
Abstract
Characterization of HIV subtypes can provide a more comprehensive understanding of the epidemic within a distinct region, and when combined with notification data, may also be helpful in enhancing current HIV prevention strategies. In this study, we characterized 1056 HIV-positive individuals (948 males and 108 females) living in Victoria and whose infection was detected for the first time between 2005 and 2010 inclusive. HIV-1 strains were subtyped based on pol gene sequence. Phylogenetic analysis was performed on all non-B subtype sequences identified. Of the 1056 sequences analyzed, 825 were subtype B and 231 were non-B. Overall 6 HIV-1 subtypes, 6 circulating recombinant forms (CRFs), and 12 unique recombinant forms (URFs) were identified. Regardless of gender, the majority of individuals were infected with a subtype B virus (78%). Subtype B was dominant in males (n=806, 85%). In contrast, the majority of females were infected with non-B subtypes (n=89, 82%), in particular subtype C (n=48, 45%). Phylogenetic analysis of the non-B subtypes revealed that the majority of clustering, and thereby transmission, occurred with CRF01_AE strains. Despite the relatively high numbers identified in females there was very little clustering of subtype C viruses. Subtypes C and A1 both historically associated with heterosexual transmission, and CRF01_AE often associated with IVDU, were also associated with transmission within the MSM population, demonstrating the potential for non-B subtypes to expand into the MSM population. The observation of increasing numbers of females and heterosexual males infected with non-subtype B viruses, the majority imported through migration and travel to countries where there is a high prevalence of HIV, suggests a targeted public health message may be required to prevent further increases within these two groups.
Introduction
T
Group M viruses, which are responsible for the majority of HIV-1 infections worldwide, have diversified further into nine phylogenetically distinct subtypes (A–D, F–H, J–K). 3 In addition to these genetic subtypes, circulating and unique recombinant forms (CRFs and URFs, respectively) occur as a result of recombination between two or more HIV subtypes. They show discrete geographic distribution patterns and contribute to up to 20% of HIV infections worldwide. 4 –6
When the first cases of HIV infection were detected in Australia in the mid-1980s, and for many years afterward, the majority were associated with subtype B virus infections in homosexual men. 7 Other risk groups, including bisexual men and intravenous drug users (IVDU), although much less common, also usually involved infection with subtype B strains. A small number of cases, including some linked phylogenetically, involved subtype CRF01_AE in IVDUs and heterosexuals. 8 Historically in Australia, very few females have been infected, with the most recent data (to December 2009) showing only 8.9% of a total of 29,395 infections in this country being of this gender. 9
In a previous study of HIV-1 subtypes circulating in Australia (in New South Wales), non-subtype B infections were generally acquired either overseas before travel to this country, via mother-to-infant transmission, through travel of Australian citizens to high prevalence countries, or through direct contact in Australia with foreign nationals. 10 Those cases that were sexually acquired were strongly associated with heterosexual transmission. As well as this direct evidence obtained through phylogenetic analysis, the potential for a diversity of subtypes in Australia was also predicted based on an increase in the number of new HIV diagnoses in females infected through heterosexual contact. 11
In Australia, approximately 1000 new cases of HIV are diagnosed annually, around 300 of them from Victoria. 9 We undertook a phylogenetic study of HIV subtypes circulating in Victoria between 2005 and 2010 involving cases detected for the first time as either recent infections or new diagnoses. As well as investigating the distribution of subtypes and the exposure risk associated with these infections, we also sought to determine whether the infecting strains are spreading through onward transmission. Evidence for changes in subtype distribution, including the risk groups associated with these subtypes and their transmission routes, requires a public health response different from that used in a setting that has previously mainly involved homosexual men.
Materials and Methods
Study population
The study was carried on people who were residents of Victoria, Australia, a state of approximately 5.6 million. With 4845 people estimated to be living with HIV in Victoria in 2010, the prevalence of HIV infection is approximately 86.5 per 100,000 with an annual incidence of approximately 290. 9 A total of 1056 HIV-positive individuals whose infection was detected for the first time between 2005 and 2010 inclusive were investigated (Table 1). Of these, 354 were diagnoses of newly acquired infection, where there was conclusive serological evidence that they had become infected within the preceding 12 months. The remaining 702 were new diagnoses of HIV infection confirmed by a full positive Western blot at first presentation. All cases were antiretroviral (ARV) drug treatment naive. Notification data obtained from the Department of Human Services (Victoria) were used to provide information on the likely mode of transmission of the virus.
