
Abstract
The APOBEC3 proteins are cytidine deaminases that can introduce G→A mutations in the HIV-1 plus DNA strand. This editing process may inhibit virus replication through lethal mutagenesis (hypermutation), but could also contribute to viral diversification leading to the emergence of escape forms. The HIV-1 Vif protein has the capacity to counteract APOBEC3 factors by recruiting a CUL5-based ubiquitin ligase complex that determines their proteasomal degradation. In this work, we analyzed the APOBEC3-mediated editing in proviral HIV-1 from perinatally infected children (n=93) in order to explore its association with polymorphisms of APOBEC3G and CUL5 genes (APOBEC3G H186R, APOBEC3G C40693T, and CUL5 SNP6), the Vif protein variability, and also the time to AIDS development. To calculate the level of editing, we have developed an index exploiting the properties of a region within the HIV-1 pol gene that includes the central polypurine tract (cPPT). We detected a reduced editing associated with the CUL5 SNP6 minor allele and also with certain Vif variants (mutations at sites 46, 122, and 160), although we found no evidence supporting an impact of APOBEC3 activity on disease progression. Thus, our findings suggest that APOBEC3-mediated editing of HIV-1 could be modulated by host and virus genetic characteristics in the context of pediatric infection.
Introduction
T
Virus and host genetic variability affecting the Vif-APOBEC3 axis has been widely analyzed in terms of its relevance to disease progression. In that sense, it has been reported that the single nucleotide polymorphisms (SNPs) APOBEC3G H186R (rs8177832), APOBEC3G C40693T (rs17496018), and CUL5 SNP6 (rs11212495) might affect viral transmission or progression to AIDS, although discrepancies between different cohorts have been observed. 10 –15 The nonsynonymous polymorphism APOBEC3G H186R (an A→G change in exon 4) has been demonstrated to alter the enzyme processivity, 16 whereas APOBEC3G C40693T (a change in intron 4) is located close to the exon 4 natural 3′ splice site and could potentially affect mRNA splicing by creating an alternative acceptor splice site. 12 The CUL5 SNP6 (an A→G change in intron 3) was found to modify the binding affinity of DNA to nuclear proteins from T lymphocytes, suggesting that this SNP may affect the gene regulation or interaction. 13 In addition, a number of vif alterations (including insertions, deletions, and different substitutions) have been proposed to influence the course of HIV-1 infection. 15,17 –22 Importantly, comprehensive works in diverse groups of HIV-1-infected individuals have attempted to connect the level of editing with APOBEC3 and/or vif genetic variants, but they mainly rendered weak and no conclusive associations. 3,20,23 –25
Up to now, the significance of the Vif-APOBEC3 interplay and of the proper APOBEC3-mediated editing has not been extensively studied in pediatric settings. 15,26,27 In our cohort of perinatally HIV-1-infected children, we have previously observed that Vif alterations (an insertion at position 61 and substitutions A62D/N/S and Q136P) may contribute to a rapid AIDS onset and also that APOBEC3G and CUL5 polymorphisms might select for certain Vif variants. 15 However, we found no case exhibiting a hypermutated proviral population by analyzing vif sequences.
The present work is an extension to our recent study and it is intended to further analyze the APOBEC3-mediated editing in HIV-1 from pediatric patients, exploring its association with both host and virus genetic characteristics, but also with the rate of progression to AIDS. To this end, we have developed an editing index exploiting the properties of a region within the HIV-1 pol gene that includes the central polypurine tract (cPPT).
Materials and Methods
Study patients
The study comprised 93 perinatally HIV-1-infected patients from an Argentinian pediatric cohort of white-Hispanic ethnicity. These children were born within the period 1994–1999 and were selected based on the availability of peripheral blood mononuclear cells (PBMCs) collected during the first year of life (0.2 to 11.9 months, median of 5.5 months). All patients received medical care at the Hospital de Pediatría “Juan P. Garrahan” (Buenos Aires, Argentina). HIV-1 infection status and AIDS definition were established according to the 1994 criteria of the U.S. Centers for Disease Control and Prevention (CDC) classification for children. 28 Of the corresponding 93 mother–infant pairs, 67% received no prophylactic treatment, 30% received zidovudine prophylactic treatment (PACTG 076), 29 and there were no available data for 3%. The study period ended in May 2006 and the median follow-up time was 85.8 months (2.7–141.1 months). During follow-up, 74% of patients received HAART and 72% progressed to AIDS. The analysis of time to AIDS was performed according to recorded clinical data. The Ethics Committee and the Institutional Review Board of the hospital approved the study. Written informed consent was obtained from the parents or legal guardians of the children.
