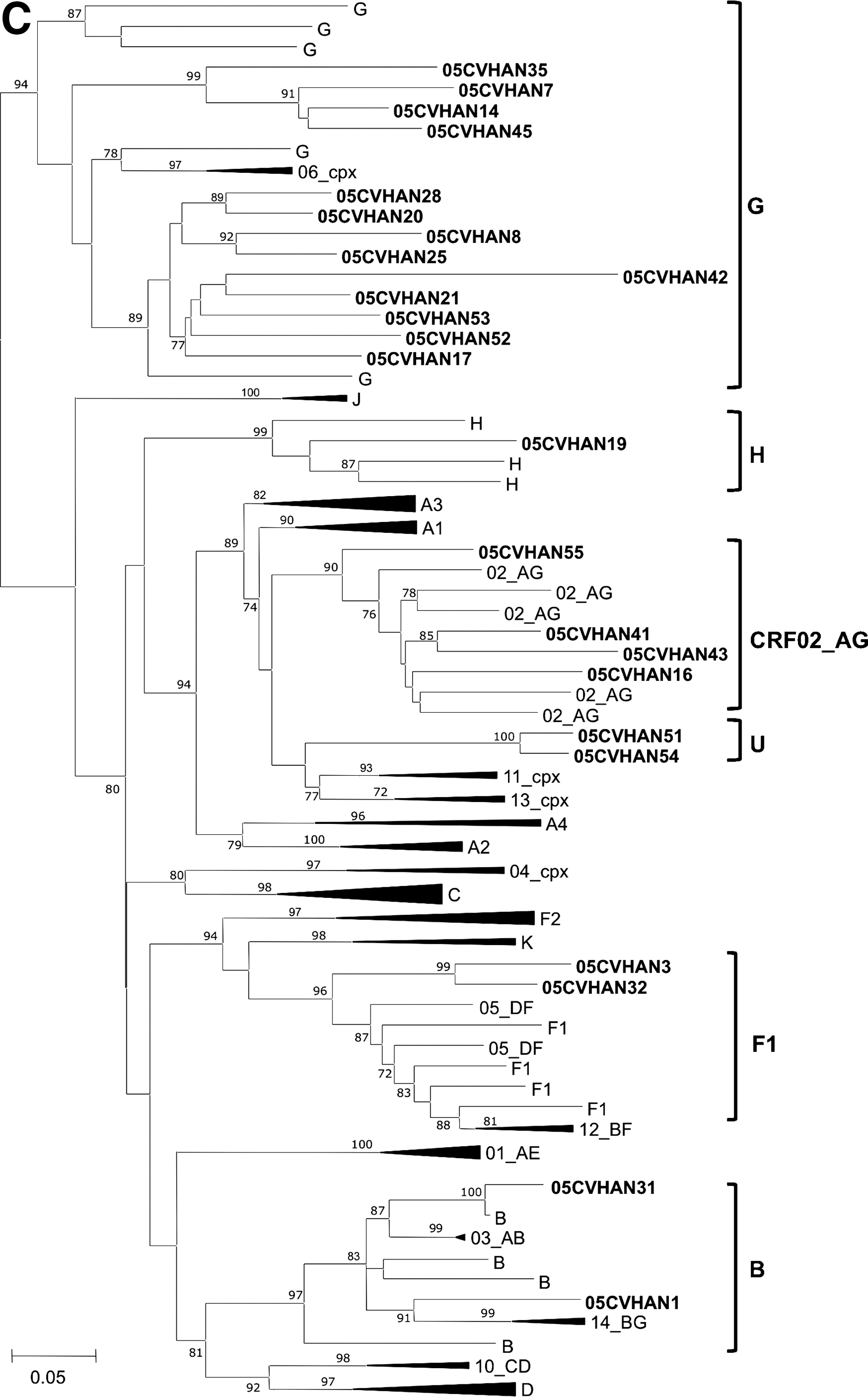
All HIV-2 isolates were classified as group A (Fig. 5). Eleven out of 14 isolates (76%) formed two clusters of unknown origin. Remarkably, however, isolates 05HANCV26 and 05HANCV36 revealed a strong evolutionary relationship with reference sequences from Guinea-Bissau and with the historic isolate HIV-2ROD. These results demonstrate that ROD-like strains are still circulating in this country (Fig. 5).