Sequencing HIV pol for subtype assignment
Patients provided written consent for blood collection and sequencing of pol for drug resistance testing. Phylogenetics was performed anonymized from patient identifiers. The methods for amplification and sequencing individual pol sequences have been previously described. 12 Briefly, a 1035 base-pair product spanning the entire coding region for protease (PR) and the first 246 codons of reverse transcriptase (RT) was amplified from HIV-1-specific RNA derived from 500 μl of plasma, and sequenced using ABI Prism reagents, hardware, and software (Applied Biosystems, Foster, CA). HIV strains were subtyped based on their pol sequences using online programs such as Los Alamos 13 and the Viral Genotyping Tool at NCBI. 14 Subtype assignment was confirmed using the REGA HIV-1 subtyping tool version 2.0 15 and, if subtype assignment was unclear due to potential recombination, Recombinant Identification Program (RIP) version 3.1 16 and Jumping Program Hidden Markov Model (jPHMM). 17
Phylogenetic analysis
Sequences were manually aligned using BioEdit. 18 Phylogenetic reconstruction was performed using distance-based (F84) and neighbor-joining methods in the Phylip suite of programs. 19 Transition transversion ratio calculated using MEGA version 3.1 20 and gamma parameter alpha estimated using Tree Puzzle 21 were incorporated in the algorithm to optimize tree topology. Bootstrap analysis of the entire dataset was performed on 1000 replicates using MEGA version 3.1. 20 Trees were viewed and edited for publication purposes using FigTree version 1.3.1. 22 The average genetic distances between viral sequences within a cluster were estimated using MEGA version 3.1. 20
Results
Subtypes circulating between 2005 and 2010
The numbers of males and females investigated and the overall distribution of subtypes (B or non-B) over the 6-year period are shown in Table 1. A total of 948 males (89.8%) and 108 females (10.2%) were genotyped and analyzed phylogenetically, revealing 6 subtypes, 6 CRFs, and 12 discrete URFs (Table 2). Overall, the majority of individuals were infected with a subtype B virus (78.1%). Using the number of males as the denominator (n=948), subtype B was the most common subtype (85.0%), followed by CRF01_AE (7.5%), subtype C (4.3%), subtype A1 (1.1%), and <1% of subtypes D, G, and F1, CRF02_AG, CRF14_BG, CRF_BG, and CRF12_BF, and the URFs. In contrast, when using the number of females as the denominator (n=108) the majority were infected with a subtype C strain (44.4%), followed by CRF01_AE (20.4%), subtype B (17.6%), subtype A1 (4.6%), CRF02_AG (3.7%), subtype D (2.8%), subtype G (1.9%), and <1% for subtype CRF07_BC and the URFs. During the 6-year period, the percentage of non-B subtypes in females rose from 60% to 85% (with a peak of 95% in 2009) (Fig. 1). In terms of percentages, infection with non-B strains almost doubled in males (10% of all cases in 2005 to 19% in 2010), but numerically, this increase was less pronounced (Fig. 1).
