
Abstract
The antibody-dependent cell-mediated cytotoxicity (ADCC) mechanism involves both innate and adaptive immune systems. While a number of epitope mapping studies of neutralizing (Nt) antibodies and cytotoxic T lymphocyte (CTL) against a variety of HIV-1 clades have been reported, there has been a paucity of similar studies aimed at identifying epitopes of ADCC-inducing antibodies. Herein we screened 35 sera from HIV-1 CRF01_AE-infected blood donors for ADCC antibody activity against gp120 utilizing an EGFP-CEM-NKr flow cytometric assay. Eighteen sera with high ADCC antibody activity were then comprehensively examined for ADCC antibody epitopes using the HIV-1 subtype CRF01_AE TH023 gp120 peptide set consisting of 126 peptides of 15 amino acids in length, overlapping by 11 amino acids. This peptide set was divided into five pools (E1–E5). Each positive peptide pool was further investigated for fine epitope mapping of ADCC antibody activity using a 5 by 5 peptide matrix format. Interestingly, six and three peptides from peptide pools E1 and E2, respectively, responded to at least 33.33% of the tested sera. These nine common ADCC epitopes were localized to the C1 and V2 region of gp120. Furthermore, 5/9 epitopes were also shown to serve as full or partial Nt antibody targets for HIV-1 subtypes B and CRF01_AE. We submit these data on epitope mapping of ADCC or dual ADCC-Nt antibodies against HIV-1 gp120 that should be considered in the formulation of vaccines against HIV-1.
Introduction
HIV-1
This view is supported by the previously published findings of Baum et al. who reported that a correlation exists between titers of HIV-1 gp120-specific ADCC antibody and the rate of disease progression. 5 Thus, rapid progressors had significantly low ADCC antibody levels against gp120-coated CEM-NKr target cells compared with levels found in the sera of typical progressors. In addition, HIV-1 elite controllers who have undetectable viremia showed higher ADCC antibody titers than infected individuals with viremia. 6 Further support for the importance of ADCC functioning antibodies is derived from the findings in the SIV-infected nonhuman primate model. Thus, vaccinated rhesus macaques elicited ADCC antibodies with titers that correlated with lower viremia after mucosal challenge with a pathogenic SIV strain. 7 A correlation between the levels of nonneutralizing ADCC antibodies with significant control of viremia against SIV infection has also recently been published. 8 –10 It is of interest to note that the ability of a broadly neutralizing monoclonal antibody to confer protection appears to require the interaction between the Fc portion of the antibody and the Fc receptor-bearing effector cells. 11 These studies in concert demonstrate the positive impact of ADCC on HIV.
ADCC is an immune response that combines components of both the innate and humoral immune system. Thus, in the case of HIV infection, ADCC can occur by the binding of HIV-specific antibody to HIV antigen expressed on the cell surface of infected cells. The Fc portion of the bound antibody can then bind to Fc receptors expressed on the surface of various lineages of innate immune effector cells that predominantly include the natural killer (NK) cells but also macrophages and neutrophils. It is thus reasonable to assume that the identification of the epitopes of HIV that contribute to ADCC activity would be important new information that can be utilized for vaccine development. In efforts to address this issue, our laboratory has established the EGFP-CEM-NKr flow cytometric method to measure NK cell-mediated ADCC antibody activity against HIV-1 gp120. 12 We have utilized this newly established assay to identify common ADCC antibody epitopes of the gp120 of HIV-1 CRF01_AE. Results of these studies constitute the basis of this report.
Materials and Methods
Volunteer samples
All sera samples from the blood donor volunteers were obtained following approval from the Institutional Review Board of the Faculty of Medicine Siriraj Hospital, Mahidol University prior to initiation of this study. The sera samples obtained had no donor identifiers.
