
Abstract
The vast diversity of HIV-1 infections has greatly impeded the development of a successful HIV-1/AIDS vaccine. Previous vaccine work has demonstrated limited levels of protection against SHIV/SIV infection, but protection was observed only when the challenge virus was directly matched to the vaccine strain. As it is likely impossible to directly match the vaccine strain to all infecting strains in nature, it is necessary to develop an HIV-1 vaccine that can protect against a heterologous viral challenge. In this study we investigated the ability of polyvalent and consensus vaccines to protect against a heterologous clade B challenge. Rhesus macaques were vaccinated with ConB or PolyB virus-like particle vaccines. All vaccines were highly immunogenic with high titers of antibody found in all vaccinated groups against SIV Gag. Antibody responses were also observed against a diverse panel of clade B envelopes. Following vaccination nonhuman primates (NHPs) were challenged via the vaginal route with SHIVSF162p4. The PolyB vaccine induced a 66.7% reduction in the rate of infection as well as causing a two log reduction in viral burden if infection was not blocked. ConB vaccination had no effect on either the infection rate or viral burden. These results indicate that a polyvalent clade-matched vaccine is better able to protect against a heterologous challenge as compared to a consensus vaccine.
Introduction
It is estimated that 33 million people worldwide are currently living with HIV-1 with 1.9 million people becoming newly infected in 2009, highlighting the need for a preventative vaccine. 1 One of the greatest struggles against developing an HIV-1 vaccine is the large diversity of viral isolates with differences in envelope sequences, which differ as much as 10% within a given clade and 35% across clades. 2 Previous vaccine studies in nonhuman primates (NHPs) demonstrated sterilizing immunity, but protection was observed only when the vaccine was exactly matched to the challenge strain. 3 –8 An effective HIV/AIDS vaccine will need to protect against heterologous viral challenges.
A number of various strategies have been investigated to address the issue of Env diversity. 9 Polyvalent vaccines are an effective strategy to protect against a number of infections including pneumococcus, influenza, and polio. 10 Polyvalent vaccines are typically composed of multiple copies of a given target(s), thereby increasing the diversity of the epitopes presented to the immune system. If the diversity of the epitopes is large enough within the polyvalent vaccine, it can present one or more epitopes present in any given isolate. Polyvalent HIV/AIDS vaccines do increase the breadth and strength of both cellular and humoral immune responses compared to monovalent vaccines. 11 –20
Another strategy to address the issues of Env diversity is the construction of envelope antigens based upon a consensus sequence derived from numerous HIV-1 isolates. These vaccines utilize a consensus sequence that has been artificially generated to represent the most common amino acid at each position of a given target from a collection of sequences. The goal of this strategy is to minimize the genetic difference between the vaccine strain and any given primary isolate. Previous studies have indicated that consensus Env proteins are functional and highly immunogenic. 15,21 –27 Consensus vaccines can induce a broader immune response as compared to a primary isolate. 15
The first aim of this study was to compare the ability of a consensus clade B (ConB) and a polyvalent clade B (PolyB) Env vaccine to develop a broadly reactive immune response in an NHP model. Both vaccines were delivered on the surface of a virus-like particle to facilitate the presentation of envelope in its native conformation. The second aim was to determine the ability of a consensus and polyvalent vaccine to protect against an SHIV challenge. Following vaccination, all NHPs were challenged with an SHIVSF162p4 via the intravaginal route. SHIVSF162p4was heterologous to both the ConB and PolyB vaccines thus better representing a potential transmission event. The vaginal route was chosen as this is the most common transmission route worldwide. 28 This is the first study to directly compare the breadth of immunity generated by a consensus and polyvalent vaccine in an NHP model.
