
Abstract
A new syncytia-inducing human immunodeficiency virus type 1 (HIV-1) strain designated PokA79 was first isolated from a peripheral blood mononuclear cells (PBMC) pool from 24 HIV-1-infected patients from Russia and adapted to growth in MT2, MT4, and U937 cell cultures. The full-length genome of this isolate was amplified by RT-two-round PCR and sequenced. The obtained sequence belongs to the A1 subtype and clustered with other A1 former Soviet Union (FSU) strains. Full-length genome analysis showed that some of the PokA79 genes contain insertions and frame shifts, and the gp120 V3 loop has the three amino acid insertion inside its apex, which is rare for subtype A1 sequences. Nonidentical insertions of the same size are also present in the V3 apex region of several HIV-1 sequences from Central Africa.
The HIV/AIDS epidemic in Russia, Ukraine, and other former Soviet Union territories has achieved an infectivity rate of over 1% of the total population, with over 90% of all HIV-infected people in Eastern Europe represented in Russia and Ukraine. 1 The development of an HIV-1 vaccine remains an unsolved problem and is considered to be crucial for the nation's survival under these circumstances. At present, clinical trials for several anti-HIV vaccines containing fragments of viral DNA and/or HIV-1 proteins are being carried out in different countries. Commonly, laboratory HIV-1 strains are widely used for HIV vaccine design, and the majority of these strains such as HTLV-IIIB/H9, RF, and SF2 belong to subtype B. At the same time, taking into account the striking genetic variability of HIV-1, the use of native isolates and laboratory HIV-1 strains more similar to viral variants widespread in affected regions and cohorts of patients seems to be more useful for effective region-specific vaccine development.
At present HIV-1 subtype A1 strains predominate in the territory of the Russian Federation and circulate mainly among intravenous drug users (IDUs) and their sexual partners, 2 unlike the situation in North America, Western Europe, and Australia where subtype B predominates. Moreover, the HIV epidemic in Russia was initially caused by a variant of monophyletic origin often referred as IDU-A, which was introduced from southern Ukraine, and later spread widely throughout the territory of Russia, subsequently forming different subvariants. 3 –6 However, no laboratory subtype A1 HIV strain capable of propagating infection in vitro that is genetically similar to region-specific variants and suitable for HIV biology research has yet been obtained in Russia. Therefore, laboratory subtype B HIV-1 strains and several subtype A strains of African origin such as U455, along with primary isolates, have been used for HIV-1 vaccine research in Russia. Thus, obtaining such a domestic laboratory HIV-1 subtype A strain from native Russian viral isolates is an actual problem.
Currently, according to the literature, new laboratory HIV-1 strains capable of replicating in vitro were routinely isolated from biomaterial obtained from a single infected person. In this work in order to increase the probability of a positive outcome we used another approach based on pooled peripheral blood mononuclear cells (PBMCs) from HIV-1-infected patients for virus strain isolation. This article describes the isolation of a new subtype A1 strain from the PBMC pool of 24 HIV-1-infected patients, analysis of its full-length genome sequence, in vitro multiplication dynamics, and other biological characteristics.
Blood samples were obtained in September 2008 from 24 HIV-1-positive patients of a single cohort from Russia. All individuals were IDUs and all were infected with HIV-1 subtype A1 viruses. PBMCs from these patients were isolated by a standard Ficoll gradient total PBMC blood fractionation method. Heparinized blood was diluted with an equal volume of RPMI-1640 medium. Diluted donor blood was stratified slowly on a Ficoll-Histopaque (1.078 g/sm3) and centrifuged for 20 min at 2300 rpm. Gradient PBMC rings were carefully transferred to sterile tubes, diluted with an equal volume of RPMI-1640, and centrifuged for 15 min at 1500 rpm. Isolated PMBC pellets were resuspended in 30 ml of RPMI-1640 supplemented with 10% fetal bovine serum (FBS; HyClone, USA) and 5 μg/ml phytohemagglutinin (PHA) and placed in a culture flask at a density of 1–2×106 cells/ml. Isolates were incubated at 37°C in 5% CO2 for 24 h, then cells were transferred to RPMI-1640 medium supplemented with 10% FBS and 10 units/ml of interleukin-2 (IL-2). Ficoll-separated PBMCs from HIV-1-infected patients were mixed together in one pool.