Genotyping of APOBEC3G and CUL5 polymorphisms
Genotypes for the SNPs APOBEC3G H186R, APOBEC3G C40693T, and CUL5 SNP6 were determined by PCR-restriction fragment length polymorphism (RFLP) assays using the enzymes HhaI, BseRI, and XceI, respectively. Assays were developed based on the reports by An et al., 10,13 as previously described. 15
HIV-1 gene amplification, sequencing, and analysis
Proviral genes were amplified from PBMC samples obtained during the first year of life of the 93 children studied. This was intended to minimize the possible effects of confounding factors, such as the influence of time since infection and/or therapy history on proviral population characteristics. Amplification of a 312-bp pol segment (HXB2 positions 4584–4895) surrounding the cPPT was performed by nested-PCR, using the degenerate primers (5′-3′) HypIN-F1 AARTTAGCARRAARATRGCCAG/HypIN-R1 TTTGCTGGTYYTTTYYAAASTGG for the first round and HypIN-F2 AACAATACATACARACAATRGC/HypIN-R2 CTGTYYYTGTAATAAACYYGRAA for the second round. Both pairs of degenerate primers were designed to efficiently recognize edited and nonedited target sequences, similarly to Janini et al. and Suspène et al.
30,31
The PCR products were purified with the QIAquick purification columns (QIAGEN, Germany) and then subjected to bulk sequencing using both second round primers. Sequencing reactions were conducted with the Big Dye Terminator v1.1 sequencing kit (Amersham Biosciences, England), run on an ABI PRISM 3130 automated sequencer and analyzed with the DNA Sequencing Analysis Software v5.3.1 (Applied Biosystems, USA). Chromatograms were visually inspected and edited using FinchTV v1.4.0 (
Detection of APOBEC3-mediated editing
To estimate the APOBEC3-mediated editing, the sequences of the 312-bp pol segment were analyzed. The Hypermut 2.0 software (
Statistical analysis
Differences in the level of editing between HIV-1 subtypes, APOBEC3G/CUL5 genotypes or, Vif variants were analyzed with the Wilcoxon rank sum test. The normal distribution of the ODDSMM index was evaluated with the Shapiro–Wilk normality test. The association of editing and time to AIDS was analyzed with a likelihood ratio test under a Cox proportional hazards regression model. The association among different Vif mutations and the association of genetic distance with ODDSMM were determined with the Spearman rank correlation test. All tests were conducted using Statistix v7.0.
Results
Analysis of APOBEC3-mediated editing
Sequences of the cPPT region of proviral HIV-1 were obtained by bulk sequencing of early samples from 93 perinatally infected children. The 312-bp amplified segment contains both a maximum and a minimum for APOBEC3 susceptibility and thus could be considered highly informative for the study of editing. It was observed that 72.0% (67/93) of sequences were subtype F1, 26.9% (25/93) were subtype B, and 1.1% (1/93) were subtype A1. To identify highly edited sequences, the Hypermut 2.0 software was employed. Sequences were separately analyzed by using appropriate references according to their viral subtype. This procedure indicated no sequence as hypermutated (p>0.05 for general GR or specific GG/GA contexts), which pointed out the rarity of pediatric cases showing a major fraction of proviral population composed of severely edited forms.
