
Abstract
We recently reported a novel adhesion pathway in lymphocytes that is mediated by cyclin-dependent kinase (Cdk) 4 activity and mediates lymphocyte interactions with endothelial matrix. We now demonstrate that HIV-infected lymphocytes also use Cdk4 to mediate spontaneous adhesion to fibronectin and endothelial matrix. We further demonstrate that HIV-infected lymphocytes require Rap-1 activity for phorbol-stimulated adhesion. Understanding adhesion pathways used by HIV-infected lymphocytes may lead to interventions to regulate aberrant adhesion and migration.
L
We recently reported a new adhesion pathway that is important in posttrafficking, i.e., mediating lymphocyte interactions with and migration through subendothelial extracellular matrix. 1 We showed that leukocytes could adhere to high-density fibronectin or exposed endothelial matrix in the absence of exogenous stimulation. We found several novel features of this pathway that clearly distinguish it from “conventional” phorbol ester-stimulated adhesion: it is dependent on Cdk4 activity, it requires microtubules, it does not require exogenous lymphocyte activation, and it does not require the small GTPase Rap-1 activity. 1 Because this novel pathway allows lymphocyte adhesion to physiologically relevant substrates such as exposed endothelial matrix in the absence of exogenous stimulation, we have termed it “ligand-induced adhesion” (LIA). We demonstrated a role for Cdk4-mediated adhesion in lymphocyte recruitment following lung injury and in thymocyte maturation and adhesion. 1,2
Cyclin dependent kinases (Cdk) are serine/threonine kinases that regulate cell cycle progression. Because of the role of Cdks in cell proliferation, much interest has focused on the development of pharmacological Cdk inhibitors as a therapeutic for cancer. Several studies have demonstrated that Cdk inhibitors can also inhibit replication of several viruses, 3 including HIV. 4 –6 Of additional interest, Cdk9, the catalytic subunit of positive transcription elongation factor b (P-TEFb), is part of the TAK complex (Tat-associated kinase complex), and binds to Tat protein of HIV, suggesting a possible role for Cdk9 in AIDS progression. 7 Therefore, pharmacologic inhibitors of Cdks have been proposed as therapeutic agents for HIV infection. 8 –10
We questioned whether HIV-infected lymphocytes use the Cdk4-mediated pathway for adhesion, and if so, what the contributions of the different adhesion pathways in lymphocyte adhesion are in HIV-infected cells.
We first compared the adhesion of HIV-infected lymphocytes and mock-infected lymphocytes to high-density fibronectin without phorbol stimulation. Primary blood lymphocytes (PBL) were isolated and infected as previously described. 11 –13 As controls, cells were stimulated with PHA alone (“mock-infected”). Infection was monitored by measurement of p24 Ag (Coulter HIV-1 P24 antigen assay kit). Cells were used 7 days after infection. Adhesion assays were performed as previously described. 1 We found HIV-infected lymphocytes had the same adhesion profile to fibronectin as mock-infected cells (Fig. 1A), demonstrating that HIV-infected lymphocytes are similarly capable of ligand-induced adhesion. In addition, HIV-infected Jurkat T cells exhibited similar adhesion to fibronectin as mock-infected Jurkat cells (Fig. 1B). As expected, phorbol ester stimulation of lymphocytes with phorbol dibutyrate (PDBu) increased adhesion to fibronectin (Fig. 1C and D), which was similar in infected versus control lymphocytes. We also examined the spontaneous adhesion of HIV-infected lymphocytes to a more physiologic substrate, human umbilical vein endothelial cell (HUVEC)-derived matrix (Fig. 1D). Similar to our results with fibronectin, we found that HIV-infected and control lymphocytes adhered similarly to HUVEC matrix.