The virus was isolated by cultivation of the PBMC mixture together with PHA-stimulated healthy donor PBMCs in RPMI-1640 medium supplemented with 10% FBS and 10 units/ml of IL-2. Next, 10 ml of the culture was placed in a 25-cm2 culture flask and maintained for 24 days. Cultures were monitored for syncytia formation and cytonecrosis once or twice per week, and fresh donor PBMCs (1 ml) were added to the culture every 1–2 weeks if necessary. Virus replication was monitored by p24 production in culture supernatants using a p24-specific immune-enzyme assay. For this purpose samples of culture supernatant were taken every 3–4 days, and isolates were supplemented with 4–5 ml of fresh growth medium. p24 concentration was measured by an indirect immune-enzyme assay according to a protocol developed in our laboratory using monoclonal mouse IgG antibodies to p24 protein and polyclonal biotin-conjugated goat antimouse antibodies. The sensitivity of this test system is 160 pg/ml of HIV-1 p24 antigen.
A titrated solution of recombinant p24 with an initial concentration of 10 ng/ml was used as the positive control. Culture supernatants of uninfected MT-2, MT-4, and donor PBMCs were used as negative controls. Briefly, 150 μl of virus containing culture supernatant was added to each well of the first 96-well plate (plate #1), 20 μl Tween-80 (1%) was added, and the plate was incubated for 1.5 h at 37°C. Next, 100 μl of monoclonal murine IgG antibodies recognizing p24 (4 μg/ml in PBS) was added to each well of a second 96-well plate (plate #2), and the plate was incubated for 1 h at 37°C. Plate #2 was emptied and washed four times with PBS buffer (250 μl per well). Then, 100 μl of PBS/casein (1%) was added to each well of plate #2, and incubated 1 h at 37°C. Plate #2 was emptied and washed four times with PBS-Tween (0.2% v/v Tween-20 in PBS), 250 μl per well. The samples (100 μl) were then transferred from plate #1 to plate #2. Recombinant p24 (10 ng/ml) was added to the control wells, and the plate was incubated for 1.5 h at 37°C. The plate was emptied and washed four times with PBS-Tween buffer (250 μl per well). Then, 100 μl of polyclonal biotin-conjugated goat antimouse antibodies in PBS/casein (1:2000) was added to each well, and the plate was incubated 1 h at 37°C. The plate was emptied and washed four times with PBS-Tween (250 μl per well) and 100 μl of the conjugate (6 μl of HRP-streptavidin in 12 ml of PBS) was added per well and the plate was incubated for 1 h at 37°C. The plate was emptied and washed five times with PBS-Tween buffer (250 μl per well). Next, 100 μl of TMB was added per well and the plate was incubated for 10–15 min at room temperature. The reaction was stopped by adding 100 μl of 1 M H2SO4 per well. The obtained results were analyzed spectrophotometrically. The quantity of p24 was estimated in P/N units, where P is the optical density of the antigen-containing sample and N is the optical density of the negative control.
Virus-containing supernatant was then used for infecting of MT-2 and MT-4 cells. Once virus production was detected in infected PBMC supernatants by measuring the p24 antigen levels, the virus was transferred to 4-day-old MT-2 and MT-4 cultures (human T cell leukemia, NIH AIDS Research and Reference Reagent Program, USA) with ≥85% viability (final cell concentration 1.25×105 cells/ml), and cells were cocultivated for 3 days in RPMI-1640 supplemented with 2% FBS at 37°C in 5% CO2. Virus passaging was performed after the appearance of visible signs of viral cytopathic action. After the first two passages a new virus strain was provided every 2–3 days with fresh uninfected MT-2 or MT-4 cells grown in RPMI-1640 supplemented with 10% FBS; 2% FBS was used for cocultivation. The resulting stably infected cell lines were further used to obtain total viral RNA preparations. HTLV-IIIB/H9 HIV-1 subtype B (NIH) and U455 HIV-1 subtype A (NIH) laboratory strains cultivated under the same conditions were used as positive controls.