It should be remarked that Hypermut 2.0 is a broadly employed tool fundamentally applied to classify sequences in the categories of “hypermutant” or “nonhypermutant,” and that the same “all or nothing” criterion is usually adopted by other approaches based on editing scores and cluster analysis. 3,24 However, strategies that do not rely on such distinction have proven valuable for association studies, 4,5,20,25 stressing that the levels of editing may be informative through a wide range before the affected sequence can be considered as hypermutated. Thus, we decided to carry out a careful study focused in the spectrum of low APOBEC3-mediated editing potentially observed in our pediatric cohort. To this end, we especially developed the ODDSMM index, attempting to improve the analysis of the editing phenomenon when it occurred at mild levels. In this index, the editing signal is estimated as the A:G ratio within the APOBEC3 contexts in relation to the A:G ratio outside of them, which is detailed in the expression [(AG+AA):(GG+GA)]/[(AC+AT):(GC+GT)]. To obtain the ODDSMM value, the editing signal is calculated for a nucleotide sequence placed in maximum susceptibility to APOBEC3 activity (immediately upstream the cPPT) and then adjusted by the signal in a contiguous minimum (the proper cPPT and the immediately downstream sequence). This estimation was intended to increase the specificity for editing measurement, correcting the signal of editing attributable to APOBEC3 activity by a signal that could be considered as basal or spurious.
The ODDSMM values were calculated for the 93 study cases and it was observed that they significantly differed between HIV-1 subtypes (p<0.0001, Fig. 1a). Consequently, we decided to restrict the following analysis to the most abundant subtype F1 cPPT regions. In addition, to minimize further confounding related to viral lineage (given the presence of different B/F1 