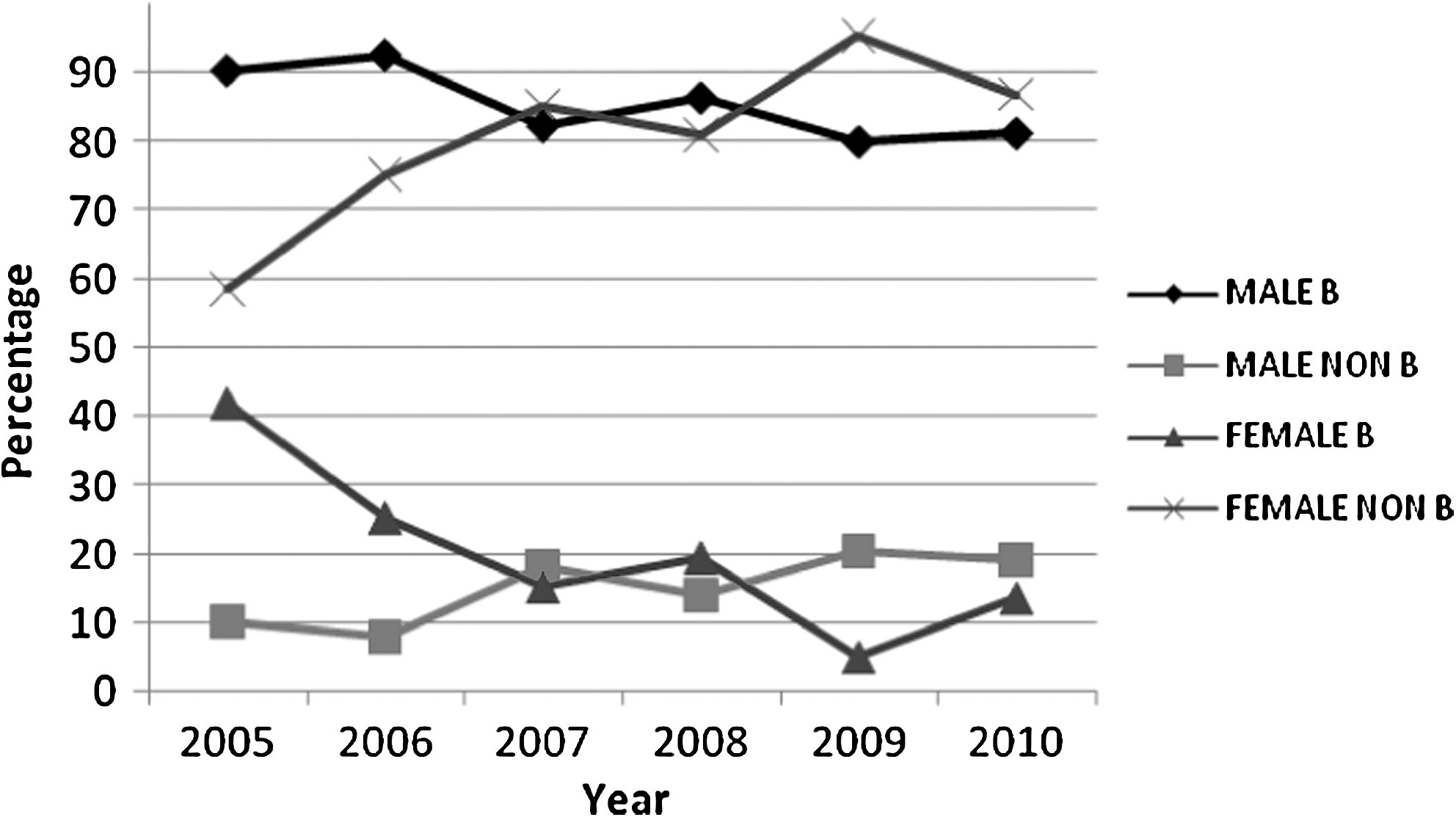
Temporal changes in subtype according to gender between 2005 and 2010.
HIV exposure risk by gender and subtype
Females
The likely exposure risks for females infected with both subtype B and non-B strains are shown in Fig. 2a. Nineteen females were infected with a subtype B virus, only one through drug use. Most females infected with subtype C viruses were exposed either in their country of origin or through heterosexual contact with a resident of a high HIV prevalence region, including sub-Saharan Africa (Zimbabwe, Botswana, Ethiopia, Sudan, Ghana, South Africa), Papua New Guinea, and East Timor. Despite CRF01_AE infection often being associated with IVDU in Australia, the majority of females infected with this subtype reported heterosexual contact as their transmission risk either in their country of origin or in Australia with a male from a high prevalence country (Vietnam, Cambodia, Burma, and Indonesia). Five females were infected with subtype A1, one from Uganda, a mother and daughter infected in Tanzania, and a mother and daughter infected in the Congo. CRF02_AG was identified in four females, two of whom were exposed in South Africa and Ghana. The three females infected with subtype D were from sub-Saharan Africa. Two females infected with subtype G were from Nigeria and the Congo. CRF07_BC was identified in a female infected in Burma. Four females were either exposed to or infected with one or more viruses of different subtypes that gave rise to URFs. Based on their pol sequence and analyses using different recombination identification programs these were categorized as URFs B/D/AG, A1/G, D/A1, and AE/B. Each of the infected women had migrated to Australia from countries in which HIV was prevalent, namely Ghana, Liberia, Uganda, and Malaysia, respectively.