The new viral isolate was an HIV-1 strain, confirmed by standard methods of microscopy, an immune-enzyme assay, and PCR. Infected MT-2 and MT-4 cells were analyzed by microscopic study in order to find cell syncytia that are specific signs of HIV infection. Simultaneously U455 and HTLV-IIIB/H9 HIV-1 strains were passaged on MT-2 cells and used as positive controls, and uninfected MT-2 and MT-4 cells were used as negative controls. The monitoring of MT-2 cells infected with the new viral isolate demonstrated large diffusely spread syncytia after 2–3 days of incubation (Fig. 1). The syncytia formation dynamics of the HIV-1-positive control variants were similar to those of the new isolate. PCR amplification from the infected cell genomic DNA preparations and sequencing of four short viral genome regions corresponding to HIV-1 gag [G10 (HXB2 942–1307) and G13 (HXB2 1758–2018) regions], pol [P4 (HXB2 2548–3256) region], and env [E14 (HXB2 6998–7318) region] also confirmed that our new viral isolate was an HIV-1 strain.
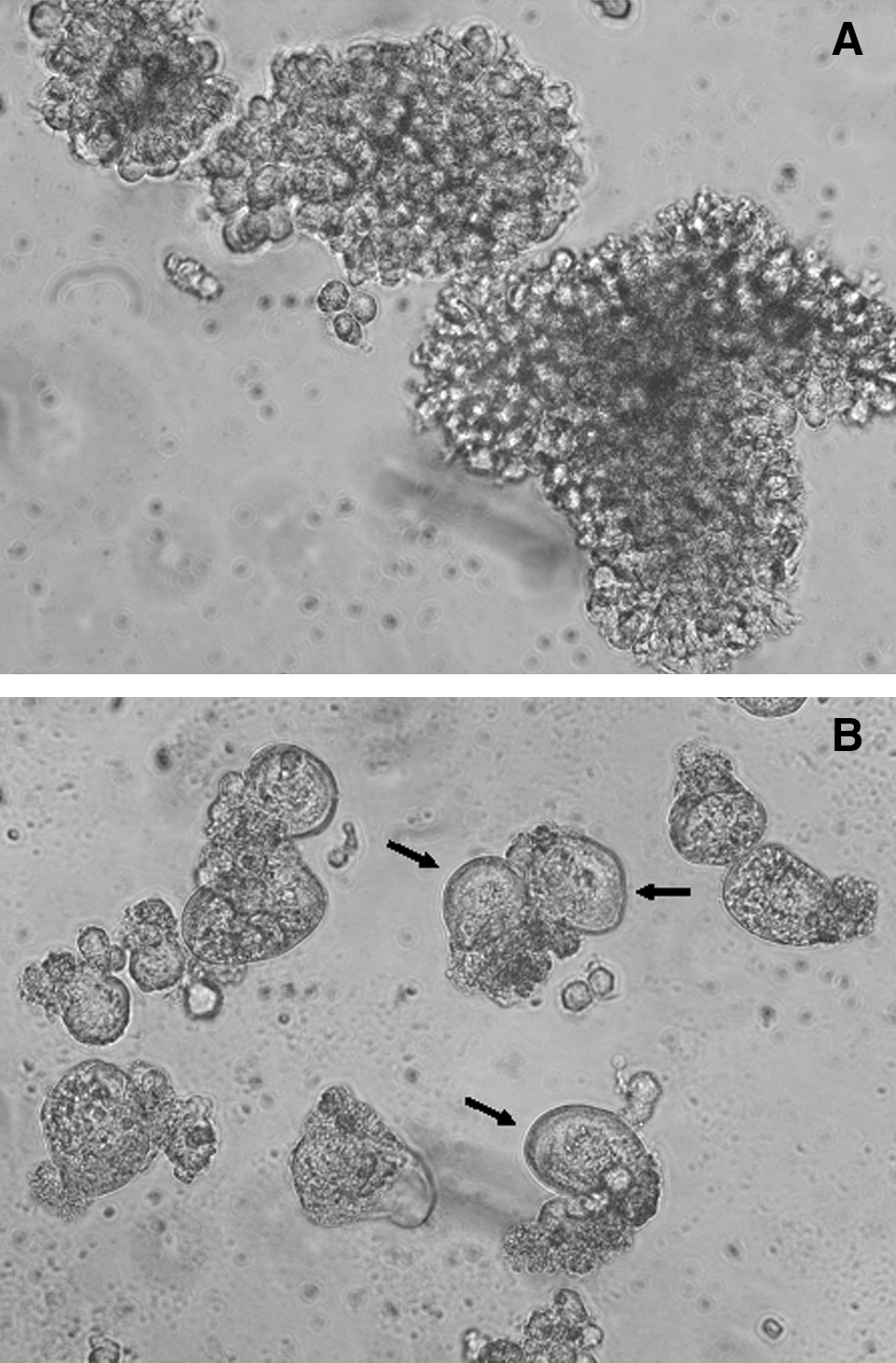
Uninfected MT-2 cells
The analysis of the obtained short sequences allowed us to preliminarily classify our virus as the A1 subtype. The new viral HIV-1 strain was named PokA79 and was subjected to detailed phenotypic characterization. A number of available cell cultures were used for the study of virus phenotropic infectivity. We determined the spectrum of PokA79 sensitive and optimal for virus propagation cell lines using our laboratory panel consisting of HIV-1-sensitive lymphocyte, macrophage, and CD4 receptor expressing cell lines and several resistant to HIV-1 infection cell lines. Uninfected cells of the same cell lines were used as negative controls. PM-1 cells (human T-lymphoblast