Cell line
The natural killer cell resistant EGFP-CEM-NKr cells
12
were cultured in growth medium containing RPMI 1640 (Gibco, Grand Island, NY) supplemented with 2 mM
Peptides
The gp120 of HIV-1 CRF01_AE CM243 (Protein Sciences, Meriden, CT) was used to pulse the EGFP-CEM-NKr cells. In addition, the complete peptide set based on the HIV-1 subtype CRF01_AE TH023 gp120 sequence consisting of 126 peptides 15 amino acids in length, overlapping by 11 amino acids, was kindly provided by the Natural and Medical Sciences Institute (University of Tűbingen, Germany). These peptides were divided into five pools (pools E1–E5) each containing 26–27 peptides. To further identify the reactive peptide, each informative peptide pool was again divided into 10 small pools according to the 5 by 5 matrix formats. Each peptide pool was also used to pulse the EGFP-CEM-NKr cells.
Preparation of sera
Thirty-five HIV-1-seropositive sera of blood donor volunteers were kindly provided by the Department of Transfusion Medicine, Faculty of Medicine Siriraj Hospital. All sera were from individuals infected with HIV-1 CRF01_AE as determined by C2-V4 nucleotide sequencing as described elsewhere. 13 An HIV-1-seronegative pooled serum was kindly provided by the serology laboratory, Department of Microbiology, Faculty of Medicine Siriraj Hospital. This pool of HIV-negative sera was aliquoted and an individual aliquot was used as the negative control with each assay. Similarly, an aliquot of a single source of an HIV-1 CRF01_AE-seropositive serum with high ADCC antibody activity was used as the positive control with each assay. All sera were inactivated at 56°C for 30 min prior to use and used at dilutions of 1:250, 1:2,500, and 1:25,000 in the assays.
Target cells
The gp120- or peptide pool-pulsed EGFP-CEM-NKr target cells were prepared by the addition of either 0.1 μg gp120 of HIV-1 CRF01_AE CM243 or various concentrations ranging from 0.02 μg to 10 μg of each of the five peptide pools of HIV-1 CRF01_AE TH023 gp120 to 1×105 target cells. The peptide and target cells were incubated for 1 h at room temperature. The gp120 pulsed cells were washed three times with phosphate-buffered saline (PBS). The target cells were then resuspended in growth medium at a concentration of 1×105 cells/ml.
Effector cells
Cryopreserved peripheral blood mononuclear cells (PBMCs) were prepared from leukopaks of a single HIV-1-seronegative blood donor as previously described. 12 The cells were later thawed and overlaid overnight at 1×106 cells/ml in growth medium at 37°C with 5% CO2. The cells were then resuspended in growth medium at a concentration of 2.5×106 cells/ml and used as effector cells.
EGFP-ADCC flow cytometric assay
The EGFP-ADCC flow cytometric assay was based on a method previously described. 12 Briefly, the sera at various dilutions were incubated with gp120 pulsed EGFP-CEM-NKr target cells and then 100 μl of effector cells was added to provide an effector/target ratio (E/T) of 50/1 and incubated at 37°C, 5% CO2 for 1–2 h. In case of peptide pools, each peptide pool was used to pulsed target cells and incubated for 3–4 h. An HIV-1 CRF01_AE-seropositive serum and an HIV-1-seronegative pooled serum were examined simultaneously as positive and negative controls. Lysis of target cells by ADCC activity was measured and analyzed with a FACScan flow cytometer [Becton Dickinson Biosciences (BDB), San Jose, CA]. Data analysis for two parameter dot plots of EGFP versus PI was performed using CELLQuest (BDB) software to enumerate live versus dead target cells. Debris and dead cells were excluded by setting a threshold of forward light scatter. Instrument settings and two-color compensation were performed prior to analyzing the samples using CaliBRITE beads (BDB). Percent lysis of target cells at each serum dilution was calculated by the following formula: % lysis=(UR/UR+LR)×100 where UR represented compromised target cells and LR represented intact target cells. Subtraction of the percent lysis of target cells in the absence of serum from that of target cells with tested sera gives the actual percent lysis of ADCC activity. The cut-offs for positive results using gp120 pulsed target cells and peptide pool pulsed target cells are 7.21% lysis and 5.46% lysis, respectively.
