
Abstract
Cellular immune responses play a critical role in the control of human immunodeficiency virus type 1 (HIV-1), but less is known about the impact of transmission routes on immune defenses against HIV-1. Here, we report that subjects infected with HIV-1 through contaminated blood showed stronger HIV-specific T cell responses than those infected through mucosa, both in breadth (6.9±2.5 vs. 2.3±0.5, p=0.0293) and in magnitude [1270.0±544.9 vs. 409.5±121.3 SFU per million peripheral blood mononuclear cells (PBMCs), p=0.0223], by using a matrix of 404 overlapping peptides spanning all expressed HIV-1 proteins in an interferon (IFN)-γ enzyme-linked immunospot (ELISpot) assay. Our observation indicates that different mechanisms might be involved in the priming/generating of anti-HIV-specific T cell responses through different transmission routes.
HIV-1-
Blood samples from 32 chronic HIV-1-infected Chinese Han individuals [all on highly active antiretroviral therapy (HAART)] were obtained and investigated in this study, among whom eight patients were infected by contaminated blood (six FPDs and two intravenous drug users) and 24 were infected through sexual transmission (heterosexual and homosexual). Fresh peripheral blood mononuclear cells (PBMCs) were separated from whole blood using Ficoll-Hypaque (Sigma, St. Louis, MO) density gradient centrifugation and were stored in vapor-phase liquid nitrogen until use. Study subjects were recruited from the Beijing Youan Hospital. Relevant clinical data of all subjects are summarized in Table 1. The study was approved by the respective Institutional Review Boards of Beijing YouAn Hospital [IRB number 00004929], and all subjects gave written informed consent.
Measured at time of HIV-1-specific T cell analysis.
Between time of diagnosis of HIV/AIDS infection and time of HIV-1-specific T cell analysis.
Between time of diagnosis of HIV/AIDS infection and initiation of HAART treatment.
Who had undergone several treatment interruptions.
CHR, chronic; M, male; F, female; MSM, men who have sex with men; Hetero, heterosexual contact; FPD, former plasma donors; Ivdu, IV drug use; HAART, highly active antiretroviral therapy; LDL, lower than detectable levels.
A total of 404 peptides, overlapping by 10 amino acids, most of which were 18 amino acids in length and spanned the entire expressed HIV-1 clade B genome (Gag, Pol, Vif, Vpr, Vpu, Rev, Tat, Env, and Nef ), and 124 matrix peptide pools were modeled as described previously, 5 with each of the 404 peptides represented in three different pools (three-dimension or three-dimensional peptide matrix) and with every pool including 8 to 11 different peptides. All peptides and peptide pools were then frozen at −80°C until use. ELISpot assays were performed as described previously. 6 We used ELISpot for the Human Interferon-γ kit (Mabtech, Sweden) and 96-well Multiscreen HTS IP plates (MSIP4510; Millipore, United Kingdom) in this section. The PBMCs were plated at a 0.5×105 to 1×105 viable PBMC per well and the final concentration of the peptides in the well was 2 μg/ml. The spots were developed with AP conjugate substrate kit (Bio-Rad Laboratories; Hercules, CA). Plate results were read using a ChamSpot III ELISpot Reader System (Sage Creation, Beijing, China) and the number of specific T cells was calculated by subtracting the negative control values. The ELISpot data are expressed as the number of spot-forming units (SFU) per million PBMCs. Responses were regarded as positive if they had at least three times the mean number of SFU in six control wells and had to be >50 SFU/106 PBMCs.
Data are presented as mean±standard error of the mean. Statistical analysis and graphical presentation were performed using SPSS 13.0 (SPSS Inc., Chicago, IL) and GraphPad Prism 5.0. The data were analyzed using the Student's t-test with significance at p<0.05.
Using the afore-mentioned three-dimentional peptide matrix approach with ELISpot assays, we screened a total of 32 HIV-1-infected individuals for T cell responses against the entire expressed HIV-1 consensus clade B genome on a single peptide level. Through analysis of the data, we found that the majority of the HIV-1 protein subunits are targeted by virus-specific T cell responses, and the overall peptide recognition within all studied subjects is dominant in Nef, Vpu, and Gag proteins, as reported previously. 7 We have identified 67 individual overlapping peptides given responses at least once over the total 404 peptides (67/404, 16.6%).