recombinants), we considered only those cases also assessed as subtype F1 for the previously sequenced vif genes. Hence, we continued our studies of editing with a subset of 66 pediatric patients. This group presented an ODDSMM distribution that did not significantly deviate from normality (p=0.3121, Fig. 1b and c).
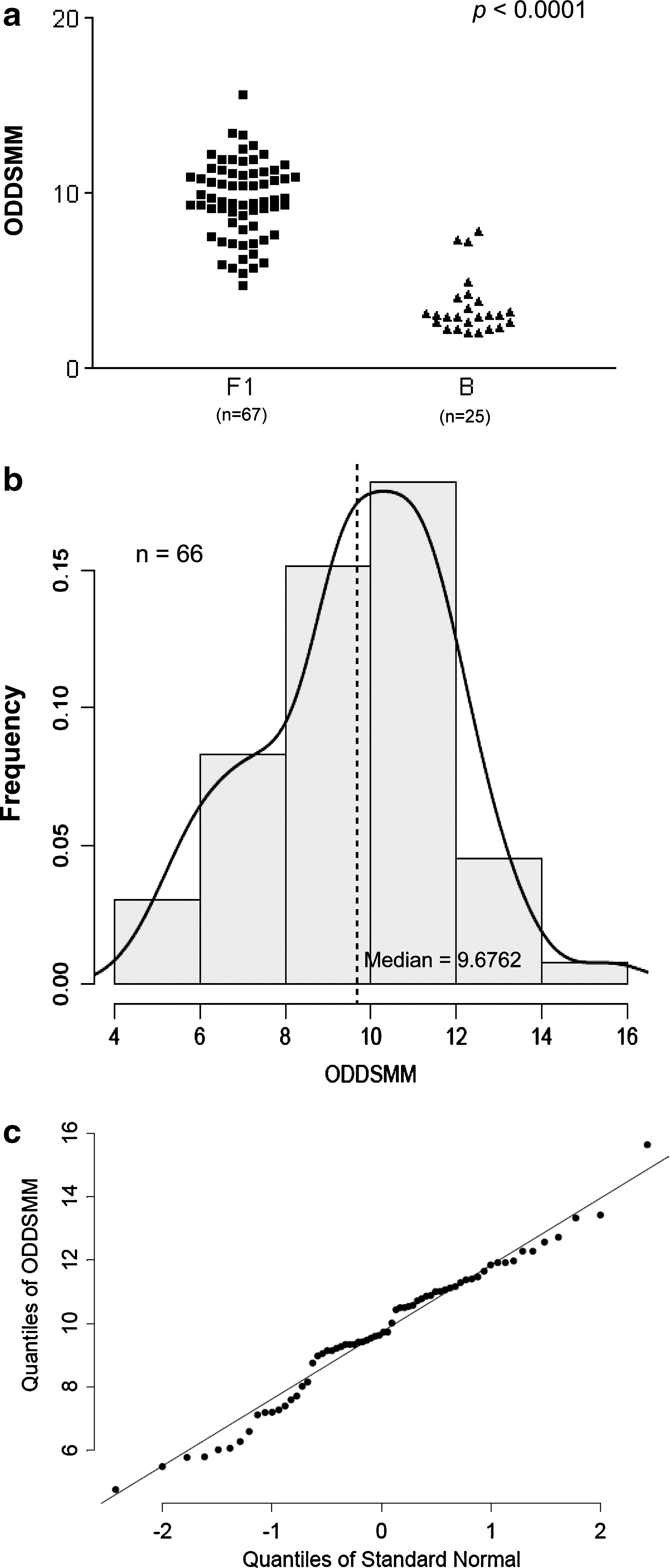
APOBEC3-mediated editing and ODDSMM. Statistically significant differences of ODDSMM were observed between HIV-1 subtype F1 (mean=9.69, SD=2.19) and B (mean=3.57, SD=1.65) (Wilcoxon rank sum test, p<0.0001)
APOBEC3-mediated editing and APOBEC3G/CUL5 polymorphisms
To determine whether different levels of editing were associated with the polymorphisms APOBEC3G H186R, APOBEC3G C40693T, or CUL5 SNP6, we compared the ODDSMM values between major allele homozygotes and minor allele carriers for each of these loci. While the APOBEC3G SNPs were not related to differences of ODDSMM (p>0.05, Fig. 2a and b), the CUL5 SNP6 minor allele was associated with a reduced editing (p=0.0479, Fig. 2c).
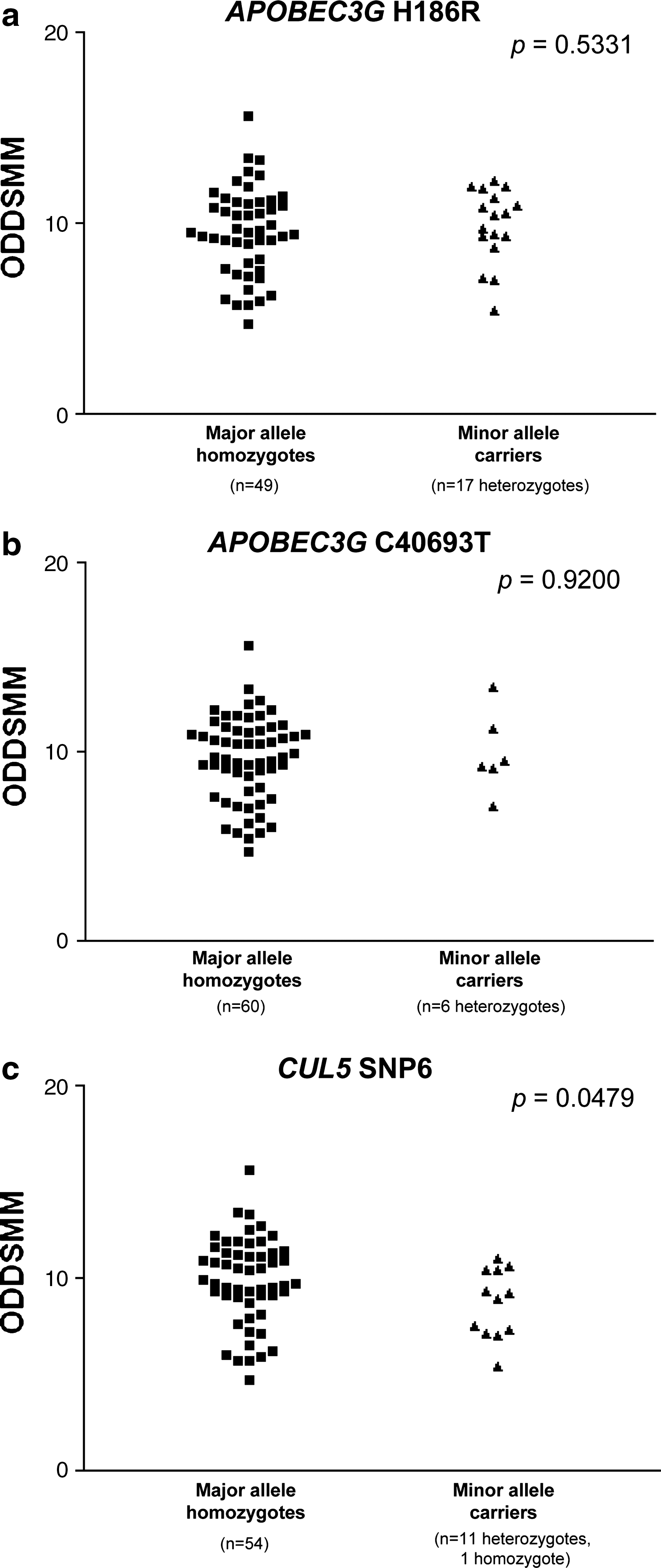
Association of editing with APOBEC3G/CUL5 polymorphisms. No differences in the level of editing (ODDSMM) were observed between genotypes of APOBEC3G H186R (major allele homozygotes mean=9.63, SD=2.30; minor allele carriers mean=9.93, SD=1.93) and APOBEC3G C40693T (major allele homozygotes mean=9.68, SD=2.22; minor allele carriers mean=10.00, SD=2.12) (Wilcoxon rank sum test, p=0.5331 and p=0.9200, respectively)
APOBEC3-mediated editing and Vif variability
We next evaluated whether differences in the level of editing were associated with differences in Vif variability. The cases studied were divided according to their ODDSMM into two groups showing values below or above the median, to subsequently compare the entropy at each Vif position between them. The analysis indicated that increased entropy at sites 46, 122, and 160 was linked