Results
Screening for high ADCC antibody activity in HIV-1-seropositive sera using gp120 pulsed target cells
The serologically positive sera from 35 blood donors infected with HIV-1 CRF01_AE were examined for ADCC antibody activity against gp120 pulsed target cells using the EGFP-CEM-NKr flow cytometric assay. 12 The percent lysis of target cells ranged from 9.43% to 51.77% (data not shown). Eighteen of the 35 sera with relatively high ADCC antibody activity (percent target cell lysis higher than mean value: 39.38%) were further investigated for ADCC antibody activity against each of the five peptide pools of HIV-1 CRF01_AE TH023 gp120 formulated as described in the Materials and Methods section.
Optimal concentration of the 15-mer amino acid peptide pool for pulsing target cells
In efforts to define the optimal concentration of the 15-mer amino acid peptide pool for pulsing target cells, various amounts of each of the peptide pools ranging from 0.02 μg to 10 μg were analyzed. The peptide pool concentration at 1 μg demonstrated the highest ADCC activity at a dilution of 1:2,500 and 1:25,000 of the positive sera (Fig. 1). This peptide concentration was thus subsequently used for a more detailed study of the ADCC antibody responses to the pools of the HIV-1 gp120 peptides.
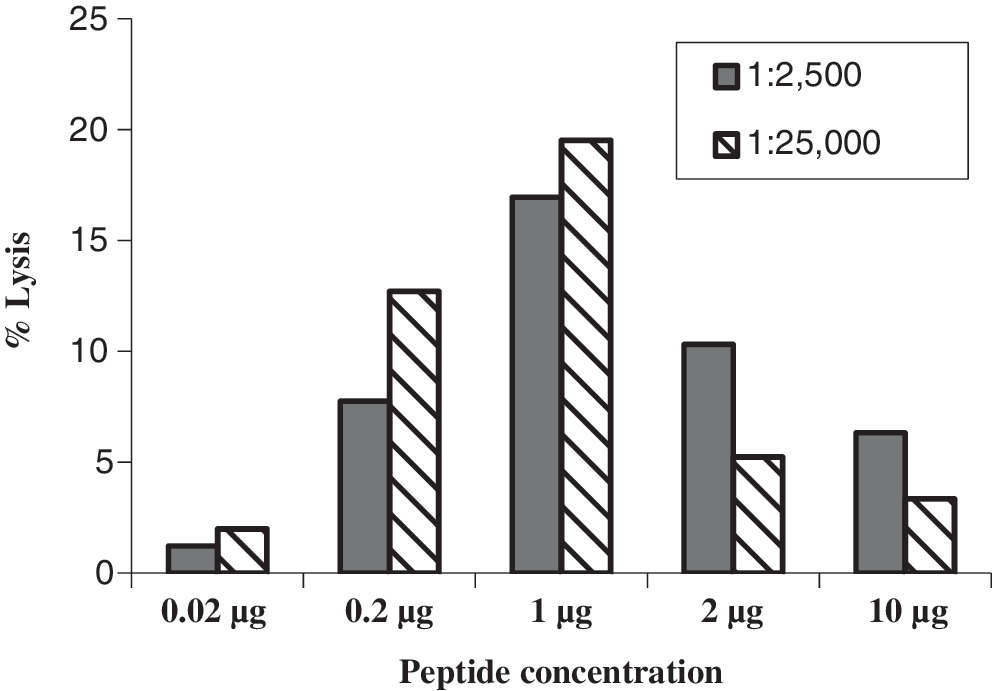
Determination of the optimal concentration of the 15-mer amino acid peptide pool used for pulsing target cells. Various concentrations of peptide pool, i.e., 0.02 μg to 10 μg, were used to pulse target cells and then examined for antibody-dependent cell-mediated cytotoxicity (ADCC) antibody activity using an HIV-1-seropositive at serum dilutions of 1:2,500 and 1:25,000 and effector cells from a healthy individual. The results displayed are representative of three separate experiments.