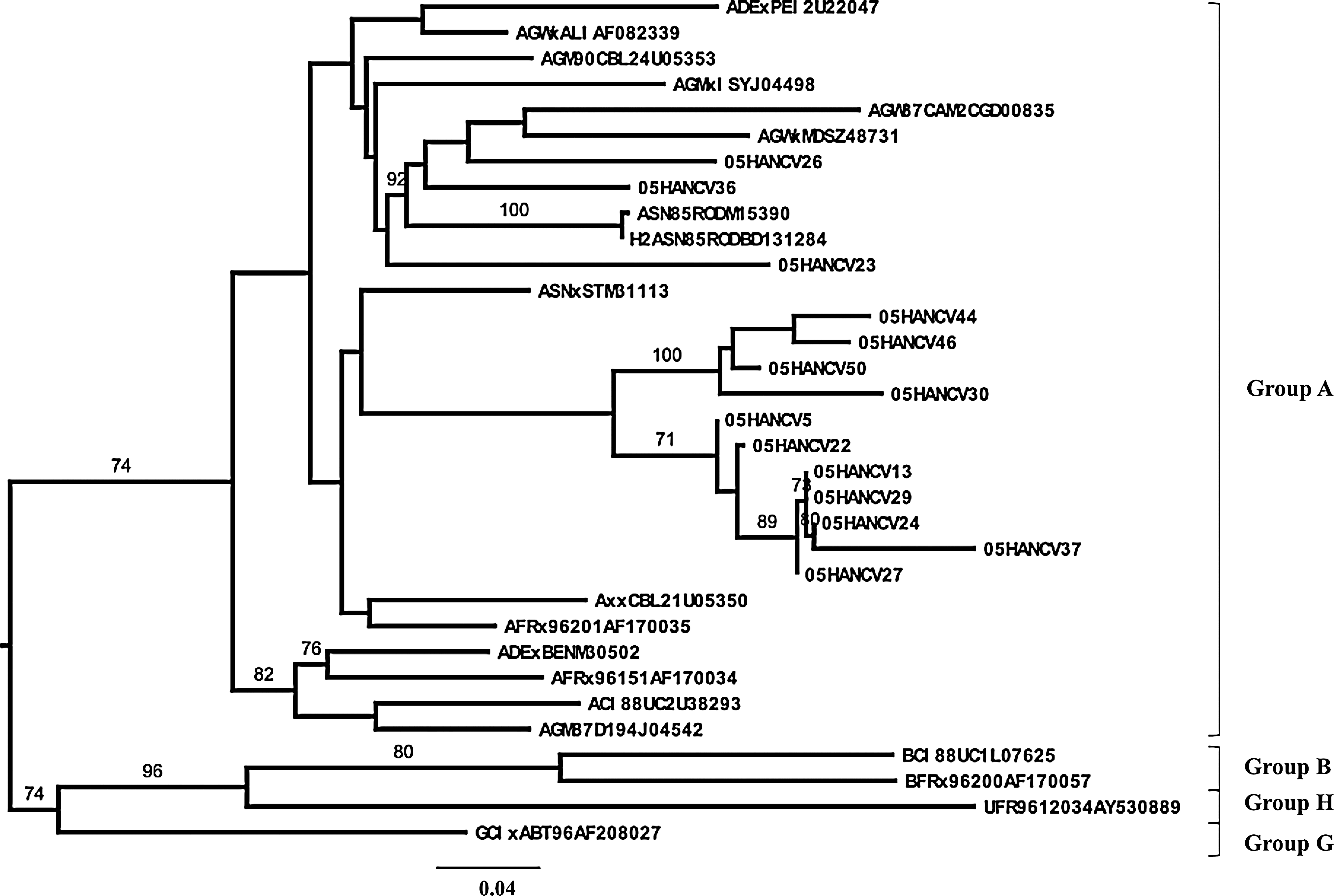
Genetic groups and evolutionary relationships of the HIV-2 viruses sequenced in this study based on maximum likelihood phylogenetic trees of the C2V3C3 env gene region. The phylogenetic tree was constructed with reference sequences from all HIV-2 groups. Sequences from Cape Verde isolates start with 05HANCV.
Discussion
This work is the first characterization of the HIV-1 and HIV-2 genetic forms present in Cape Verde Islands and the drug resistance profiles in treated and untreated patients. HIV-1 G was the prevailing subtype in our patients (48%), but most other subtypes and some untypable variants were also detected, either alone (subtypes B, F1, and C and CRF02_AG) or in recombinant forms with subtype G and/or CRF02_AG (B, C, H, and U). The introduction of different HIV genetic forms in the country is likely related with the intense population mobility motivated by emigration or with international tourism, an important economic activity in the country. The G strains circulating in Cape Verde were highly divergent when compared to the reference strains, which is consistent with multiple introductions of divergent G strains from different origins. 51 –56 Indeed, we found that G variants present in Cape Verde were imported mostly from Angola and Portugal where highly divergent subtype G strains prevail. 39,57 This was not surprising since Cape Verde has close historical, social, and economic relationships with these two countries.
Viral recombination occurs in geographic areas where several subtypes and CRFs circulate simultaneously in the population. In these regions, 8–47% of the infections may be due to intersubtype recombinants. 51,58 In this study 30% of the viruses were recombinant and 63% of them had at least one untypable genomic fragment. Of particular interest was isolate 05CVHAN42, which is subtype C in pol and G in env, a type of recombinant that has not been described before even though these two subtypes cocirculate in many African and European countries. 16,39,57,59 Full-length genomic sequencing analysis will be required to determine if this isolate is indeed the first representative of a new C/G recombinant strain.
