
Abstract
To test tipranavir (TPV) or darunavir (DRV) as treatment options for patients with phenotypic resistance to protease inhibitors (PIs), including lopinavir, saquinavir, atazanavir, and fosamprenavir, the PhenoSense GT database was analyzed for susceptibility to DRV or TPV among PI-resistant isolates. The Monogram Biosciences HIV database (South San Francisco, CA) containing 7775 clinical isolates (2006–2008) not susceptible to at least one first-generation PI was analyzed. Phenotypic responses [resistant (R), partially susceptible (PS), or susceptible (S)] were defined by upper and lower clinical cut-offs to each PI. Genotypes were screened for amino acid substitutions associated with TPV-R/DRV-S and TPV-S/DRV-R phenotypes. In all, 4.9% (378) of isolates were resistant to all six PIs and 31.0% (2407) were resistant to none. Among isolates resistant to all four first-generation PIs, DRV resistance increased from 21.2% to 41.9% from 2006 to 2008, respectively, and resistance to TPV remained steady (53.9 to 57.3%, respectively). Higher prevalence substitutions in DRV-S/TPV-R isolates versus DRV-R/TPV-S isolates, respectively, were 82L/T (44.4% vs. 0%) and 83D (5.8% vs. 0%). Higher prevalence substitutions in DRV-R/TPV-S virus were 50V (0.0% vs. 28.9%), 54L (1.0% vs. 36.1%), and 76V (0.4% vs. 15.5%). Mutations to help predict discordant susceptibility to DRV and TPV in isolates with reduced susceptibility to other PIs were identified. DRV resistance mutations associated with improved virologic response to TPV were more prevalent in DRV-R/TPV-S isolates. TPV resistance mutations were more prevalent in TPV-R and DRV-S isolates. These results confirm the impact of genotype on phenotype, illustrating how HIV genotype and phenotype data assist regimen optimization.
Introduction
Direct comparisons of HIV genotypes and in vitro phenotypes can provide useful information to further guide the development of resistance prediction algorithms and ultimately help to guide treatment decisions. However, routine evaluations of HIV genotypic and phenotypic data from patient-derived isolates are uncommon, 1 –3 and these analyses are generally limited to an individual drug or involve only a small number of patients, 4 thus limiting their overall utility. Furthermore, the emergence of mutations that confer resistance to protease inhibitors (PIs) in the context of background substitutions in the protease enzyme itself has been reported, 5 but few large-scale examinations of the relationship between genotype and phenotype across multiple antiretroviral drugs in the same class have been conducted to date. 6,7 A decline in the prevalence of resistance mutations to reverse transcriptase and protease inhibitors [nucleoside reverse transcriptase inhibitors (NRTIs), non-NRTIs (NNRTIs), and PIs] has been reported in a recent study of phenotypic resistance mutation patterns using patient-derived isolates from routine clinical samples collected between 1998 and 2007. 8
Decisions regarding the next regimen to use for an antiretroviral therapy-experienced, HIV-1-infected patient require an assessment of resistance. This assessment may involve genotypic sequencing of the viral population circulating in the peripheral blood or an assessment of both genotype and phenotype. Evaluations based on phenotypic testing are attractive when considering the choice of PI in a PI-experienced patient because the complex patterns of mutations in treatment-experienced patients make interpretations based on genotype alone extremely difficult, despite the development of a number of different interpretative algorithms.
The combination phenotype–genotype test (PhenoSense GT; Monogram Biosciences, South San Francisco, CA) includes both the phenotypic resistance measure and a prediction of the susceptibility of the isolate based on its genotype. The analysis presented in this report was derived from 7775 clinical isolates with matched genotypic and phenotypic data for the years 2006 and 2008. To be included in this analysis, each isolate was required to show reduced susceptibility [i.e., partially susceptible (PS) or resistant (R)] to at least one of four first-generation PIs. The dataset was queried to see what resistance patterns were observed and how those patterns might have changed during that timeframe.
Darunavir (DRV) and tipranavir (TPV) were both developed as agents capable of treating patients who already harbored resistance to one or more PIs. The objectives of the present study were to identify patterns of phenotypic susceptibility to TPV and DRV in this commercial database and to identify amino acid substitutions associated with samples having divergent phenotypic susceptibility to these two agents. These findings add new insights for clinicians who are faced with difficult therapeutic decisions for treatment-experienced patients, specifically those with PI-resistant virus.