Identification of HIV-1 CRF01_AE gp120 epitopes that are the target of ADCC reactive antibodies
Eighteen sera with high ADCC antibody activity were examined against each of five peptide pools (pools E1–E5) of the HIV-1 CRF01_AE Env TH023 gp120 peptide set. The cut-off for positive results using peptide pool pulsed target cells is 5.46% target cell lysis (Fig. 2). This cut-off value was obtained from examining the ADCC activities of 30 seronegative sera against peptide pools and the average ADCC activity was 3.962±1.498 (data not shown). Thus, we used the cut-off value at>mean+SD. These 18 sera demonstrated positive ADCC antibody responses against a number of the peptide pools as shown in Table 1. Thus, whereas 14 (77.78%) and 12 (66.67%) sera demonstrated significantly higher frequency of ADCC antibody responses against peptide pools E1 and E2, respectively, eight to nine (44.44–50.00%) of these sera showed significant but somewhat lower frequency of ADCC antibody responses to peptide pools E3–E5. Thus, a 5 by 5 matrix of each of the two peptide pools E1 and E2 was selected to perform the fine epitope mapping of ADCC antibody responses. The potential ADCC antibody epitopes were identified by verifying the common peptides from the matrix (Fig. 2). The common peptides within peptide pools E1 and E2 that responded to ADCC antibody activity of each of the 17 sera tested are shown in Table 2. Nine potential ADCC epitopes (peptides 6, 8, 11, 13, 16, 23, 40, 41, and 