lymphoma, NIH) were grown in RPMI with 2 mM glutamine and 10% FBS. HeLa, HeLa-CD4-LTR-beta-Gal, U373-MAGI, U373-MAGI-CCR5E, U373-MAGI-CXCR4, COS-1 (NIH), and COS-7 (DSMZ, Germany) cultures were grown in DMEM (Paneco) supplemented with 2 mM glutamine and 10–12% FBS. The antibiotics hygromycin B, puromycin, and G418 (Sigma) were added when necessary according to standard protocols for cell line maintenance.
Experiments on all selected cell lines were carried out in six-well plates. Cells were placed in the wells and after 24 h of incubation in optimal media with glutamine and 2% FBS they were infected with the new HIV-1 isolate PokA79 or the control HTLV-IIIB/H9 and U455 strains. Cultures were monitored microscopically for syncytia formation. Finally, infected MT-2, MT-4, U-937, HeLaT4, HeLa-CD4-LTR-B-gal, and Raji cells showed clear signs of syncytia formation. At the same time, negative control HeLa, COS-1, and COS-7 cells were not susceptible to infection with PokA79, HTLV-IIIB/H9, or U455 HIV-1 strains. Maximal syncytia formation was detected on PokA79-infected MT-2 cells. An indirect immune-enzyme assay was performed for detection and measurement of the HIV-1 p24 protein concentration in the medium. Supernatant aliquots from infected cell cultures were taken daily for p24 analysis by immune-enzyme analysis on days 0–8 after infection. The maximum quantity of p24 in primary isolates was detected after 10–14 days of cultivation. Analysis of p24 accumulation dynamics showed that the PokA79 strain belongs to the rapid/high type. Finally, the MT-2 and MT-4 suspension cell lines (maximum p24 on day 2–3) and the cell line U-937 (maximum p24 on day 3–4) were selected as being optimal for high, stable PokA79 virus reproduction (Fig. 2).