Materials and Methods
Laboratory methods
Assays for phenotypic and genotypic drug susceptibility were performed at a commercial laboratory (Monogram Biosciences, South San Francisco, CA), as described previously. 9 –11 Phenotypic outcomes were categorized according to previously established values for the lower clinical cut-off (LCO) and the upper clinical cut-off (UCO). 12,13 Each cut-off was derived by analyses of virologic outcomes in clinical trial datasets where the specific PI boosted with ritonavir (r) was used as a new agent and after accounting for the impact of background therapy. A value below the LCO indicates a fully responsive, susceptible (S) virus. A value above the UCO indicates a nonresponsive, R virus. A value between the LCO and the UCO indicates a partially responsive, partially susceptible virus. Values between the LCO and UCO demonstrate residual drug activity that diminishes the closer the fold-change (FC) value is to the UCO. For TPV/r, the LCO is 2 FC and the UCO is 8 FC. 12,13 Similarly the following LCO/UCO was used for this analysis: atazanavir (ATV)/r at 5.2/30, lopinavir (LPV)/r at 9/55, saquinavir (SQV)/r at 2.3/12, fosamprenavir (FPV)/r at 4/11, and DRV/r 14 at 10/90. The selection criteria for this analysis included any specimen with >LCO for at least one of the four first-generation PIs or for DRV. Exactly 1 of the 7775 isolates qualified, based on susceptibility to DRV and all the first-generation PIs.
Genotypic characterization was made using pol genotypic data derived from the same test vector used to generate the phenotypic data. Genotypic resistance calls used previously defined algorithms. These algorithms were derived from phenotypic–genotypic correlations of isolates within the database and that were distributed around the LCO. 9,10,15
Sample population
The complete commercial database includes more than 110,000 commercial samples with matched genotypic and phenotypic information. It does not include samples from clinical trials and so is independent of the databases used to determine clinical cut-offs. The database population was queried for patterns of resistance over the period 2006 through 2008 to four widely used PIs (i.e., ATV, FPV, LPV, and SQV) along with patterns of resistance and cross-resistance with the two most recently introduced second-generation PIs: DRV and TPV. Clinical isolates with both phenotypic and genotypic test results available (n=7775) were selected from subtype B commercial samples submitted between 2006 and 2008, based on phenotypic drug resistance to PIs. All isolates demonstrated some drug resistance by having a phenotypic result above the LCO for at least one of the four boosted PIs (ATV, FPV, LPV, and SQV).
Statistical methods
Results were reported descriptively as rates and proportions. Changes in rates over time were tested using the Cochran–Armitage test for trend.
Results
Phenotypic resistance assessments over time and cross-resistance
As seen in Fig. 1A, the rate of phenotypic resistance (defined as an FC greater than the UCO) to TPV actually declined from 2006 (22.2%) to 2008 (18.5%). However, resistance to DRV increased significantly (p<0.0001), nearly doubling from 2006 (6.8%) to 2008 (11.6%). This trend is even more apparent when subgroups of resistant isolates (isolates resistant to two or more first-generation PIs) were analyzed (Fig. 1B–D). Low levels of resistance were seen for second-generation PIs among isolates exhibiting resistance to one or none of the first-generation PIs. These rates did not change over the 3-year period analyzed for this study (data not shown).