Materials and Methods
DNA plasmids
The pTR600 vaccine plasmid 29 and the HIV-1 virus-like particle (VLP)-expressing plasmid have been previously described. 30 Briefly, the pHIV-wtVLPADA plasmid encodes for the following gene sequences: HIV-1BH10 gag–pol (pHIVBH10 nt 112–3626) (accession number M1564) and HIV-1ADA vpu, env, rev, tat (nt 5101–8159). Safety mutations were engineered into Gag to prevent viral RNA packaging 31,32 and RT to prevent reverse transcriptase and RNase H activity (pHIV-VLPADA). 33 –35 A codon-optimized SIVMac239 p55 Gag gene (generous gift from Dr. Andrea A. Gambotto) was cloned into pTR600 to generate the SIV Gag VLP. Each VLP was expressed from a cytomegalovirus immediate-early promoter (CMV-IE) for initiating transcription of eukaryotic inserts and the bovine growth hormone polyadenylation signal (BGH poly A) for termination of transcription.
Consensus VLPs were constructed by substituting ADA env with the consensus env sequence from the consensus clade B envelopes (LANL database). These sequences represent the most common amino acids found at each location within the Env gene from over 200 isolates for each clade. The primary isolate VLPs were constructed by substituting ADA env with the primary env sequence. These isolates were obtained from the AIDS Reagent and Reference Program. These isolates were chosen as they were isolated from diverse geographic locations during the acute phase of HIV infection. 36,37
Each plasmid was amplified in Escherichia coli strain-DH5 alpha, purified using anion-exchange resin columns (Qiagen, Valencia, CA), and stored at −20°C in dH2O. Plasmids were verified by appropriate restriction enzyme digestion and gel electrophoresis. The purity of DNA preparations was determined by optical density reading at a wavelength of 260 and 280 nm.
Purification of virus-like particles
Supernatants from COS cells, transiently transfected with plasmid expressing Gag or VLPs, were purified via ultracentrifugation (100,000×g through 20% glycerol, weight per volume) for 4 h at 4°C. The pellets were subsequently resuspended in PBS and stored at −20°C until use. Protein concentration was determined by Micro BCA Protein Assay Reagent Kit (Pierce Biotechnology, Rockford, IL).
Immunization of rhesus macaques
Rhesus macaques, Macaca mulatta, were anesthetized with ketamine. Vaccinations were completed at 8 week intervals over 32 weeks. The first two vaccinations were given as DNA vaccinations. All groups received 1 mg each of DNA encoding the HIV VLP containing Env and SIV Gag VLP in 1 ml saline via an IM injection. All groups received 4 μg of HIV VLP and SIV VLP plasmids via GeneGun (Bio-Rad) over the inguinal lymph nodes. The third and fourth vaccinations were administered as 250 μg each of purified HIV VLP and SIV VLP with CpGs. The vaccines were split with 125 μg given via an intramuscular injection in the quadraceps and the second 125 μg given via the intranasal route. Each CpG ODN:
K3–AT
K123–T
D19–GGTGCAT
Collection of samples
Animals were anesthetized using an intramuscular injection of either ketamine or Telazol. Blood was collected from the femoral vein using the Sarstedt Monovette blood collection system (Sarstedt, Inc., Newton, NC). Peripheral blood mononuclear cells (PBMCs) were collected and purified using ACCUSPIN System-Histopaque-1077 tubes according to the manufacturer's instructions (Sigma-Aldrich, St. Louis, MO). Vaginal washes were collected following anesthetization with Telazol. The vaginal mucosa was washed with 5 ml of sterile PBS. Samples were collected and centrifuged to separate cells and supernatants. Cells and supernatant were taken separately and stored at −80°C until used.
Intravaginal challenge
All animals were challenged intravaginally with SHIVSF162p4, which was obtained from Dr. Nancy Miller (DAIDS, NIAID) and Dr. Ranajit Pal (ABL, Kensington, MD). Briefly all animals were anesthetized using an intramuscular injection of either ketamine or Telazol before virus exposures. All virus exposures were a 1 ml dose of 640 TCID50 with the animal in the supine position, which was maintained for at least 10 min. Viral exposures were completed on days 0, 3, 7, and 10.