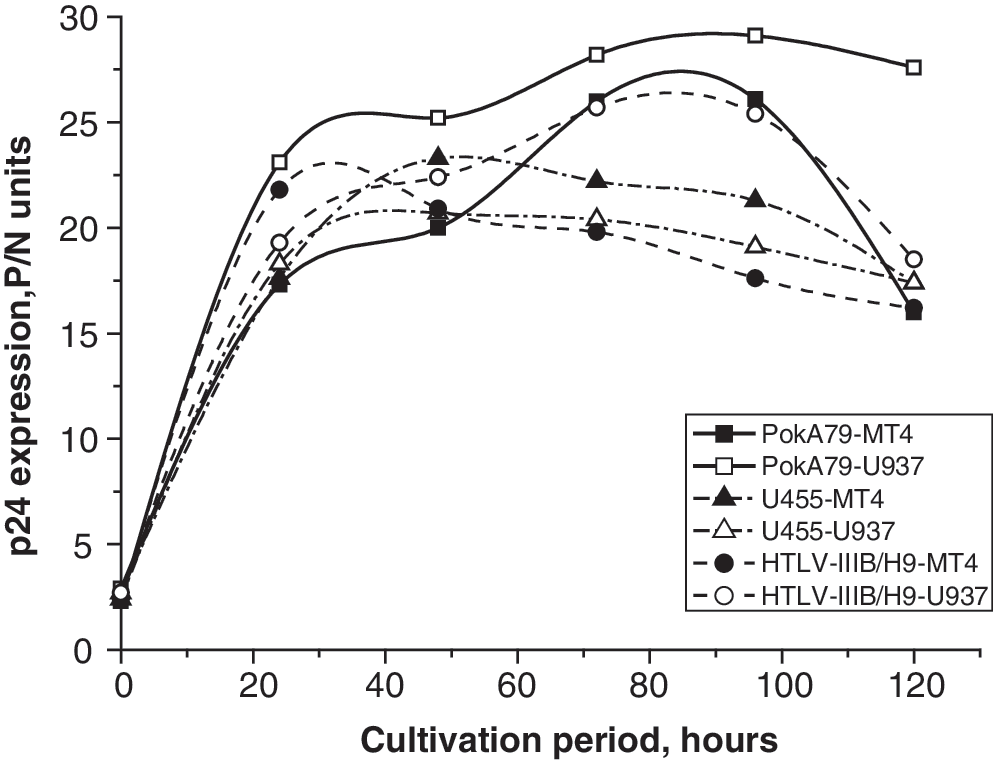
p24 accumulation dynamics for HTLV-IIIB/H9, U455, and PokA79 HIV-1 laboratory strains on MT-4 and U937 cell lines. The p24 quantity is estimated in P/N units, wherein P is the optical density of the antigen-containing sample and N is the optical density of the negative control.
A full-length DNA variant of the PokA79 genome was obtained by RT-two-round nested PCR. Total cellular RNA from MT-2 cells infected with the new HIV-1 isolate was extracted with the SV Total RNA isolation system (Promega). The obtained total RNA preparation was used as the template for cDNA synthesis. Reverse transcription was performed using the RevertAid First Strand cDNA Synthesis kit (Fermentas) with HIV-1 LTR primer MSR5. Thus, long cDNA corresponding to the full-length HIV-1 RNA genome was obtained. Then we performed long-range two-round nested PCR to obtain a sufficient amount of HIV-1 genomic DNA for subsequent sequencing needs. The obtained cDNA was used as the template for primary PCR. The primary PCR product (frA) was obtained using LTR primers E11 (91F) and MSR5 (allowing amplification of nearly the entire HIV-1 genome between positions 516 and 9632 in HXB2) and the special Long PCR Enzyme Mix (Fermentas) allowing the amplification of long DNA matrixes. The obtained long dsDNA was used as the template in the second rounds of PCR amplification for the generation of seven overlapping fragments Fr1–Fr7 representing the entire HIV-1 genome. The second PCR rounds were performed using the High Fidelity PCR Enzyme Mix (Fermentas). The primers used for the amplification of secondary PCR products are listed in Table 1. The obtained Fr1–Fr7 DNA fragments were separated on a 1.7% agarose gel and gel purified with the QIAEX II Gel Extraction Kit (Qiagen). Purified DNA fragments were fully sequenced directly on both strands without cloning using the ABI PRISM BigDye Terminator v 1.1 Cycle Sequencing Kit on an automated ABI Prism 377 DNA sequencer (Applied Biosystems Inc., Foster City, CA). The sequenced Fr1–Fr7 fragments were analyzed and assembled into a contiguous sequence using Vector NTI Advance 9.1 ContigExpress software. Sequencing primers were taken from the literature sources 7 and the NIH catalogue.