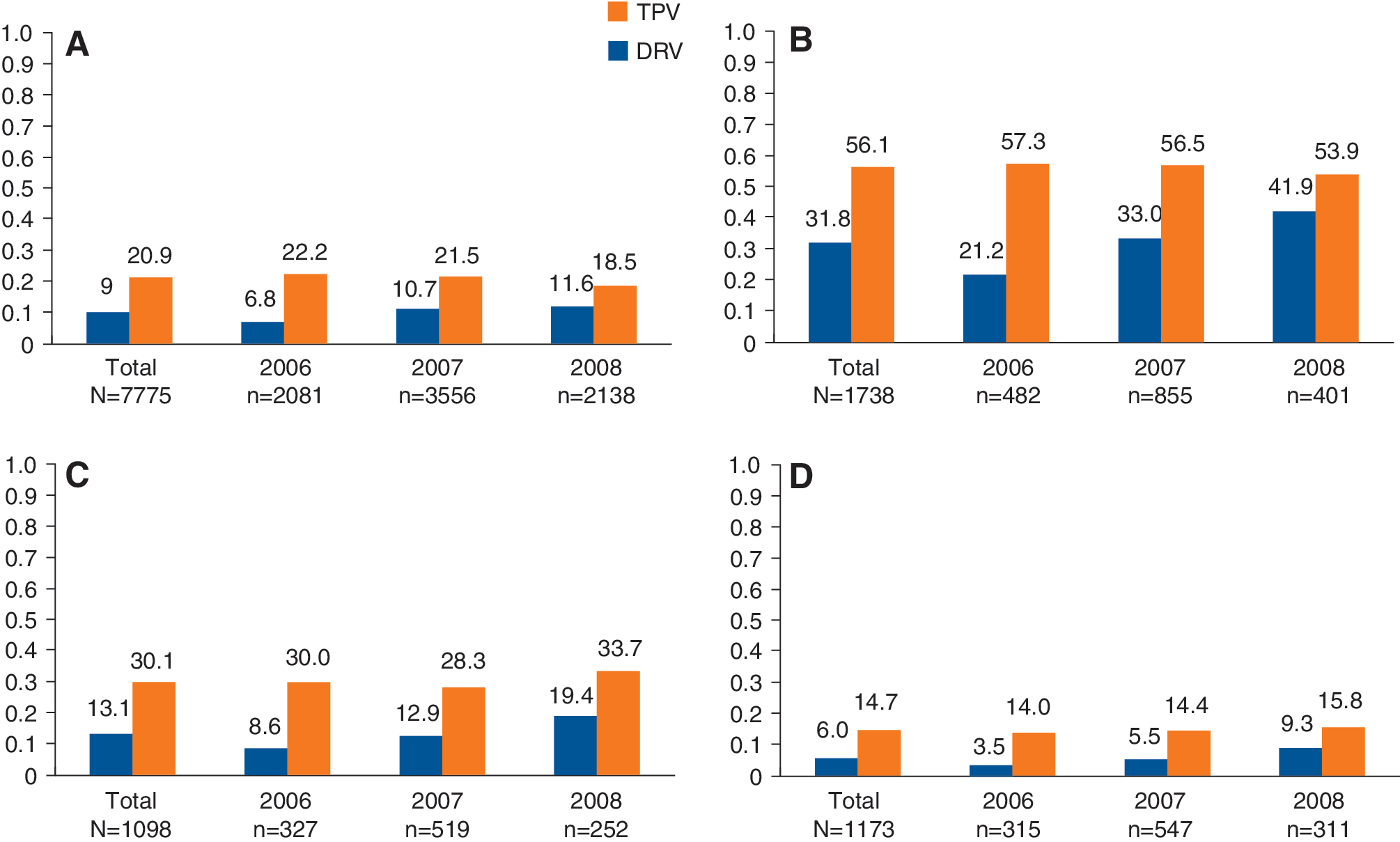
Rates of phenotypic resistance to second-generation protease inhibitors (PIs) tipranavir (TPV) and darunavir (DRV) from 2006 to 2008 as a function of resistance to first-generation PIs, lopinavir, saquinavir, atazanavir, and fosamprenavir. Proportion of isolates resistant to
Samples had to have reduced susceptibility in at least one PI in order to be included in the study, and 31.0% were resistant to none of the six evaluated PIs. Only 4.9% of the clinical isolates included in this study were resistant to all six evaluated PIs. Even among viruses that were resistant to the majority (at least three of the five) of the other PIs, 1592 of 2906 (54.8%) clinical isolates remained partially or fully susceptible to TPV. In the 552 patients whose virus was resistant to all five of the other PIs, TPV was predicted to be a viable therapeutic option (S or PS) in 174 (31.5%) patients; furthermore, 16.1% of patient isolates not susceptible to any of the other five PIs were still susceptible to TPV. Also, 37.5% of patient isolates susceptible to at least one of the other five PIs were also susceptible to TPV.