Viral load determination
Viral RNA (vRNA) levels were determined by Siemens Diagnostics Clinical Laboratory using the SIV RNA 4.0 bDNA Assay (Siemens Diagnostics, Berkeley, CA). The first 0.5 ml plasma samples were collected and concentrated via centrifugation. The SIV genomic RNA is then captured to a microwell by probes targeted to the pol gene. The preamplifier probes are then added binding to the vRNA. Amplifier probes are then added forming a branched DNA (bDNA) complex. Substrate is added resulting in light emission, which is directly proportional to the amount of vRNA present in the sample. A standard curve is used to determine the concentration of vRNA per sample.
CD4/CD8 cell count
EDTA-treated whole blood was stained using CD3, CD4, and CD8 monoclonal antibodies and analyzed using a BD FACSCalibur Flow Cytometer (BD Biosciences, San Jose, CA). Cell counts were determined using the BD Truecount tubes according to the manufacturer's instructions (BD Biosciences, San Jose, CA).
CD8+ lymphocyte depletion
Rhesus macaques were treated with the antibody cM-T807 starting at day 168 postchallenge (Nonhuman Primate Reagent Resource, Boston, MA). Antibody was given on day 0 at a dose of 10 mg/kg subcutaneously. Additional doses were given at days, 3, 7, and 10 at a dose of 5 mg/kg given via the intravenous route. Intravenous treatments were given as a slow bolus over 10–20 min. CD8 depletion was confirmed by flow cytometry.
Antibody response to VLP immunizations
Serum and mucosal wash samples were individually collected and tested for antibody (IgG) responses to SIV Gag and multiple gp120 isolates by ELISA. Primary Env isolates were obtained from the NIH ARRRP as previously described. 36 SIV Gagp55-purified protein was obtained from the NIH AIDS Research and Reference Reagent Program. Envgp120 was obtained from homologous isolates ConB, AC10, PVO.4, SC42, parental challenge strain SF162, historical late stage isolates IIIB, ADA, YU-2, and early X5 tropic 65353, Qh0692.29, TRO.11, CAAN53542, WITO4160.35, REJO4541, and TRJO4551.54 from supernatants collected from 293T cells transiently transfected with gp120-expressing plasmids. For the Env ELISAs each well of a 96-well plate was pretreated with 5 μg of concanavalin A for 1 h at 25°C. For all ELISAs each well of a 96-well plate was coated with 50 ng per well of the respective protein (4°C for 16 h). Plates were blocked (25°C for 2 h) with PBS containing Tween 20 (0.05%) and nonfat dry milk (5%) and then incubated with serial dilutions of each sample (25°C for 2 h). Following thorough washing in PBS-Tween 20 (0.05%), samples were incubated (25°C for 1 h) with biotinylated goat antimouse IgG conjugated to horseradish peroxidase (HRP) diluted 1:5000 in PBS-Tween 20 (0.05%) and nonfat dry milk (5%). The unbound antibody was removed, and the wells were washed. Samples were incubated with TMB substrate (1 h), and the colorimetric change was measured as the optical density (OD, 405 nm) by a spectrophotometer (BioTek Instruments, Winooski, VT). The OD value of prebleed sera was subtracted from the test samples to adjust for background responses. Results were recorded as the arithmetic mean±the standard deviation.
Neutralization assay
Antisera was tested for the ability to neutralize virus infection in vitro using TZM-Bl indicator cells and pseudotyped HIV-1 SF162. 39,40 These cells express human CD4 (hCD4), human CCR5 (hCCR5), human CXCR4 (hCXCR4), and a luciferase reporter driven by the HIV-1 LTR. TZM-Bl cells were cultured in cDMEM with 10% fetal calf serum (10%) (Atlanta Biologicals, Atlanta, GA). Infectivity was determined using serial dilutions of antisera with cells in complete, nonselective media in the presence of DEAE dextran (20 μg/ml) (25°C for 1 h). Cell lysates were harvested in lysis buffer (25 mM Tris phosphate, pH 7.8, 2 mM DTT, 2 mM 1-2-diaminocyclohexane-N,N,N′,N′-tetraacetic acid, 10% glycerol, 1% Triton X-100) (48 h) and then clarified by centrifugation. Virus neutralization by antisera was determined by measuring the relative light units (RLU) using a Femtomaster FB12 Luminometer (Zylux, Maryville, TN).