to lower ODDSMM, while increased entropy at site 182 was linked to higher ODDSMM (p<0.05, Fig. 3a). Then, the editing levels exhibited by wild-type Vif were directly contrasted to those showed by mutants for the mentioned positions. Although scatterplots revealed overlapping values, statistically significant differences were found between Vif variants at sites 46, 122, and 160 (p<0.05, Fig. 3b–e). The simultaneous occurrence of these mutations was evaluated but no association was found (p>0.05).

Association of editing with Vif variants. Vif positions 46, 122, 160, and 182 showed entropy differences related to the level of editing. Statistical confidence (10,000 randomizations with replacement) is indicated
APOBEC3-mediated editing and progression to pediatric AIDS
To assess the relevance of editing for the rate of disease progression in our pediatric cohort, we evaluated whether variations of ODDSMM were linked with changes in time to AIDS. The analysis showed no evidence for an association in the 66 cases considered (p=0.9884). In addition, to discard therapy as a confounding variable, we conducted an analysis censoring patients at time of HAART initiation, but the association remained not significant (p=0.8199). Thus, the levels of editing observed seemed to have no effect on the risk of pediatric AIDS development.
APOBEC3-mediated editing and viral phylogenetic divergence
To explore whether editing affects HIV-1 evolutionary processes, phylogenetic trees were constructed with the sequences obtained for the pol segment and for the complete vif gene. We noticed that for the two regions there was an inverse correspondence between branch length and level of editing (Fig. 4a and b). Genetic distances of each pol or vif sequence to the respective consensus (generated for the 66 subtype F1 cases) showed a negative correlation with ODDSMM values for both viral genomic regions (p<0.05, Fig. 4c and d).