The first HIV-2 cases were identified in 1985 in individuals from Guinea-Bissau and Cape Verde. 2 However, until now, no study has characterized the genetic diversity of HIV-2 in Cape Verde. We found that all HIV-2 isolates circulating in Cape Verde were phylogenetically related to group A. The A group is also endemic in several West African countries related to Cape Verde such as Guinea-Bissau, Ghana, Gambia, and Senegal. 60 –62 Most HIV-2 viruses circulating in Cape Verde share common ancestries and are unrelated to any of the reference isolates. Interestingly, however, two isolates were found to share a close evolutionary relationship with the historic HIV-2ROD strain, the first virus to be sequenced in 1987. 63 These results indicate that part of the long-standing HIV-2 epidemic in Cape Verde is self-renewed and suggest that the origin of the ROD isolate might have been Cape Verde. 2 In this sense, the sequences from the Cape Verde isolates are likely to provide important new insights into the origin and epidemic history of HIV-2. 64
The recent introduction of cART in Cape Verde prompted us to investigate drug resistance mutations in isolates from treated and untreated HIV-1-infected populations. We did not find major PI resistance mutations in either population. The minor mutations associated with PI resistance, L10I and A71V, were detected in two (6%) drug-naive patients. However, L10I occurs in 5–10% of untreated persons and A71V is a polymorphism that appears in 2–3% of untreated individuals. Therefore these mutations are not considered transmitted drug resistance mutations. 49,50,65
In contrast, we detected NRTI or NNRTI resistance mutations in three unrelated drug-naive patients infected with subtype G (05HANCV8, 05HANCV45) or a G/B recombinant (05HANCV31). M41L, present in patient 05HANCV31, confers intermediate-to-high level resistance to AZT and d4T and a lower level of resistance to ddI, ABC, and TDF. 50,66 K103N, present in patient 05HANCV08, causes high-level resistance to NVP and EFV. It has a synergistic effect with L100I on etravirine (ETR) susceptibility. 50,66 Y318F, present in patient 05HANCV45, causes low/intermediate NVP resistance. It is a rare mutation and, unlike M41L and K103N, it is not considered a transmitted drug resistance mutation. 48 Hence, overall, we found two cases of transmitted drug resistance in 17 patients (12%). The finding of a moderate prevalence of transmitted HIV-1 drug resistance in Cape Verde was a surprise as free access to cART was officially started at the end of 2004. 3 Despite the relatively high prevalence of drug-resistant strains found in treated individuals (30%), the K103N mutation was found in only one treated patient and M41L was not found in treated patients. Hence, the most probable explanations for the origin of these resistant isolates in Cape Verde are (1) the unregulated and unmonitorized use of antiretroviral drugs previous to 2004 and (2) people displacements from countries in which ART has been available for a longer period of time. 10,16
Resistance mutations were detected in 3 of 10 (30%) treated patients but there was clear evidence of virological failure in only one of these patients (05HANCV55). The isolate from this patient harbored mutations conferring resistance both to NRTIs (K65R, T69d) and NNRTIs (K103N, Y181C). K65R causes intermediate resistance to ddI, ABC, 3TC, FTC, and TDF, low resistance to d4T, and hypersusceptibility to AZT. 50,66 Mutations at codon 69 usually occur as T69DS/A/I/N/G and are NRTI-selected mutations. 50 T69d is a highly unusual mutation at this position. Deletions at codon 69 occur at a frequency of about 0.1% and their phenotypic and clinical significance are not known. Y181C confers high resistance to NVP and low resistance to EFV. 50 Moreover, mutations at position 181 compromise the ETR response and may provide the foundation for the development of high-level ETR resistance 50 . K103N confers full resistance to EFV and NVP. 50,67 Unfortunately, we had no access to the therapeutic regimen of this patient. However, based on the resistance genotype only, this patient is a clear candidate for a PI-based second line therapeutic regimen.
In conclusion, the long-standing HIV-2 epidemic in Cape Verde is partially rooted in the ROD strain, suggesting that the origin of this historic isolate was Cape Verde. The HIV-1 epidemic results from the introduction of multiple subtypes and recombinant forms by migrant workers or tourists. The prevailing and highly divergent HIV-1 subtype G has its origins in Portugal and Angola. We found a moderate prevalence of primary and secondary HIV-1 drug resistance in Cape Verde and evidence suggesting that most resistance isolates showing primary resistance mutations were imported. In the actual context of access to standardized and free therapeutic regimens, the development and implementation of drug resistance surveillance strategies in both treated and untreated populations will be important to maximize the efficacy of antiretroviral therapy and prevent further transmission of drug resistance isolates in Cape Verde.
GenBank Accession Numbers
Sequences have been assigned the following accession numbers: JF267400– JF267423 (HIV-1 env), JF267438–JF267463 (HIV-1 PR), JF267464–JF267490 (HIV-1 RT), and JF267424–JF267437 (HIV-2 env).
Footnotes
Acknowledgments
This work was supported by Fundação GlaxoSmithKline para as Ciências da Saúde, Portugal, Fundação para a Ciência e Tecnologia (project PTDCSAU-FCF6767/2006), Portugal, and CHAIN (Collaborative HIV and Anti-HIV Drug Resistance Network), European Union. Pedro Borrego, Cheila Rocha, and Inês Bártolo are supported by Ph.D. grants from Fundação para a Ciência e Tecnologia, Portugal.
Author Disclosure Statement
No competing financial interests exist.