Primers Used for Amplification of Secondary Polymerase Chain Reaction Products
PCR, polymerase chain reaction.
The obtained sequence was deposited in the GenBank database under accession number FJ864679 and subjected to detailed sequence analysis. Comparison of the PokA79 genomic sequence with the HIV-1 subtype reference sequences in the SimPlot 2.5 program showed that it belongs to subtype A1 and has no recombination breakpoints. Similar results were obtained in the online RIP 3.0 program where PokA79 was compared to subtype consensus sequences. Multiple alignment of the PokA79 sequence with HIV-1 subtype A, A1, and A2 full genomic DNA sequences from the Los Alamos HIV Sequence Database 8 was generated using the Clustal X2 program. Phylogenetic analysis was performed using MEGA version 4.1 and revealed the similarity of the PokA79 sequence to other HIV-1 A1 full-length sequences from the former USSR presented in the HIV Sequence Database.
The resulting neighbor-joining phylogenetic tree constructed using the neighbor-joining method with the genetic distances calculated using the Kimura two-parameter model and 500 bootstrap replicates is shown on Fig. 3. Bootstrap values equal to or greater than 70% were considered to be significant. The gene structure of the new isolate was normal; the genome sequence contained the full set of intact open reading frames, encoding the Gag, Pol, Env, Vif, Vpr, Vpu, Tat, Rev, and Nef proteins. At the same time, the PokA79 sequence has some interesting unique features.
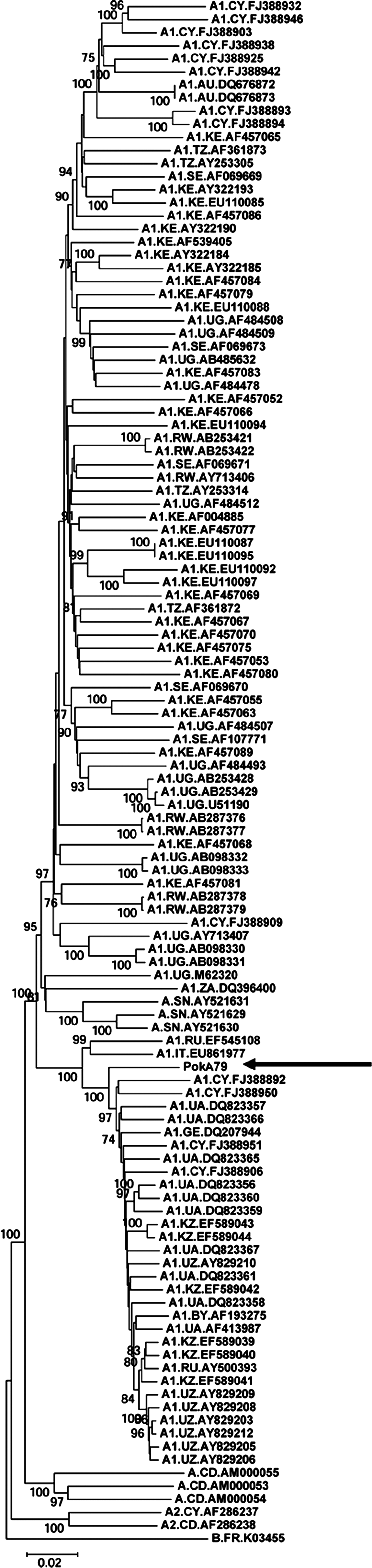
Neighbor-joining phylogenetic tree of full-length HIV-1 subtype A, A1, and A2 genomic sequences. The PokA79 sequence is marked by an arrow. The subtype B sequence K03455 was used as the outgroup. Bootstrap values lower than 70% are not shown.