Antibody-dependent cell-mediated virus inhibition
Antibody-dependent cell-mediated viral inhibition (ADCVI) antibody activity was measured using methods similar to those described previously. 41 Briefly, target cells (CEM.NKR-CCR5 cells infected with SHIVSF162p4 virus for 48 h) were incubated with serum and with fresh human PBMC effector cells (effector/target ratio=10:1). Seven days later, p27 from the supernatant was determined by ELISA (Zeptometrix Corporation, Buffalo, NY). Percent virus inhibition was calculated relative to negative control sera.
ELISpot assays
The number of anti-Gag and anti-Env-specific NHP interferon (INF)-γ (mINF-γ)-secreting splenocytes was determined by enzyme-linked immunospot (ELISpot) assay (R & D Systems, Minneapolis, MN). Briefly, precoated anti-IFN-γ plates were incubated (25°C for 2 h) with cRPMI (200 μl) and then were incubated with PBMCs (2.5×105/well). PBMCs were stimulated (48 h) with peptides (15mers overlapping by 11 amino acids) representing the SIVMac239 Gag or SF162 Env proteins (NIH ARRRP). Peptide pools were split into 5′ and 3′ pools for both SIVMac239 and SF162 Env. Control wells were stimulated with PMA(+) (50 ng)/ionomycin (500 ng) or were mock stimulated(–). Plates were washed with PBS-Tween (3×) and were incubated (37°C for 24 h; 5% CO2) with biotinylated anti-mIFN-γ and incubated (4°C for 16 h). The plates were washed and incubated (25°C for 2 h) with streptavidin conjugated to alkaline phosphatase. Following extensive washing, cytokine/antibody complexes were incubated (25°C for 1 h) with stable BCIP/NBT chromagen. The plates were rinsed with dH2O and air dried (25°C for 2 h). Spots were counted by an ImmunoSpot ELISpot reader (Cellular Technology Ltd., Cleveland, OH).
Results
The goal of this study was to enhance the breadth of immune responses directed against the HIV-1 envelope. To achieve this goal, a mixture of wild-type (polyvalent) or consensus envelopes was used as immunogens on the surface of a VLP. These vaccines were measured by determining their ability to elicit broadly reactive immune responses and to protect against a heterologous infection. A consensus clade B (ConB) and a polyvalent B (PolyB) VLP vaccine were based on an identical VLP background with the only difference being the envelopes expressed on the surface (Table 1). As previously described, 15 the consensus Env sequences were generated by selecting the most common amino acid at each position of a full-length Env alignment derived from 200 subtype B viruses deposited in the Los Alamos HIV Sequence Database. Each set of subtype sequences was supplemented with env sequences used in a standardized neutralizing panel. 36
Virus-Like Particle Vaccines
Direction of sexual transmission: M-M, male to male; M-F, male to female; F-M female to male.
Length of infection is defined as time between last negative HIV-1 test and first positive HIV-1 test.
The PolyB vaccine is composed of four separate VLPs with each expressing an individual Env isolate. These isolates were selected for their genetic, geographic, and transmission diversity. In addition, isolates were chosen only if they were isolated within a few weeks after transmission, since these early isolates may represent the majority of mucosally transmitted HIV isolates. 42 Vaccinations were administered systemically (intramuscular injection) and mucosally (intranasal delivery) to generate both mucosal and systemic immune responses. Each NHP was identified by a color-coded number within each given study group to facilitate easy comparisons between immune responses and infection outcome (Table 2).
Individual Nonhuman Primates Coding
Cellular immune responses were investigated against both SIV Gagp55 (mac239) and HIV-1 Envgp160 (SF162). Positive cellular responses were not detected either systemically (peripheral blood draw) or mucosally (vaginal wash) from any of the vaccine groups (data not shown). However, both ConB and PolyB VLP vaccines elicited strong SIV Gagp55 antibody responses with a trend toward higher antibody responses induced by the PolyB VLP group compared to the ConB VLP vaccine group (p=0.0676) (Fig. 1A). Limited SIV Gag antibody responses were also detected in vaginal wash samples (Fig. 1B).