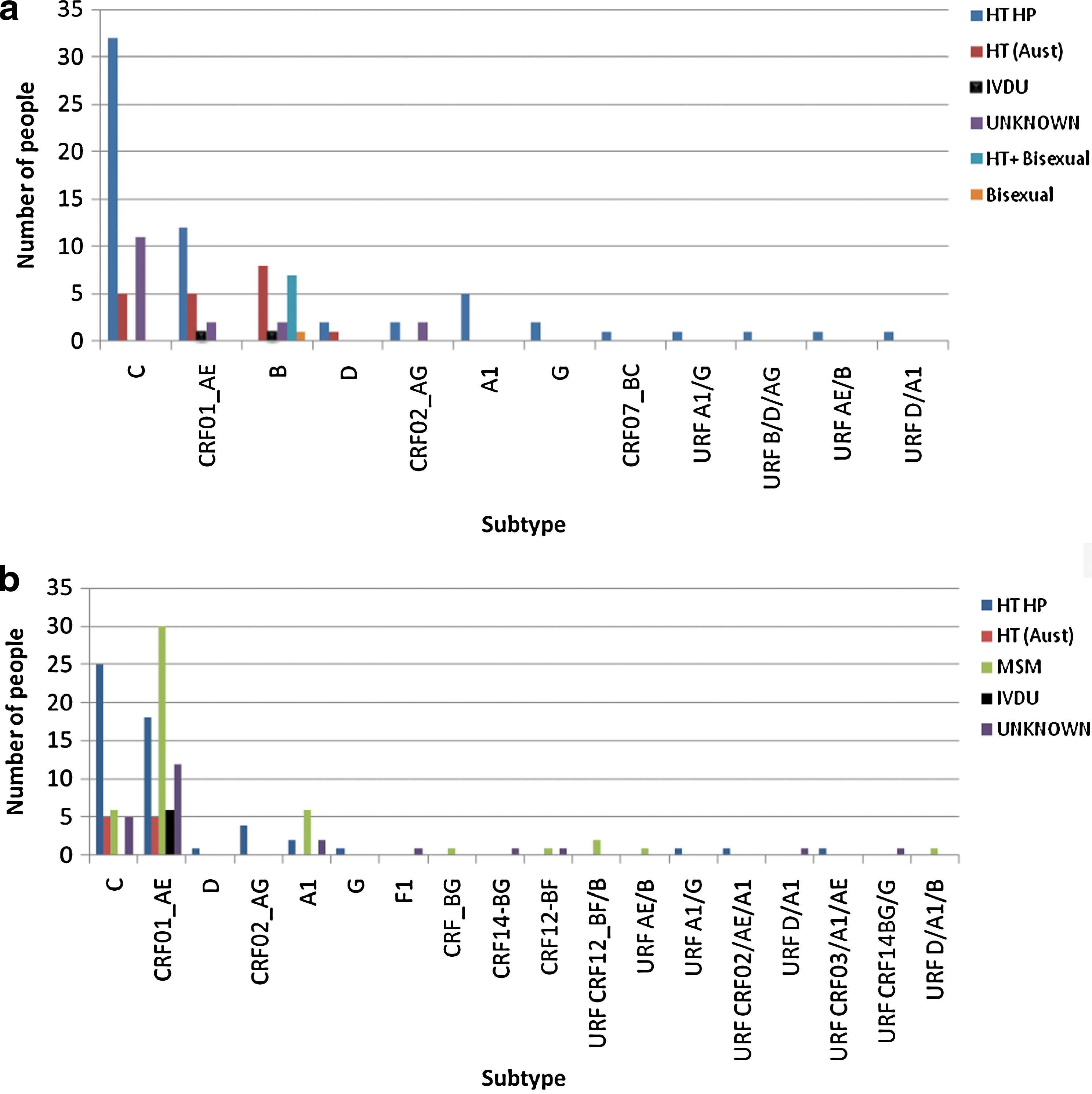
Males with non-B subtype infection
The majority of non-B subtype infections in males were CRF01_AE (n=71) (Table 2, Fig. 2b). Most of them (n=30, 42.3%) were men who have sex with men (MSM), 13 from high prevalence countries in Southeast Asia. Twenty-three (32.4%) followed heterosexual exposure with a female who had lived in a high prevalence country (including Cambodia, Indonesia, Vietnam, and Thailand). Six reported IVDU as their exposure risk. The majority of subtype C infections (n=25, 61%) were associated with heterosexual transmissions involving residents from high prevalence countries that included sub-Saharan Africa (Zimbabwe, Botswana, Sudan, Ethiopia, South Africa), Malaysia, and Indonesia. Six MSM were also infected with this subtype. Of the 10 individuals infected with subtype A1 strains, six were MSM (three whose viruses were closely related phylogenetically) (Fig. 3), two were exposed through heterosexual contact in the Congo, and two were siblings of a subtype A1-infected mother also from the Congo. Four males infected with CRF02_AG acquired their infection through heterosexual contact in Ghana, Indonesia, or Cambodia. The one (heterosexual) male with subtype D virus was infected in Sudan and the single male with subtype G was infected in Liberia. The pol sequence of an intersubtype recombinant CRF_BG in a local MSM aligned with a group of viruses recently identified among a group of MSMs in Havana, Cuba. 23 Of nine males infected with URFs, four of them were MSMs (including two Australian nationals). The URFs identified were two identical CRF12_BF/B sequences, one each of CRF01_AE/B and D/A1/B and one strain each of CRF02_AG/A1/CRF01_AE, CRF03_AB/A1/CRF01_AE, and A1/G identified in heterosexual males migrating from the Congo, Vietnam, and Sierra Leone, respectively. No information was available on the remaining two.