Assessment of mutations associated with phenotypic resistance to DRV and TPV
A number of resistance-scoring algorithms have been developed based on changes in the baseline sequence and their associated effects on phenotypic or clinical responses. These scores have revealed that the pathways to the emergence of resistance to both agents can be complicated, often involving clusters of amino acid changes and a number of unique substitutions. 16
The results of the present study confirm previously reported associations between specific mutations and either reduced or improved clinical response to TPV. 17 Mutations associated with a reduced clinical response were most commonly found in the isolates with phenotypic resistance to TPV, whereas those associated with improved response were most commonly found in the isolates that were susceptible (Figs. 2 and 3). Of the mutations associated with reduced clinical response, more than 80% of isolates that encoded the 82L mutation were TPV resistant (TPV-R), followed by 82T (58.6%), 83D (52.6%), 74P (49.9%), and 47V (42.1%). In contrast, the mutations associated with improved response were most frequently found in TPV-sensitive isolates 18 : 50V [79.7% of isolates with that mutation were susceptible to TPV (TPV-S)], 50L (74.9%), 76V (63.1%), 24I (61.0%), and 54L (50.4%) were all most commonly found in isolates that were susceptible to TPV.
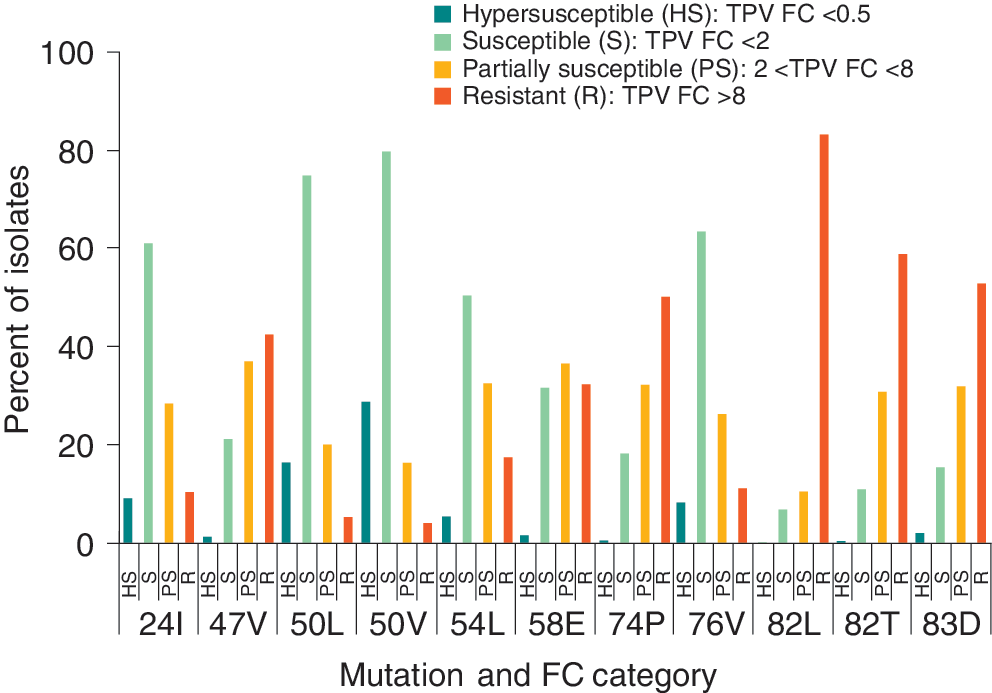
Occurrence of key mutations across tipranavir (TPV) phenotypic resistance classifications. The denominator for all participants is the number of isolates with that mutation and the numerator is the number of isolates in that phenotypic classification grouping with that mutation.