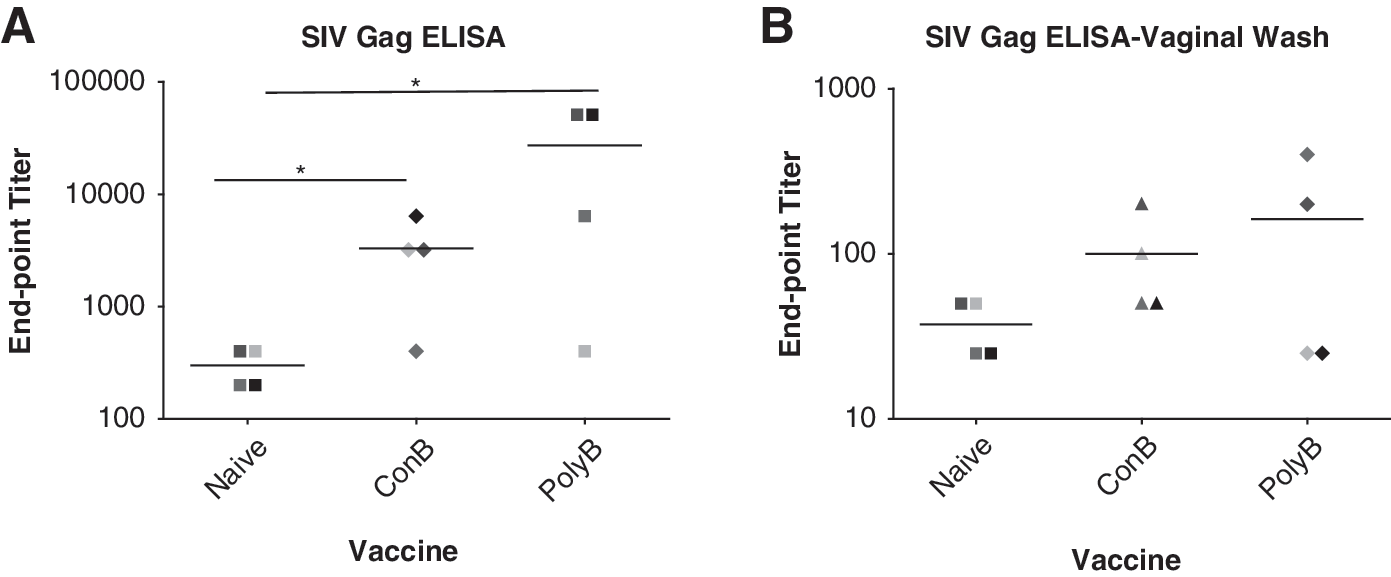
Systemic and mucosal antibody responses are generated by both vaccines. Serum samples and vaginal washes were taken 2 weeks following the completion of the vaccine regimen. Individual serum samples
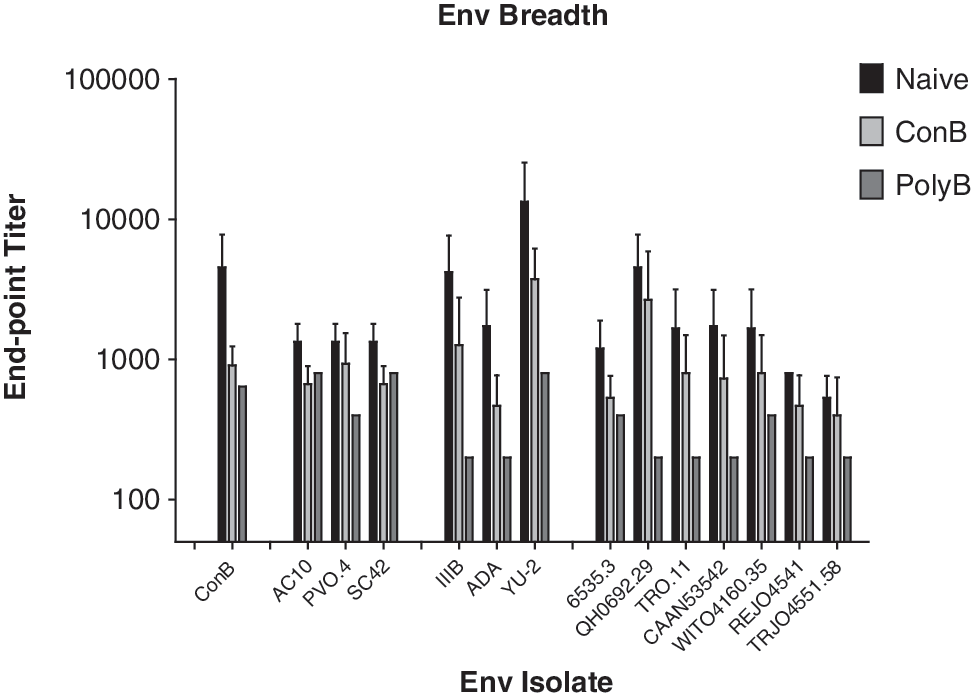
The increased breadth of antibody responses directed against Env was measured following a step-down process utilizing a panel of clade B viral isolates. The first analysis was an increase in anti-Env antibody recognition to a panel of HIV-1 envelopes as measured by ELISA binding (Fig. 2). The PolyB VLP group demonstrated a significantly greater breadth of anti-Env antibody responses than either the unvaccinated or ConB VLP groups (p-value <0.05, Tukey's multiple comparison test). NHPs vaccinated with the PolyB Env VLP vaccine elicited anti-Env antibodies that recognized 12 of the 13 primary Env isolates, including all the isolates obtained from patients soon after infection (Fig. 2). Of particular interest was the response to the SF162 envelope, since this strain matches the challenge virus. Antiserum from one out of four NHPs from the ConB Env VLP group recognized the SF162 Envgp120, while the serum from two out of four of the NHPs from the PolyB Env VLP group recognized the SF162 envelope (Fig. 3). Mucosal antibody responses were not detected against any of the envelopes tested (data not shown). The second level of analysis was to determine enhancement of antibody neutralization activity. Neither vaccine induced neutralizing antibodies against any of the Env isolates tested. The third level of analysis was antibody-dependent cellular virus inhibition (ADCVI) of SHIVSF162p4 virus infection. ADCVI activity was not observed in any of the vaccine groups following the vaccination regiment (data not shown).

Breadth of anti-Env antibody responses. Serum samples obtained 2 weeks following the final vaccination were tested by ELISA via serial dilution using plates coated with homologous (ConB, AC10, PV0.4, SC42) and heterologous (historic controls IIIB, ADA, YU-2; X5 early isolates 65353, Qh0692.29, TRO.11, CAAN53542, WITO4160.35, REJO4541, and TRJO4551.54) gp120. Results are represented as the average response from the four nonhuman primates (NHPs) per group minus background responses shown with error bars representing the standard deviation of the mean. (*p-value <0.05, Tukey's multiple comparisons test.)

SF162 antibody responses. Serum samples obtained 2 weeks following the final vaccination were tested by ELISA via serial dilution using plates coated with parental SF162gp120.