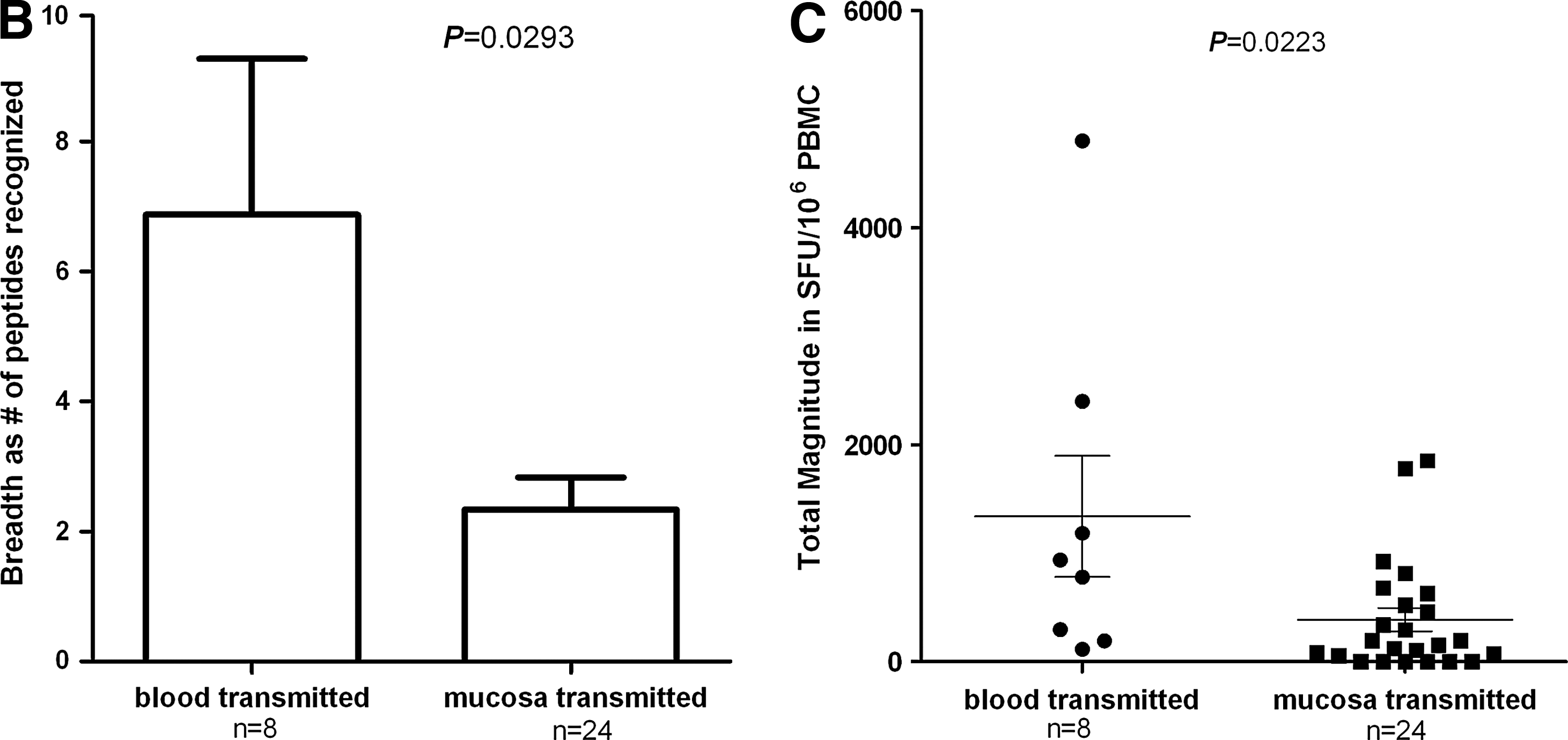
The specific T cell responses within individual subjects are depicted in Fig. 1A in breadth and total magnitude, respectively. Variables listed in Table 1 were demonstrated to be evenly distributed between the blood-borne and mucosa transmission groups, specifically 98.0±17.0 vs. 81.0±8.3 (m), p=0.249 for duration of diagnosis, 42.3±17.4 vs. 33.9±6.6 (m), p=0.879 for timing of HAART initiation, 45.6±7.9 vs. 40.3±3.8 (m), p=0.557 for duration of HAART, and 401.9±52.5 vs. 422.1±34.4 (cells/mm3), p=0.948 for CD4 count. Even though there were no significant differences in the clinical data among our studied subjects between transmitted routes, former plasma donors together with IV drug users presented a stronger HIV-1-specific T cell response in total. The HIV-1-infected subgroup infected through contaminated blood recognized more overlapping peptides than mucosa, expressed through the number of responsive overlapping peptides (6.9±2.5 vs. 2.3±0.5, p=0.0293, Fig. 1B). Also, the HIV-1-infected subgroup through contaminated blood gave stronger specific T cell responses based on the total magnitude of response within each patient (1270.0±544.9 vs. 409.5±121.3 SFU per million PBMCs, p=0.0223, Fig. 1C).