43) with the antibody responses from at least 33.33% of the tested sera were identified in this study. For the parent E1 peptide pool, nine (50%) sera showed ADCC activity against peptide 16; eight (44.44%) sera showed ADCC activity against peptides 6, 8, 11, and 13; and seven (38.89%) sera showed ADCC activity against peptide 23. For the parent E2 peptide pool, 6 (33.33%) of the sera showed ADCC activity against peptides 40, 41, and 43.
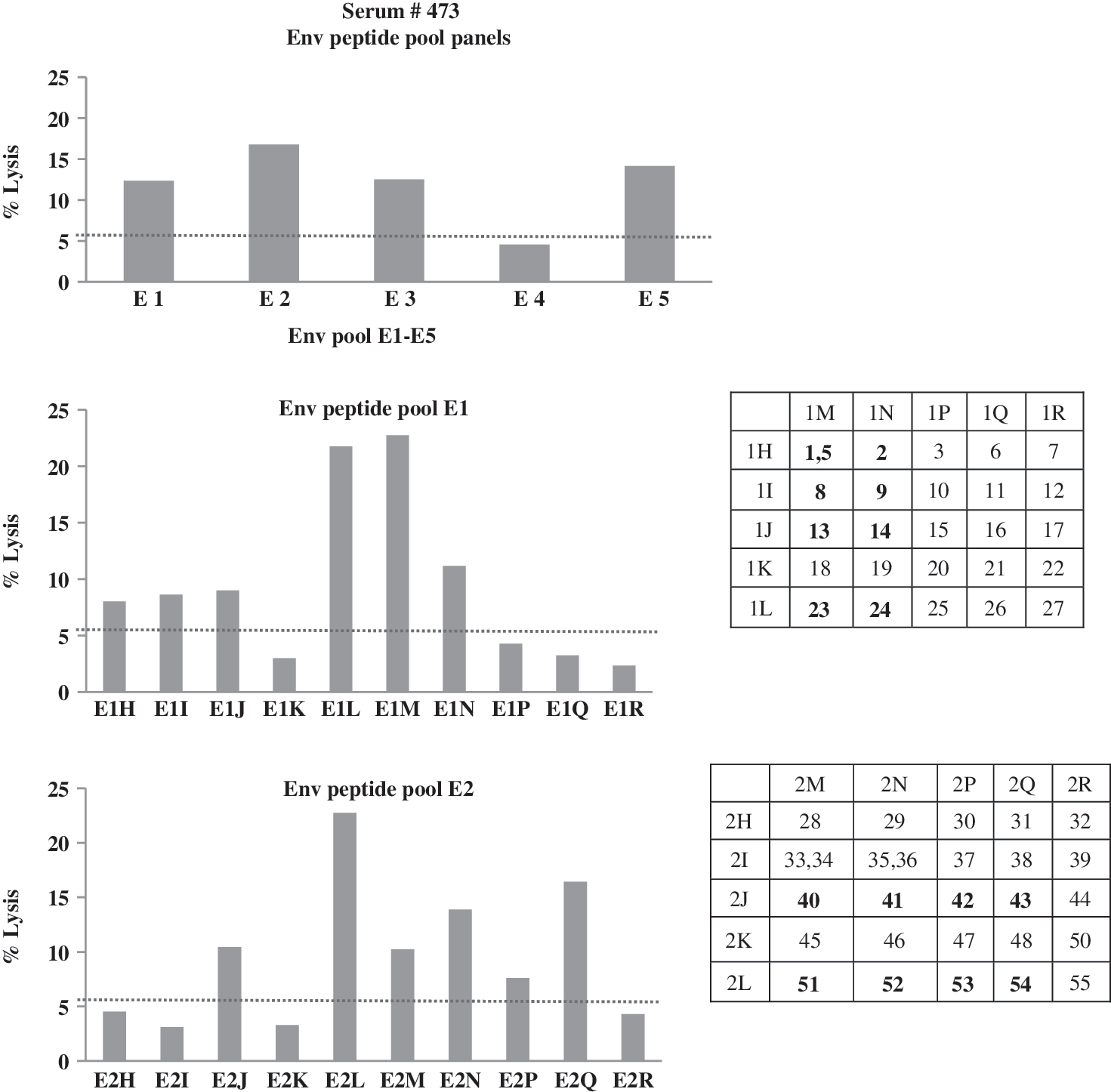
Identification of ADCC antibody epitopes using serum from HIV-1-seropositive blood donor #473. Positive results are shown as the mean % lysis of target cells of duplicate tubes greater than the cut-off value at 5.46% (dotted line) after background subtraction, performed in triplicate (upper graph). Individual peptide responses were assessed by testing with a 5 by 5 matrix of each Env pool of E1 and E2 (lower graphs). The common positive peptides of matrix are shown in bold.
High antibody-dependent cell-mediated cytotoxicity (ADCC) antibody activity against HIV-1 gp120 pulsed target cells of 18 HIV-1-seropositive sera was analyzed for individual responses to target cells pulsing with each of five peptide pools (E1–E5) comprising the complete HIV-1 subtype CRF01_AE TH023 gp120 peptide set consisting of 126 peptides 15 amino acids in length. Each pool contains 26–27 peptides.
The peptide epitopes that showed ADCC targeted responses against at least 33.33% of the sera tested are shown in bold.