Specifically, the sequence encoding PokA79 Tat is 15 nucleotides longer than 461 other HIV-1 subtype A tat sequences in the HIV Sequence Database 9,10 due to an A→G substitution in the second position of the TAG stop codon that led to an additional five amino acids extension (WRADS) on the C terminus of Tat (Fig. 4). The other main Tat domains remain unchanged. Tat is a transactivator of HIV gene expression and is essential for productive and processive transcription from the HIV-1 proviral LTR promoter. 9,10 It acts by binding to the TAR RNA element and recruits several cellular proteins to the transcription initiation site leading to hyperphosphorylation of cellular RNA polymerase II and potentiating the synthesize of full-length HIV transcripts from the viral promoter. Tat also acts as a modulator of the expression levels of a number of cellular genes. 10 Moreover, the extracellular form of Tat, which is released from productively infected cells, can also enter uninfected and latently infected cells, inducing apoptosis in the former and activating the transcription of the viral genome in the latter. 10 Taking into account the multiple regulatory roles of the Tat protein, the revealed five amino acid C-terminal extension of this protein in PokA79, which is unique among HIV-1 subtype A, A1, and A2 Tat sequences presented in the HIV Sequence Database, may be important. The PokA79 sequence was also characterized by a 21-bp in frame insertion (seven additional amino acids) in the region encoding the p6 Gag protein; simultaneously, a 21-bp insertion in another reading frame appeared in the overlapping Gag-Pol precursor polyprotein coding region (Fig. 5). The p6 protein has a helix–flexible hinge–helix structure, 11 represents a docking site for several cellular and viral binding factors, and promotes the detachment of assembled virions from the cell surface and from each other. Two major p6 domains, the PT/SAPP motif at the N terminus required for virus detachment and the LXXLF motif required for incorporation of the Hiv-1 Vpr protein into budding virions, remain unchanged in PokA79.
Alignment of the deduced C-terminal protein sequences of Tat. Dots indicate identical amino acids. Dashed lines represent gaps. The specific Tat extension in the PokA79 sequence is marked.

Alignment of the deduced Gag p6 protein sequences. The seven amino acid-long insertion in the PokA79 sequence is marked.
Search of the putative protease and reverse transcriptase drug resistance mutations in the PokA79 pol gene using HIVdb program version 6.0.8 from the Stanford University HIV Drug Resistance Database 12 revealed the absence of primary drug resistance mutations in the protease, RT, and integrase sequences. The V77I secondary mutation, which is widely spread among HIV-1 subtype A variants in different FSU countries, was not found in the PokA79 protease sequence. At the same time, A62V and E138A substitutions were present in PokA79 RT. A62V is associated with multinucleoside resistance caused by the Q151M mutation, which was absent in the analyzed sequence. The E138A substitution recently has been added to the list of accessory NNRTI resistance mutations associated with decreased etravirine response in the DUET study. In the majority of HIV-1 subtype A1 variants from different FSU countries the V77I mutation is often linked to A62V mutation in RT. 3,13 The V77IA62V variant was found to predominate in Russia in terms of both its geographic distribution and number of cases caused. At the same time, viruses lacking the V77I and A62V mutations and viruses bearing only one of these mutations were also found, but all these variants were in the minority. The PokA79 sequence contains only the A62V substitution.