To determine whether HIV-infected lymphocytes required Cdk activity for spontaneous adhesion, we tested the effects of the Cdk inhibitors roscovitine and purvalanol A on adhesion to fibronectin and HUVEC matrix. Treatment of HIV-infected lymphocytes with either Cdk inhibitor significantly decreased adhesion to fibronectin (Fig. 1C) or HUVEC matrix (Fig. 1D). Reduction of inhibition was similar in HIV-infected and mock-infected lymphocytes. In contrast, the Cdk inhibitors had no effect on phorbol ester-stimulated adhesion of HIV-infected or mock-infected lymphocytes (Fig. 1C and D). Of note, since Cdk inhibitors can target additional pathways, 14 we previously demonstrated that inhibitors of other pathways, including PI3 kinase inhibitor (LY294002), MAPK inhibitor (PD98059), or tyrosine kinase inhibitor (genistein) had no effect on ligand-induced adhesion. 1 We also demonstrated that Cdk4, but not Cdk2, Cdk5, or Cdk6 is involved in ligand-induced adhesion using dominant negative constructs, siRNA knockdown, and thymocytes from Cdk4-/- mice. 1,2
The identification of Cdk4 as the relevant substrate was somewhat surprising since roscovitine has been shown to have limited direct effect against Cdk4. 15 –19 It is possible that Cdk4 inhibition by roscovitine was indirect, i.e., through the down-regulation of the cyclin partners or other Cdks that are required for Cdk4 activity. 20 In addition, the absence of phosphorylation of reported Cdk4-specific sites on Rb in the presence of roscovitine may be due to regulation by other Cdks. 21,22
Therefore, to confirm our previous work demonstrating the involvement of Cdk4 in ligand-induced adhesion, 1 we tested the ability of Cdk4 dominant-negative (DN) construct to inhibit ligand-induced adhesion in HIV-infected lymphocytes. Jurkat cells were stably transfected with a Cdk4 DN expression vector that contained a mutation D145N that prevents its activity. 23 We confirmed construct expression and inhibition of phosphorylation of Rb, downstream target of Cdk4 (Fig. 1G). After HIV infection, Cdk4 DN reduced the adhesion in both HIV-infected Jurkat cells and mock-infected Jurkat cells to a similar degree (Fig. 1E).
We previously showed that phorbol ester-stimulated adhesion of Jurkat cells to fibronectin required activation of the small GTPase Rap1. 24 To determine whether HIV-infected lymphocytes required Rap1 or Rho1 for spontaneous or phorbol-stimulated adhesion, we treated cells with either GGTI, which inhibits geranylgeranylation of Rap1, or FTI (farnesyltransferase inhibitor), which blocks Rho activity. As expected, roscovitine decreased spontaneous adhesion to fibronectin (Fig. 2A). We found that phorbol myristate acetate (PMA)-stimulated adhesion of HIV-infected lymphocytes and of mock-infected lymphocytes was similarly inhibited by GGTI (Fig. 2B), consistent with a requirement for Rap1 activity in phorbol-stimulated adhesion. In contrast, FTI did not affect PMA-stimulated adhesion in HIV-infected or mock-infected lymphocytes (Fig. 2B). Neither GGTI nor FTI reduced spontaneous adhesion to fibronectin (Fig. 2A), consistent with ligand-induced adhesion being independent of Rap1. As further confirmation that ligand-induced adhesion was independent of Rap1, we demonstrated that transfection of Jurkat cells with a dominant negative construct of Rap1 (N17Rap1) 24 did not inhibit ligand-induced adhesion, but did inhibit phorbol ester-stimulated adhesion, confirming our previous work 1,24 (Fig. 2D). Consistent with a lack of effect of FTI treatment, the RhoA kinase inhibitor Y7632 did not inhibit either ligand-induced adhesion or phorbol ester-stimulated adhesion. In concert, these data suggest that RhoA kinase and Rap1 activity are not required for ligand-induced adhesion.