The consensus of the potential ADCC antibody epitopes of HIV-1 CRF01_AE gp120 and the published Nt antibody epitopes of HIV-1 subtype B gp120
The amino acid sequences of the nine ADCC epitopes of HIV-1 CRF01_AE gp120 along with the previously published neutralizing or binding antibody epitopes of HIV-1 subtype B gp120 are shown in Table 3. The range of ADCC antibody responses against the tested peptide pools was 6.41–38.29% target cell lysis. As seen, the common peptide epitopes from pool E1 (peptides 8, 11, 13, 16, and 23) include the last five amino acids at the C terminal of peptide 6, which are localized within the C1 region of gp120, the common peptides 40, 41, and 43 of pool E2 are localized within the V2 loop of gp120. Five (peptides 16, 23, 40, 41, and 43) and three (peptides 8, 11, and 13) ADCC epitopes were full or partial consensus with Nt antibody epitopes and binding antibody epitopes, respectively, of HIV-1 subtype B. The distributions of ADCC epitopes of HIV-1 CRF01_AE from this study and the previously described Nt antibody or binding antibody epitopes of HIV-1 subtype B across the gp120 gene are illustrated in Fig. 3. The conservation of nine potential ADCC epitopes that are located in the gp120 region of HIV-1 CRF01_AE TH023 and CM243 is shown in Table 4.
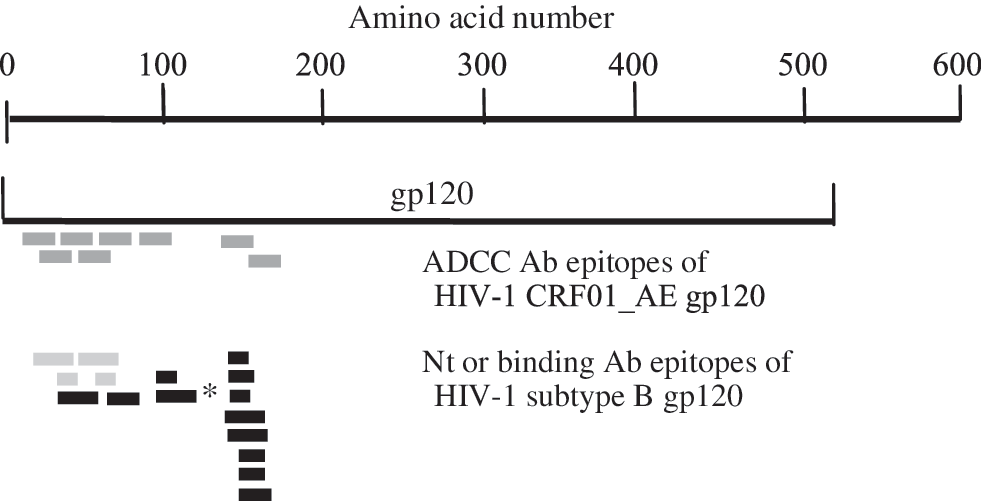
Epitope distribution across the gp120 of HIV-1. ADCC antibody epitopes (dark gray) of HIV-1 CRF01_AE identified in this study and previously identified Nt (black) or binding (gray) antibody epitopes of HIV-1 subtype B are shown. The epitope sizes are not drawn to scale and the amino acid locations of the epitopes are aligned as closely as possible with the true alignment. *This peptide was published as Nt antibody epitope of HIV-1 CRF01_AE gp120. 13
Cut-off for positive result is 5.46% target cell lysis.
More references for HIV-1 subtype B gp120 Nt epitopes could be found at the Los Alamos Database. 25
This peptide was published as the Nt antibody epitope of HIV-1 CRF01_AE gp120 13 and the aa # is according to the sequence of HIV-1 CRF01_AE gp120.
Sequences with amino acid (aa) numbers according to the total responders and range of responses compared to those reported for neutralizing (Nt) or binding antibody epitopes of HIV-1 subtype B gp120. Amino acid numbers are according to the Los Alamos Database. 25
aa, amino acid; C1, conserved region 1; V2, variable region 2; MAb, monoclonal antibody; Nt Ab, neutralizing antibody.
Accession number: L03703.
Accession number: HQ691028.