The V3 loop of the HIV-1 gp120 envelope glycoprotein is critical for virus infectivity as it is important for virus entry into the target cells, cell tropism, coreceptor usage, and syncytia formation. 14,15 Two amino acid tetrads on the V3 apex (positions N11–N18) are necessary for the fusion of the virus and the cell surface. The consensus sequence of this region for subtype A1 is SIRIGPGQ. GIHI, SVR/HI, SIHI/F, GVH/RI, GIR/HI, and other more rare motifs can also present at the N11–N14 positions of subtype A V3 loops. The new HIV-1 isolate PokA79 contains the GIHV sequence in positions N11–N14. GPGQ at positions N11–N18 is a more conservable motif (GPGR and other variants occur considerably less often). The N15–N17 positions of the V3 loop in the PokA79 sequence are occupied by a rare three-amino acid-long insertion GHT (a result of a GGACATACA nucleotide insertion) and only positions N18–N22 possess the GPGQ motif (Figs. 6 and 7). Insertions inside the V3 apex are rather rare for HIV-1 subtype A sequences; only about 50 sequences of all subtype A, A1, and A2 sequences from the HIV Sequence Database 10 contain short insertions in this region. The majority of such sequences have two-amino acid-long insertions GI (rarely RG or PG); five sequences from Central Africa obtained from symptomatic patients have three-amino acid-long insertions: the AJ404144 and AJ404175 sequences from the Democratic Republic of Congo and the U43136, L11457, and L11458 sequences from the Central African Republic. But the PokA79 insertion in this region differs from all other insertions in the sequences of African origin and seems to be unique. The functional role of these insertions and their influence on the virus entry process remain unclear.

Alignment of the nucleotide sequences of V3 loops. Insertions inside V3 loop apex are marked.

Alignment of the deduced protein sequences of V3 loops. Insertions inside V3 loop apex are marked.
The V3 loop sequence of PokA79 contains neutral amino acid N (asparagine) in position N5, positively charged amino acid K (lysine) in the N7 position, neutral amino acids G (glycine) and V (valine) in the N11 and N25 positions, correspondingly, neutral amino acid P (proline) in position N19, and has a net charge +8. Lysine in the N7 position is absent in CCR5-using viruses, as it increases the distance between V3 loop N7 and sulfotyrosine at CCR5 N14, thus abrogating their interaction. 15 The new PokA-79 laboratory strain actually uses the CXCR4 coreceptor for cell entry.
The highest rate of HIV-1 subtype A1 infection in Russia is observed among IDUs and their sexual partners. Therefore, it seemed reasonable for us to obtain a region-specific HIV-1 subtype A1 laboratory strain from native HIV-1 variants circulating in the IDUs cohort in the territory of central Russia. According to the literature, new laboratory HIV-1 strains capable of replicating in vitro are routinely isolated from the blood of a single infected patient. In this study we successfully used another approach and performed the isolation of a new subtype A1 strain from the lymphocyte pool of several HIV-1-infected patients. This approach may be a useful and timesaving method for the rapid isolation of new cell culture-adapted viral strains from small cohorts or homogeneous groups of patients.
In the present work we obtained and characterized for the first time in Russia a new stable laboratory HIV-1 subtype A1 strain from a mixture of primary virus isolates. This new strain can be used for the development and testing of region-specific vaccines against HIV-1 strains circulating in Russia as well as for viral biology research. The sequence analyses of the PokA79 full-length genome revealed several unusual features, distinguishing it from HIV-1 subtype A variants widespread in FSU countries. The revealed insertion in the PokA79 Gag p6 region does not affect functionally important motifs and lies in a region of known polymorphism, but the detected extension of the Tat protein and three-amino acid GHT insertion inside the V3 apex seem to be more interesting. The determination of the functional significance of the observed mutations in the PokA79 Tat and V3 loop regions and their possible contribution to selective advantages for facilitation of virus propagation in vitro remain topics of further research.
Sequence Data
The obtained PokA79 genomic sequence was deposited in the GenBank database under accession number FJ864679.
Footnotes
Acknowledgments
The authors would like to express their gratitude and acknowledgment of the generous contributions of the Foundation “Stiftung zur Förderung Immunologischer Grundlagenforschung,” Vaduz, Lichtenstein, for complete funding of the current research.
Author Disclosure Statement
No competing financial interests exist.