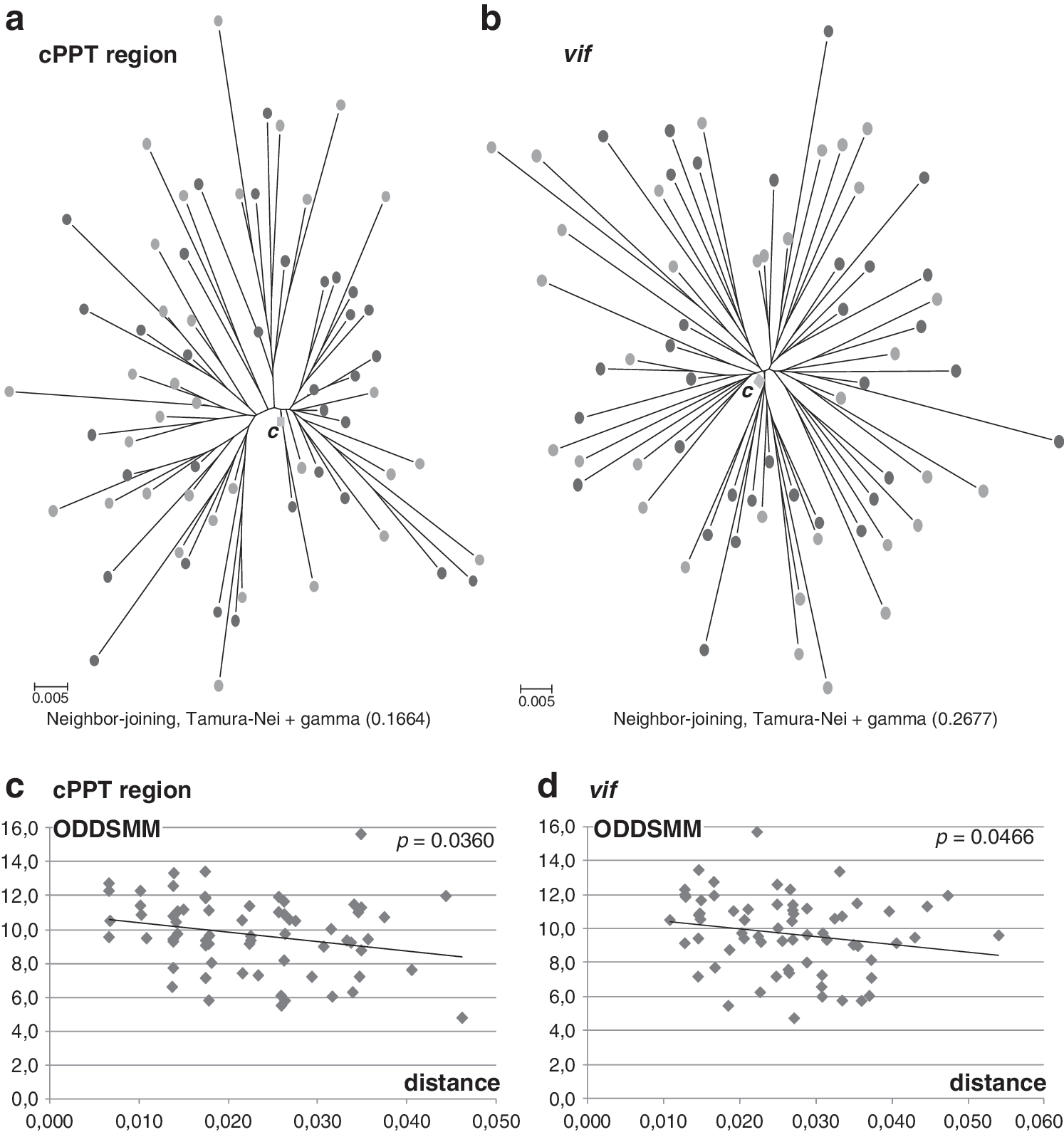
Editing and viral divergence. Phylogenetic trees for the cPPT region and the complete vif gene exhibited an inverse correspondence between branch length and the level of editing (light circles: ODDSMM<median, dark circles: ODDSMM>median,
Discussion
In this study, we analyzed the APOBEC3-mediated editing affecting HIV-1 from pediatric patients and its association with host genetic polymorphisms, virus variability, and disease progression. Despite the fact that we studied a pol segment reported as highly susceptible to APOBEC3 enzymatic activity, we detected no case exhibiting a severely edited or hypermutated proviral population. This is in sharp contrast with those works detecting hypermutation by direct sequencing in a considerable fraction of infected adults. 3 –5,24 Thus, it should be noted that we explored the editing phenomenon in a range limited to levels that could be considered as mild or sublethal. 37
Differences in the level of editing were not observed for APOBEC3G genetic variants, although an effect of CUL5 SNP6 could have been identified. The minor allele at this locus was associated with a lower editing, which would be in accordance with an incremented gene activity favoring viral replication as discussed by An et al. 13 An increase in CUL5 protein availability could contribute to Vif depletion of APOBEC3 factors, which might be reflected in a reduction of HIV-1 editing.