Discussion
In the studies reported herein, we screened 35 sera of blood donors infected with HIV-1 CRF01_AE for high ADCC antibody activity against gp120 pulsed target cells using the EGFP-CEM-NKr flow cytometric assay. 12 This assay measures the number of target cells that are killed. The gp120 was used instead of gp160 because only HIV-1 CRF01_AE gp120 is commercially available. We further developed this method to measure the ADCC antibody activity against a battery of overlapping 15-mer amino acid peptides encompassing the whole gp120 in addition to the unfractionated gp120 molecule. A study to optimize the antigen concentration for the ADCC assay was performed. The optimal amount of the 15-mer peptide required to pulse the target cells is a log higher than that of gp120, i.e., 1 μg vs. 0.1 μg (Fig. 1). This may be due to the length of peptide used to pulse target cells. The receptor-binding surfaces on gp120 are formed by discontinuous amino acids in various regions of this glycoprotein for binding to the CD4 molecules on the target cells 14 while the 15-mer peptides are able to bind to only a select portion of the CD4 molecules. Thus, the avidity of gp120 that binds to targets is likely to be significantly higher than that of the 15-mer peptides. Therefore, following the addition of the HIV-positive serum, the large spectrum of specific antibodies could bind to gp120 on the surface of target cells, which would be greater than the few clones of antibodies that bind to the 15-mer peptides on the surface of target cells.
Eighteen of the 35 sera with high ADCC antibody activity against gp120-coated target cells were further investigated for ADCC antibody activity against 15-mer peptide pools (pools E1–E5) of the HIV-1 CRF01_AE TH023 gp120 peptide set. Interestingly, peptide pools E1 and E2 were recognized by the majority of tested sera, i.e., 14/18 and 12/18, respectively (Table 1). Moreover, all tested sera except serum #608 responded to at least one of these two peptide pools. Identification of potential ADCC antibody epitopes using a matrix format led to the finding that the spectrum of peptide-specific reactivity varied for each of the sera tested and that select sera such as serum #041 showed reactivity against a total of 29 peptides (Table 2). These data indicate that the number of ADCC antibody reactive epitopes is likely much larger compared to those reported for neutralizing (Nt) antibodies. 15 A low level of ADCC antibody may be required for effector cells to kill virus-infected cells and it is not necessary for all of the viral epitopes to be bound by antibody. 16
Results of the studies further showed that six epitopes (peptides 6, 8, 11, 13, 16, and 23) of peptide pool E1 and three epitopes (peptides 40, 41, and 43) of peptide pool E2 were identified as being targets of ADCC antibodies from at least 33.33% of the sera tested (Table 2). These epitopes are localized to the C1 and V2 loops of gp120, respectively (Table 3). The surface gp120 subunits of the HIV envelope protein have been reported to serve as targets of ADCC antibody responses since 1989 17 using the complicated standard chromium release assay measuring the killing of 51Cr radio-labeled target cells. Recently, a number of simpler methods have been advanced for the identification of ADCC antibody activity and/or epitopes. These include the rapid fluorometric ADCC assay, 18 the EGFP-ADCC flow cytometric assay, 12 the NK cell activation ADCC assay, 19 and the modified ADCC GranToxiLux assay. 20
The conformational C1 region on the surface of HIV-1-infected cells is recognized by the nonneutralizing A32 mAb, which mediates potent ADCC activity. 21 We submit that our common six linear epitopes (peptides 6, 8, 11, 13, 16, and 23) localized to the C1 region may encompass the same epitopes recognized by the A32 mAb. Interestingly, A32-like antibodies were observed to mediate the majority of total ADCC activity in 50% of the chronically infected subjects 21 while ADCC antibodies from at least 38.89% of tested sera from asymptomatic infected blood donors were shown to recognize these common epitopes in our study. These findings indicate the possibility that the ADCC antibody responses to this C1 region may occur throughout the course of HIV infection. Moreover, the common epitope peptide 23 (aa 89–103, TENFNMWKNNMVEQM) has been reported for ADCC antibody responses in 5/22 (22.72%) HIV-infected subjects. 22 The results of our studies showed that 7/18 (38.89%) of the sera had reactivity that mapped to this peptide (Tables 2 and 3). Mutations of this peptide sequence in the context of the conformation gp140 demonstrated its ability to escape from ADCC immune pressure as confirmed by using a killing-based ADCC assay. 22 This epitope may be one of the main targets for ADCC antibody response in vivo.