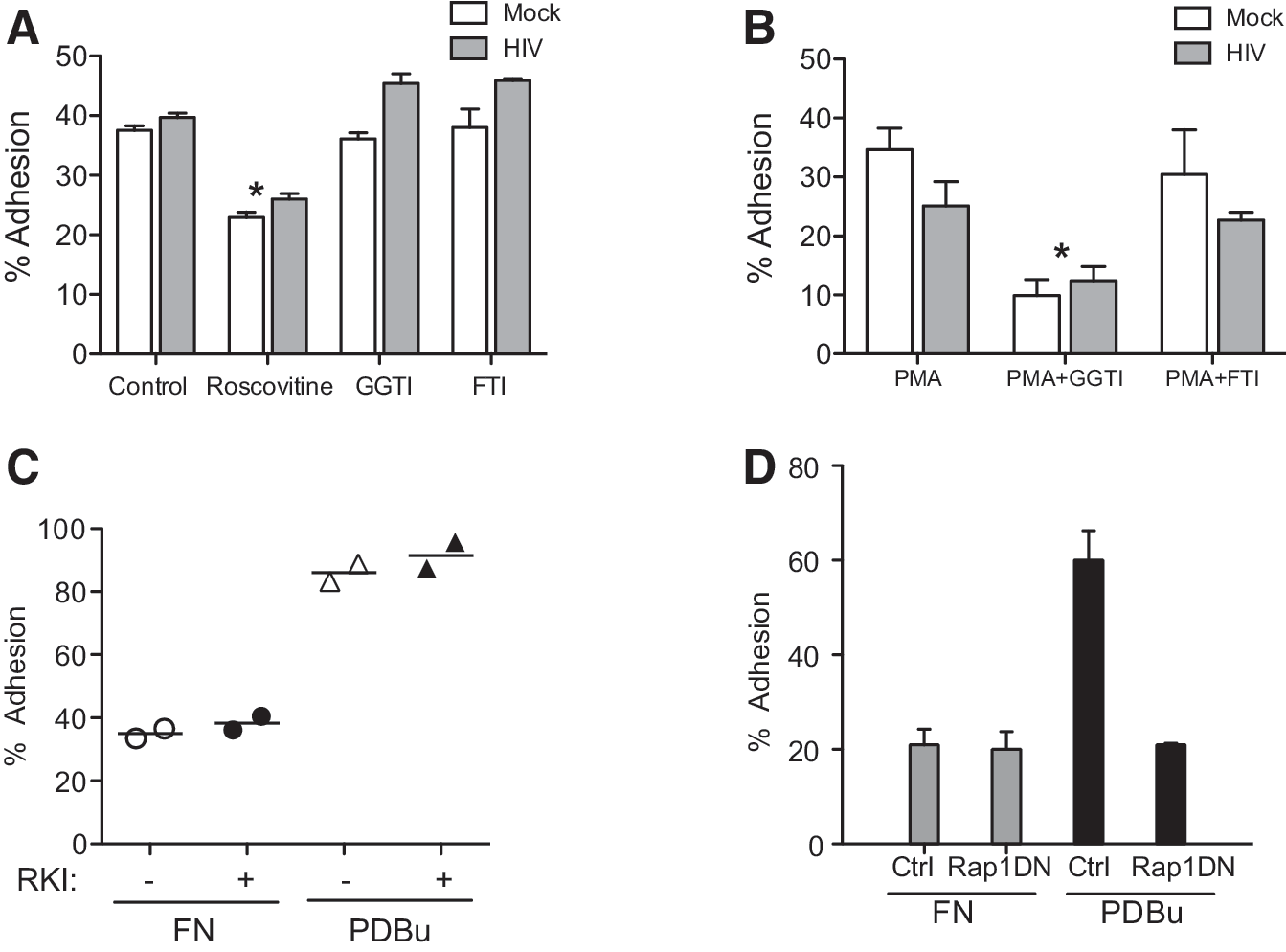
Phorbol myristate acetate (PMA)-stimulated adhesion, but not ligand-induced adhesion, of HIV-infected lymphocytes is Rap1 dependent.
In summary, we expand our previous studies to demonstrate that HIV-infected lymphocytes exhibit similar spontaneous adhesion to fibronectin and to HUVEC matrix as mock-infected lymphocytes, demonstrating that HIV-infected lymphocytes participate in ligand-induced adhesion. Using pharmacological inhibitors of Cdk and dominant negative construct of Cdk4, we show that Cdk4 is involved in ligand-induced adhesion of HIV-infected lymphocytes. In addition, we show that HIV-infected lymphocytes use Rap-1 during phorbol-stimulated adhesion. We conclude that HIV infection does not alter utilization of Cdk4-mediated or of Rap1-mediated adhesion pathways. Furthermore, use of Cdk inhibitors as therapeutic for HIV infection should take into account the potential effects on lymphocyte adhesion and transmigration.
Footnotes
Acknowledgments
This work was supported by NIH HL18645 (J.M.H.), NIH HL HL083481, K24 HL068796, and American Lung Association Career Investigator Award (L.M.S.).
Author Disclosure Statement
No competing financial interests exist.