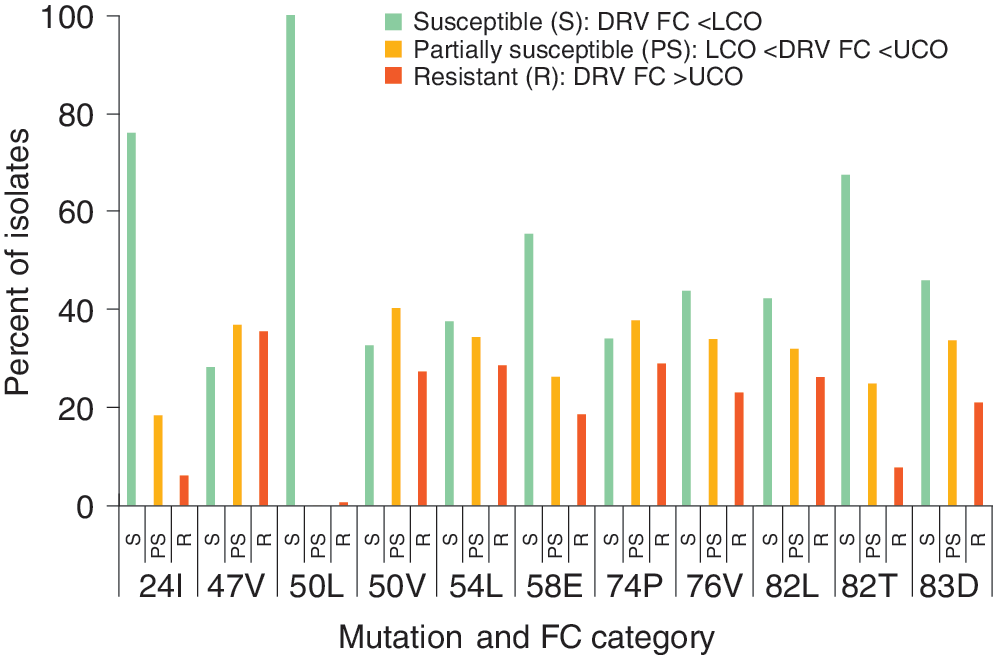
Occurrence of key mutations across darunavir (DRV) phenotypic resistance classifications The denominator for all participants is the number of isolates with that mutation and the numerator is the number of isolates in that phenotypic classification grouping with that mutation.
For DRV, the mutations associated with resistance to DRV all appeared at about the same rate across the phenotypic resistance categories. Isolates with major TPV resistance-associated mutations, such as 58E, 82T, and 83D, were commonly found in patients with virus predicted to be sensitive to DRV.
Patterns of mutations over time show that the cross-resistance patterns to DRV and TPV have remained fairly constant (Table 1), with 40.2% (2006, 40.2%; 2007, 39.1%; and 2008, 42.0%) of isolates sensitive to both TPV and DRV. Overall, 513 (6.6%) isolates were sensitive to DRV and resistant to TPV (DRV-S/TPV-R), with a slight decrease observed over time (8.1%, 6.4%, and 5.5% for 2006, 2007, and 2008, respectively; p=0.0007), whereas 97 (1.2%) isolates were resistant to DRV and sensitive to TPV (DRV-R/TPV-S), with a slight increase observed over time (0.9%, 1.3%, and 1.4% over those same years; p=0.1174).
Cross-Tabulation of Darunavir and Tipranavir Phenotypic Resistance Categories over Time
TPV, tipranavir; FC, fold change; DRV, darunavir; LCO, lower clinical cut-off; UCO, upper clinical cut-off.
For patients with discordant in vitro phenotypic resistance status for DRV and TPV, the prevalences of mutations associated with improved or reduced clinical response to TPV/r-based therapy were tabulated. As seen in Table 2, mutations associated with an improved clinical response to TPV were much more prevalent in DRV-R/TPV-S patients, e.g., 50V was found in 60.8% of DRV-R/TPV-S patients vs. 0.0% of DRV-S/TPV-R patients. Similar trends were observed for other such mutations: 54L (48.5% vs. 1.0%) and 76V (22.7% vs. 0.4%). The mutation 24I that predicts increased response to TPV was least prevalent with DRV-S/TPV-R (4.1%) and most prevalent with DRV-R/TPV-S (12.4%). Likewise, there were similar prevalence differences for DRV-PR/TPV-S patients. These mutations predict diminished response to DRV and increased response to TPV. Equally, mutations associated with an inferior clinical response to TPV were most commonly found in viruses sensitive to DRV and resistant to TPV, DRV-S/TPV-R versus DRV-R/TPV-S, respectively, e.g., 82L (7.4% vs. 0.0%), 82T (37.0% vs. 0.0%), and 83D (5.8% vs. 0.0%).
Prevalences of Tipranavir-Associated Mutations in Tipranavir/Darunavir Phenotypically Discordant Patients
Based on De Meyer et al. 24
Based on Scherer et al. 25
DVR, darunavir; TPV, tipranavir; DRV-S, darunavir-susceptible; TPV-R, tipranavir-resistant; TPV-PR, tipranavir-partially resistant; DRV-PR, darunavir-partially resistant; TPV-S, tipranavir-susceptible; DRV-R, darunavir-resistant.