Ninety-seven percent (31/32) of the vaccine recipients of the Thai phase III clinical trial RV144 had binding antibody responses against cyclic peptides of the V2 loop at 2 weeks postimmunization. 23 The majority of antibody responses targeted the crown of the V2 loop (KQKVHALFYKLDIVPI). This sequence is almost identical to the common ADCC epitope peptide 43 (KKQKVHALFYKLDIV) identified in 6/18 (33.33%) sera tested in our study (Tables 2 and 3). There is an α4β7 binding motif (LDV) in this region that could bind HIV-1 at the gut mucosal cell surface. Thus, either binding antibody or ADCC antibody to this epitope could have an effect on free virion or HIV-infected cells at the port of entry.
The percent sequence divergence between the gp120 of HIV-1 CRF01_AE CM243 used for screening of ADCC antibody responses and the gp120 of HIV-1 CRF01_AE TH023 peptide set was only 8.6, thus allowing epitope mapping of ADCC responses. Interestingly, when the amino acid sequences of identified epitopes from both strains were compared, six of nine main ADCC antibody epitopes (peptides 6, 8, 11, 40, 41, and 43) of both strains are identical (Table 4). In addition, the other three epitopes (peptides 13, 16, and 23) demonstrated only one amino acid difference. Moreover, when the amino acid sequences of a recent strain of HIV-1 CRF01_AE, i.e., T504258_sga07 (sampling year 2008; percent sequence divergences between this gp120 and those of CM243 and TH023 were 18.9 and 21.9, respectively) were included for comparison, all nine epitopes still look similar (Table 4). The amino acid sequences of these epitopes may be conserved among different strains of HIV-1 CRF01_AE. This may also explain how most tested sera obtained in the years 2008–2010 could respond to the peptide set of TH023 (Table 2). Further studies on ADCC antibody responses to autologous peptide sets matched to the infecting strains of each individual would provide a better understand of ADCC responses.
Neutralizing antibody and ADCC antibody activity are often mediated by different specificities. However, the regions of the HIV-1 Env that are the target of Nt antibody and ADCC antibody against gp120 are often overlapping. 24 The amino acid sequences of ADCC epitopes investigated in this study are fully or partially in consensus with those Nt antibody epitopes of HIV-1 subtype B or CRF01_AE that have been previously reported 13,25 (Table 3 25 –38 and Fig. 3). Five of the nine (55.56%) ADCC epitopes are overlapping with published Nt epitopes. It is thus possible for one clone of antibody to have the ability to both neutralize virus and mediate ADCC.
Since cell-to-cell transmission appears to be more efficient than that of free virion for HIV infection, 39 the ADCC responses would be a very important mechanism to eliminate the small number of virus-infected cells from an infected partner before the establishment of infection. Although, Nt antibodies have no effect on cell-to-cell transmission of HIV-1, 40 their responses would be very crucial for eliminating free virions that may enter simultaneously with the viral infected cells during viral transmission. We submit that the common ADCC and dual ADCC-Nt antibody epitope information demonstrated in this study would be useful for the design of future HIV-1 vaccine candidates.
Footnotes
Acknowledgments
We sincerely thank professor Aftab A. Ansari of the Department of Pathology and Laboratory Medicine, Emory University School of Medicine, Atlanta, GA, for his valuable comments and discussions. We would also like to thank all HIV-1-infected blood donors. This work is supported by the Thailand Research Fund for which K.P. is the TRF-senior research scholar. This work is a part of S.A.'s research thesis at the Faculty of Graduate Studies, Mahidol University.
Author Disclosure Statement
No competing financial interests exist.