We also observed that different levels of editing were linked to changes in Vif variability. Despite the fact that the affected sites have not been reported as central for Vif function, it is interesting to note that they lie within or in close proximity to relevant protein regions. Site 46 is contiguous to an APOBEC3G-binding domain (amino acids 40 to 45). 38,39 Residue at position 122 is immersed in the HCCH motif, required for Vif recruitment of CUL5. 40,41 Site 160 is immediately before the proline-rich motif (positions 161 to 164), which has been reported as important for Vif interaction with APOBEC3G, CUL5, and ElonginB, but also for Vif oligomerization. 42 –45 Position 182 seems to be an exception, since it is located in the C-terminal tail that is usually considered as dispensable for Vif functionality. 46 Thus, it could be hypothesized that substitutions at these sites modify Vif activity at least in a subtle manner, but enough to systematically alter the level of editing.
We found no evidence supporting an association of editing (in the mild levels observed) with time to AIDS when looking for a monotonous relation between these variables in vertically infected children. In fact, APOBEC3 restriction of HIV-1 by means of hypermutation might be considered as a negligible cause for a delayed AIDS outcome in our pediatric cohort, since no patient exhibited signs of hypermutation substantially affecting the proviral population. Our results are in agreement with those recently published by Amoedo et al., 27 who found a low percentage of hypermutated clones without a correlation between its abundance and the profile of disease progression in perinatally infected children.
We have previously reported that the analyzed APOBEC3G and CUL5 polymorphisms showed no effect on time to AIDS in our pediatric cohort, although some Vif variants (importantly, other than those linked to editing differences in the present work) could be associated with an accelerated AIDS outcome. 15 Hence, in a general consideration, the variability at the Vif –APOBEC3 axis may affect both the level of editing in HIV-1 proviral population and the course of disease in children, but it seems that it is not in a clearly coupled manner.
Additionally, while analyzing the influence of APOBEC3 enzymatic activity on HIV-1 evolution, we noticed that phylogenetic divergences estimated for the obtained proviral sequences were negatively correlated with editing. It is difficult to determine a clear interpretation of these results, since our ODDSMM index (similar to any other editing measurement) is to some extent related to genetic distance. However, this association was observed coincidentally for both the cPPT region and the vif gene (not used for editing calculation), leading us to speculate about an actual biological meaning. In that sense, it may be proposed that editing, even at a sublethal range, could negatively affect the replicative dynamic of virus with a consequential impact on its evolutionary rate, as seems to be reflected at the level of between-host evolution studied.
In conclusion, our exploratory findings suggest that the APOBEC3-mediated editing of HIV-1 could be modulated by both host and viral genetic characteristics in the context of pediatric disease. We consider that these results will be relevant for the design of future studies aimed to determine the role of the Vif–APOBEC3 axis in the course of infection.
Sequence Data
Sequences analyzed in this work have been submitted to GenBank. Accession numbers for pol: JN630631–JN630723. Accession numbers for vif: JF494923–JF495015.
Footnotes
Acknowledgments
We thank Ms. Natalia Beltramone, Ms. Bárbara Campos, Mr. Rodrigo Cánepa, and Mrs. Carmen Gálvez for technical assistance and Mrs. Silvia Marino for her technical and scientific contribution to this work. The study was supported by Fondo Nacional para Ciencia y Tecnología (FONCYT, PICT 25830) and Consejo Nacional de Investigación Científica y Tecnológica (CONICET, PIP 6057).
Author Disclosure Statement
No competing financial interests exist.