The mutation 50L that predicts increased response to TPV and is neutral with respect to DRV was observed exclusively in DRV-S and TPV-(R or PR) isolates (N=85; 3.8%). The mutation 47V that predicts diminished response to both TPV and DRV was observed in a higher proportion of TPV-sensitive isolates (25.2% vs. 9.2%, respectively). The mutation 58E that predicts diminished response to TPV and is not a primary mutation for DRV was observed in an equal proportion of TPV-S and DRV-S isolates (19.8% vs. 19.6%, respectively). The mutation 74P that predicts diminished response to TPV and is neutral for DRV was observed in similar proportions of TPV-S and DRV-S isolates (5.8% vs. 4.8%, respectively).
Discussion
TPV and DRV are both approved for use in patients infected with HIV-1 that is resistant to multiple PIs. Results from previous studies have shown these agents to have little cross-resistance to first-generation PIs and unique resistance profiles. 19 –21 More recently, in a study comparing the correlations for 46 protease mutations between pairs of all eight currently approved PIs, the TPV and DRV pair had the lowest correlation coefficient (0.01) among the 46 mutations of all 28 pairwise comparisons. 22 Thus, utilizing patient-specific HIV genotype and/or phenotype data can significantly improve the drug selection process for a particular patient. The analyses described in these studies have identified a set of HIV mutations that can help identify isolates with different sensitivities to TPV and DRV. These results are consistent with what has been shown in the TPV-weighted score development. 23
At least one PI remained an option for treatment in 95% of patients seeking both phenotypic and genotypic resistance testing in 2006 to 2008. Even with resistance to the majority of the other PIs, 54.8% of the clinical isolates remained partially or fully susceptible to TPV. Susceptibility or partial susceptibility to TPV was observed in 31.5% of those resistant to all four first-generation PIs and DRV. For viruses exhibiting partial susceptibility to other proteases and susceptibility to TPV, consideration might be given, therefore, to the use of TPV in a subsequent antiretroviral regimen. A key observation from this analysis was that while resistance to TPV decreased slightly from 2006 to 2008, resistance to DRV nearly doubled during the same timeframe, most likely due to its wider use.
The results here are based on the clinical isolates sent to one laboratory for a combination of genotypic and phenotypic testing. Doctor and patient judgments about when to include phenotypic as well as genotypic assessment have therefore determined the population for this study. Insurance coverage will also have limited the availability of samples for inclusion. Although the clinical history is undocumented, it is assumed that patients whose samples were being sent for phenotypic assessment were receiving adequate care, and that the samples were therefore sent from patients who had failed therapy recently. Nevertheless, all of these factors represent limitations associated with database analyses of this type, and should be considered when interpreting the study findings.
The historical resistance patterns for TPV versus DRV have shown little cross-resistance, and this new analysis highlights a few mutations that confer discordant resistance phenotypes to the two agents. Mutations at 58E, 82L/T, and 83D are highly associated with poor clinical responses to TPV, whereas these isolates are still susceptible to DRV. However, mutations at 50V, 54L, and 76V are associated with a good clinical response to TPV and with resistance to DRV. These results are supported by previous work relating genotype to response for both drugs.
Discordant resistance to TPV and DRV has also recently been reported in a laboratory strain of HIV that was selected after 50 passages with the novel PI, GRL-216. This strain harbors the L10I, L24I, M46L, V82I, and I84V mutations, leading to a diminished response to DRV (3- to 5-fold increase in EC50), and a 10-fold increase in sensitivity to TPV. 18
The present study identified unique mutations that can help predict discordant phenotypic resistance in patients whose testing is limited to genotyping, reinforcing the belief that many of the substitutions associated with either TPV or DRV resistance are not associated with cross-resistance between the two agents. Since resistance to TPV is based on a unique set of mutations that is distinct from those that confer resistance to the other PIs, these data indicate that TPV is a rational next choice therapy for patients who have developed resistance to other PIs.
Footnotes
Acknowledgments
Editorial and writing assistance was provided by José L. Walewski, Ph.D., of Envision Scientific Solutions, Inc., which was contracted by Boehringer Ingelheim for these services. The authors meet criteria for authorship as recommended by the International Committee of Medical Journal Editors (ICMJE) and were fully responsible for all content and editorial decisions, and were involved at all stages of manuscript development.
Author Disclosure Statement
The studies described in this article were supported by Boehringer Ingelheim (Canada) Ltd., who is the manufacturer of Tipranavir.