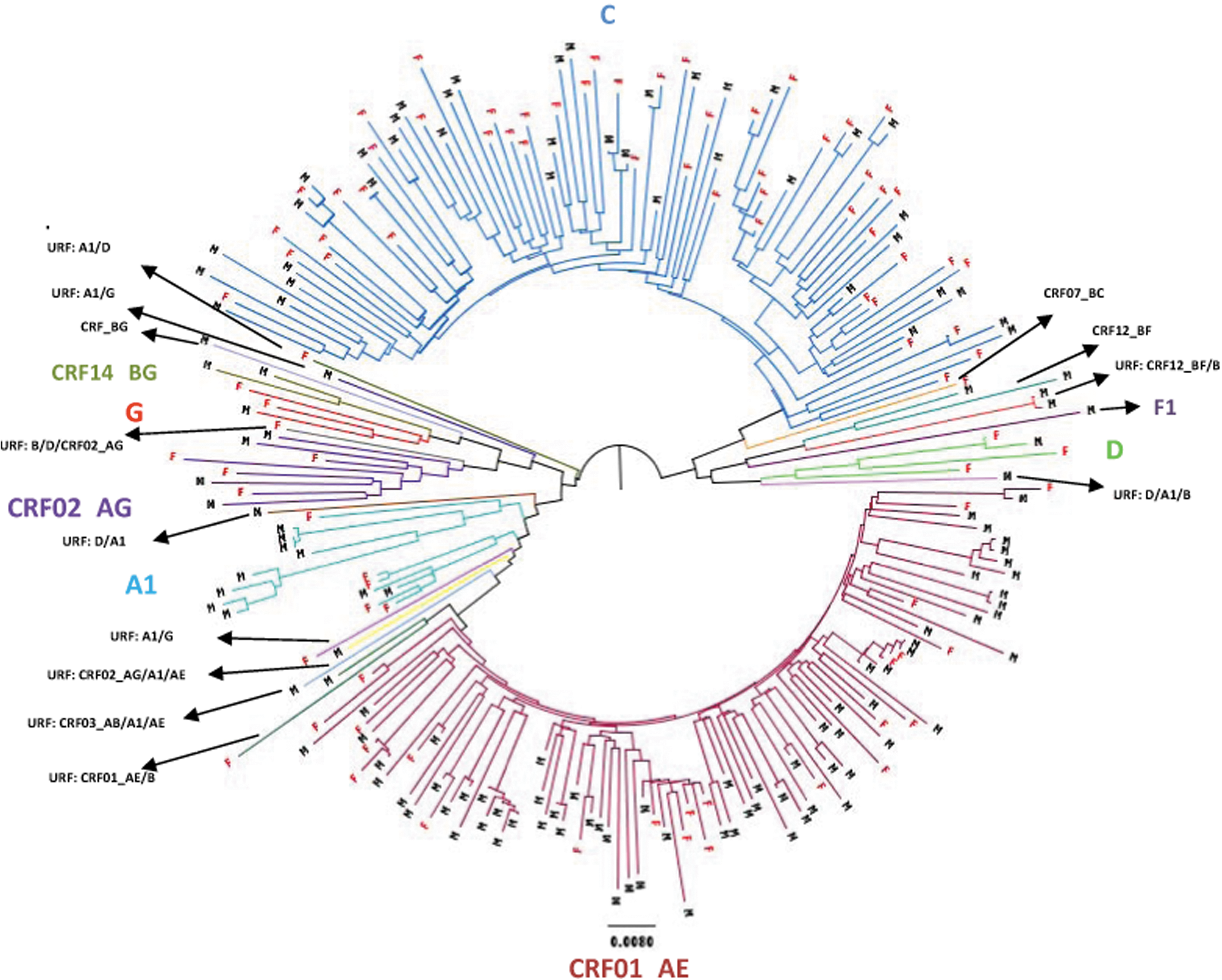
Phylogenetic tree consisting of 231 non-B HIV-1 viruses detected between 2005 and 2010 inclusive. Males (M) and females (F) are indicated.
Analysis of phylogenetic clustering of non-B subtypes linked with notification data
Phylogenetic analysis was performed on all 231 non-B subtype pol gene sequences. This reconstruction confirmed the initial subtype assignments obtained using online programs and aided in the identification and confirmation of URFs by showing both a distant genetic relationship and low support for clustering with a particular genotype (Fig. 3). It also identified several possible sexual networks within subtypes (C, CRF01_AE and A1) and a URF (CRF12_BF/B).
Of 93 CRF01_AE strains, 16 clusters involving 48 individuals (34 males and 14 females) were identified. The largest involved eight sequences and included four females from Indonesia and two heterosexual males infected in Indonesia and in Australia. One cluster containing six viruses (including two from females) were all linked to IVDU. Several clusters contained either four or five related viruses all infecting MSMs. One cluster of three related viruses involved two females who had acquired their infections in Vietnam and a male who had been diagnosed in New South Wales. Eleven clusters each containing two related viruses involved heterosexual couples (five clusters), pairs of MSM (five clusters), and one cluster linking two males, one a heterosexual and the other whose risk factor for transmission was not known.
Within the subtype C group, six subclades were identified. Notification data relating to the source country of the individuals associated with these subclades did not reveal an association between the geographic origin of the case and grouping with a particular subclade. Irrespective of gender, most subtype C infections did not have apparent virological linkages to other strains That is, their strains were not linked to other individuals in our set of sequences. Eight clusters involving a total of 18 individuals were identified amongst the 89 subtype C strains. Two clusters each containing three sequences were identified, one of which involved a female from East Timor, her son, and an Indonesian male, and the second involving a heterosexual male who acquired his infection in Australia and a male and female whose notification data were not available. The remaining six pairs of linked sequences each consisted of a male and a female, usually partners and usually immigrants from sub-Saharan Africa.
Four clusters involving 13 sequences were identified within the 15 subtype A1 infections detected. One cluster of four sequences involved a mother and her three children. A second cluster, also of four sequences, involved males, three of whom were local MSMs. A third cluster of three closely related subtype A1 viruses also involved MSMs, two who seroconverted during the same month and one who seroconverted 6 months earlier, suggesting these may represent cases of onward transmission. The final cluster of two sequences involved a mother and daughter.