Comparison of breadth and magnitude of HIV-1-specific T cell responses between the HIV-1-infected study subgroups infected through contaminated blood and through sexually related mucosal transmissions.
In this study, we have shown that HIV-1-infected subjects infected through contaminated blood give a broader HIV-1-specific T cell response with higher magnitude in comparison with mucosal transmission through sexual relations, in which we have excluded variables (importantly as duration of diagnosis, timing of HAART initiation) that could account for the observed difference in interferon (IFN)-γ response showing the groups are comparable based on the data provided. Many host and virological factors could determine the quantity and quality of the overall T cell responses such as individual genetic background, infecting virus, and infection routes.
Researchers have found only single or very limited numbers of transmitted viruses during acute HIV-1 infection in most patients infected through sexual transmission. 4 Indeed, around 80% of cases of heterosexual transmission have shown that a single virus establishes infection, 4,8 which is consistent with the epidemiological observation of the laggard efficiency of HIV-1 transmission through most sexual routes. 2,3 In the present study, 32 HIV-1-infected chronic individuals infected from different route were studied. We found that T cell responses had limited breadth and magnitude among subjects infected through sexual routes (homosexual and heterosexual), compared to subjects infected through contaminated blood. The lower HIV-1-specific T cell responses could be due to the single or narrowed transmitted virus during acute HIV-1 infection.
The route of infection and microenvironment for antigen delivery have been shown to have an important impact on the magnitude and quality of the T cell responses in an Adenovirus-Based HIV-1 Vaccine study, most likely due to the differences in the capacity of T cell priming 9 ; low-frequency, monofunctional CD8 T lymphocytes with restricted anatomic trafficking patterns were observed in mucosal routing of vaccination, which is in agreement with our observation concerning the lower magnitude and narrower T cell responses in patients infected through sexual transmission when compared with blood-borne transmission. The mechanisms that underlie this, such as antigen presentation and T cell priming in the local microenvironment, merit further investigation. Ideally it would have been preferable to study untreated patients with similar infection times and viral loads, but such a cohort was inaccessible for this study, particularly in the identification of untreated patients with blood-borne transmission.
Irrespective of these considerations, we believe that there is still an advantage in comparing the patients with HAART treatment, as it is known that HAART treatment would affect the strength of the T cell responses, mainly due to reduced viral load, therefore reducing the antigen stimulation required for T cell expansion. 10 –12 In our case, most (almost all) patients (both infected through contaminated blood and sexual relations) are on long-term treatment, with either a low or undetectable viral load, and therefore the amount of antigen for T cell stimulation/expansion in the blood should be comparable between the two groups. This suggests that the differences we saw between the two groups could not be due to treatment but more likely represent real differences due to the nature of the transmission route.
In conclusion, this study provides a comprehensive analysis of the profile of HIV-1-specific T cell responses in a group of Chinese Han subjects infected with HIV-1 through different infection routes and has observed broader and stronger HIV-1-specific T cell responses in subjects infected through contaminated blood rather than through sexual transmission.
Footnotes
Acknowledgments
We would like to thank Prof. Ning Li and Prof. Hao Wu for clinical database support and Dr. Xiaojie Huang and Ms. Yan Fu for following up with patients. We would also like to thank all the participants and all those working for this study. All studies were conducted in the Research Centre of Infection & Immunity at Beijing YouAn Hospital with support from Beijing Municipal Science & Technology Committee, China (Grants Z080507030808020 and D09050703590904) and the China National Science & Technology Key Program (Grant 2008ZX10001-003 and 2012ZX10001001-008).
Author Disclosure Statement
No competing financial interests exist.