Of the remaining non-B subtypes, two identical URFs (CRF12BF/B) were identified in MSMs, who were seroconverting simultaneously, and a cluster of two related subtype D sequences involving a heterosexual couple from sub-Saharan Africa. No evidence of clustering was observed in the three subtype G and 8 CRF02_AG strains identified.
Discussion
This study investigated the molecular diversity of HIV strains in a geographic location where HIV transmission has historically involved mainly subtype B infection in MSM. During the 6 years studied there were 1683 cases of newly diagnosed HIV infections in Victoria. 9 Of these, 1056 (63%) had baseline drug resistance genotyping performed as part of clinical care and the sequences from this testing provided the material for subsequent phylogenetic analysis. A previous study suggested that a growing proportion of these cases would be female and non-subtype B because of increasing migration from areas in which HIV infection is endemic. 11 Our results show this prediction was largely correct. However, our phylogenetic analysis also suggests that the non-B subtype strains infecting females are only rarely transmitted (Fig. 3).
The most genetically diverse viruses were identified in males (19 distinct genetic types compared to 12 in females) and were introduced directly through migration or as a result of an exposure in a high HIV prevalence country. The most diverse modes of transmission were noted in CRF01_AE, especially in males, where four recognized transmission modes were represented (Fig. 2b).
Although subtype B remained the predominant subtype in Victoria, especially in MSM, analysis of the non-B subtype infections (approximately 22% of all cases) revealed a diverse range of subtypes, CRFs, and URFs whose numbers increased annually from 2005. Compared to other Australian states and territories, the epidemic in Victoria is similar according to modes of acquisition, 24 gender infected, country of diagnosis, and (limited) subtype distribution. 9 In males, the majority of non-B subtype infections were, as previously described, the result of heterosexual contact with females in or from high prevalence countries. 25 In contrast to males, non-B subtype viruses predominated in females (82%), with subtype C the most common subtype identified.
We observed occasional variations to the common modes of transmission of several non-B subtypes. Subtype C infections, usually associated with heterosexual exposure in high prevalence countries in sub-Saharan Africa, were also identified regularly in the MSM population. CRF01_AE strains, predominant in Southeast Asia and previously associated with IVDU in Victoria, 8 were also found in MSM. Subtype A1, usually restricted to heterosexual contact in countries of sub-Saharan Africa, was also identified in six MSMs. Three of these men were infected within 6 months of each other and are likely to represent cases of onward transmission. Another likely case of onward transmission occurred in two local MSM infected with URF CRF12BF/B strains. These results demonstrate the potential for non-B subtypes to expand into the MSM population.
When the non-B subtypes were subjected to phylogenetic analysis, the majority of clustering was observed with CRF01_AE strains, and more than half of these could be linked. Within the 16 clusters identified, more than twice the numbers of males were involved in transmissions than females, and approximately half the males were MSM. The remainder were infected through heterosexual transmission and, in a small cluster of six involving males and females, through IVDU. Despite almost equal numbers of CRF01_AE and subtype C strains identified in the study, there were significantly fewer transmissions observed in the latter subtype. Females predominated within the subtype C viruses, and phylogenetic analysis showed that most sequences derived from them ended in terminal nodes, demonstrating that they rarely transmitted their infecting strain (Fig. 3).
Our observation of increased transmission of non-B subtypes CRFs and URFs into the MSM population highlights the need for manufacturers of HIV detection and quantitation assays to continue to validate their assays against all newly recognized HIV variants. The observation of an increasing number of females and heterosexual males involved in non-subtype B infections, the majority imported through migration and travel to countries where there is a high prevalence of HIV, suggests that a different public health message may be required to prevent further increases within these two groups. This includes increased awareness campaigns suggesting the need to test more regularly, emphasizing the prevention of transmission during pregnancy and the practicing of safe sex, targeted to patients with diverse cultures and languages.
Footnotes
Author Disclosure Statement
No competing financial interests exist.