Vaccinated NHPs were challenged vaginally (4×) with SHIVSF162p4 (640 TCID50). 3,43 The envelope in this SHIV does not match any of the envelopes used in the VLP vaccine formulation. The SHIVSF162p4 has an ∼86% amino acid homology with the four primary envelopes used in the PolyB VLP vaccine and an even higher homology with the ConB Env (Table 3). Seventy-five percent of unvaccinated control NHPs was infected with peak viral loads between 4.3×106 and 1.1×108 vRNA copies/ml (Fig. 4A). When normalized for initial day of infection, the peak viremia was observed at ∼14 days postinfection. Similar results were observed with NHPs vaccinated with ConB Env VLPs that had peak viral titers between 5.4×105 and 2.6×107 vRNA copies/ml (Fig. 4B). In contrast, only one NHP (AH69) vaccinated with the PolyB Env VLP vaccine had detectable virus at any point during the infection period (Fig. 4C). The virus titer peaked in this animal at 1.4×105 vRNA copies/ml at day 14 postinfection, but returned to undetectable levels (>165 vRNA copies/ml) between days 54 and 70 postinfection. CD4 levels were not affected for any individual NHP throughout the study regardless of infection status or vaccination group (data not shown). The infection status of all noninfected NHPs did not correlate with any previously described protective MHC alleles. 44,45

PolyB vaccination induced a reduction of viral load and viral burden following heterologous challenge. Plasma was collected at the day of challenge as well as following challenge to measure infection. Viral RNA copies/ml of plasma were determined by bDNA testing. Shown are the viral RNA copies/ml for each individual in the
Envelope Sequence Homology
All noninfected and infected NHPs that had undetectable virus at day 168 postinfection were depleted of CD8 T cells to release low viral titers in reservoir tissues. All NHPs were CD8 depleted via cM-T807 antibody treatment over 10 days. Complete depletion of CD8+ T cells in the blood was confirmed by flow cytometry in all treated NHPs (Fig. 5B, D, and F). All infected NHPs had a rebound virus detected in the blood by day 10 posttreatment (Fig. 5). All noninfected NHPs remained virus free through day 42 posttreatment, further confirming their noninfected status (Fig. 5).
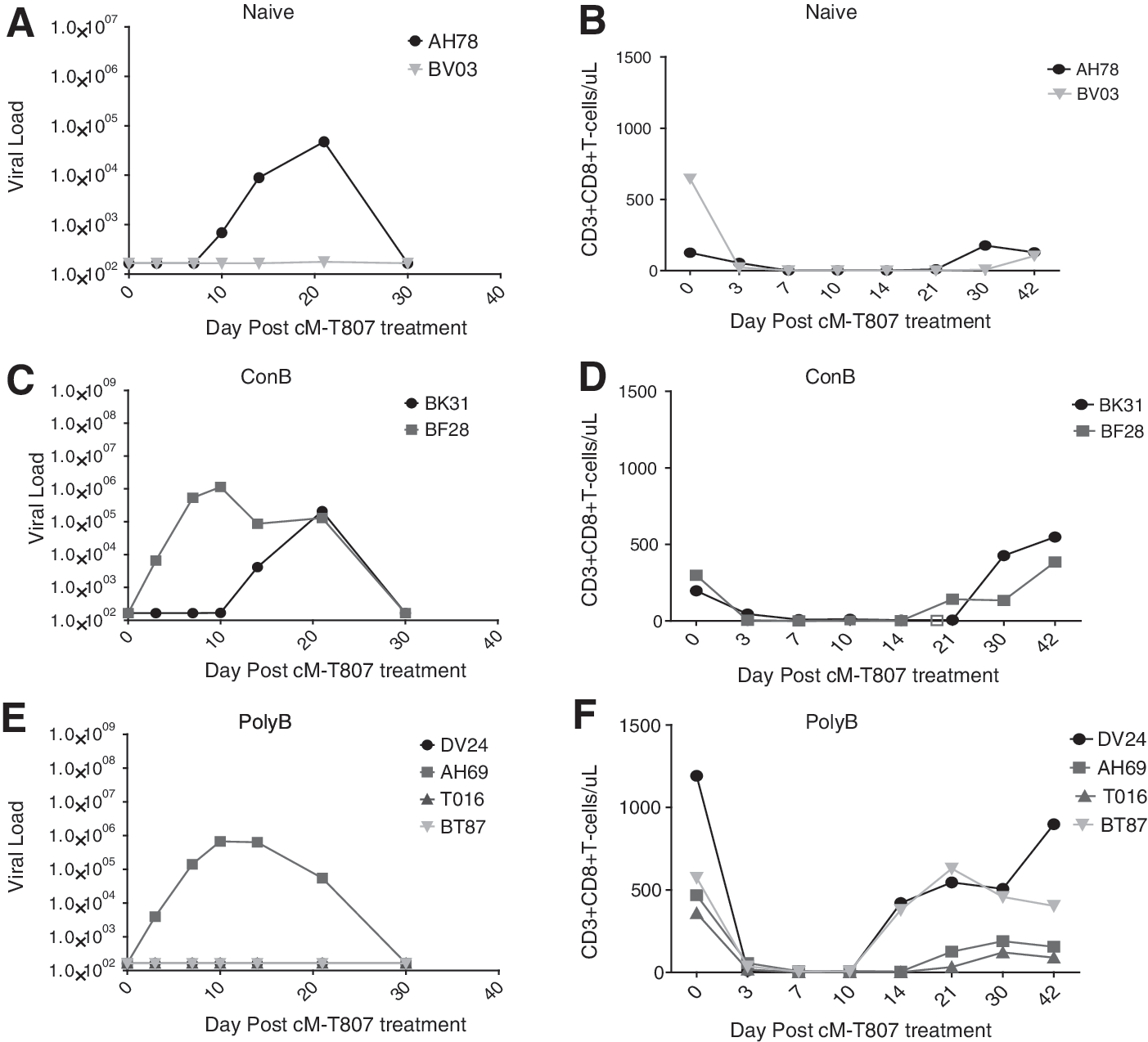
CD8 depletion confirms noninfection status of protected PolyB individuals. At day 168 postinfection all NHP with undetectable viral loads were treated with cM-T807 to deplete CD8 cells. Reported are the viral RNA copies/ml from individual plasma samples as determined by bDNA testing:
Following challenge, there was a marked increase in antibodies to the SF162 Env. There was an initial increase in anti-Env IgG at day 21 that plateaued by day 54 postinfection (Fig. 6A). At day 42, low levels of neutralizing antibodies against the closely related (>99% Env amino acid identity) HIV-1 strain, SF162, were detected. Neutralizing antibody levels continued to increase to day 56 and then were maintained through day 84 postinfection (Fig. 6B). ADCVI activity was also detected by day 84 postinfection (Fig. 6C). The kinetics of acquiring antibody responses were similar between vaccinated and unvaccinated NHPs. The effect of vaccination on the breadth of anti-Env antibody responses postinfection was also investigated using a panel of envelopes. All groups demonstrated an increase in breadth as compared to preinfection (Figs. 7 and 2). All postchallenge immune responses were dependent on the establishment of infection with all anti-Env antibody responses being positively correlated to viral burden (r 2 =0.8478, p-value <0.0001). Postchallenge antibody responses were not detected in any of the noninfected NHPs and were not included in the postinfection antibody analysis (data not shown). There were no cellular responses detected against the SF162 envelope at any point during the challenge. In contrast, SIV Gag cellular responses were detected in all infected NHPs to similar levels though anti-Gag IgG responses remained at prechallenge levels (data not shown).

Viral infection leads to the rapid expansion of binding and neutralizing antibodies directed to SF162. Following viral challenge serum samples were collected at various time points. Individual serum samples were tested by ELISA via serial dilution against SF162 gp120 envelope
Postinfection breadth of anti-Env antibody responses. Serum samples obtained at day 84 postinfection were tested by ELISA via serial dilution using plates coated with homologous (ConB, AC10, PV0.4, SC42) and heterologous (historic controls IIIB, ADA, YU-2; X5 early isolates 65353, Qh0692.29, TRO.11, CAAN53542, WITO4160.35, REJO4541, and TRJO4551.54) gp120. Results are represented as the average response from each group utilizing only those individuals who had detectable vRNA levels at any point during the challenge. Responses are shown with error bars representing the standard deviation of the mean.
Discussion
The vast diversity of HIV-1, particularly within Env, has greatly impeded the development of an HIV-1 vaccine. To addresses the issue of Env diversity, we directly compared the use consensus and polyvalent Env strategies for developing broadly reactive HIV/AIDS vaccines. These vaccines were clade B specific in the intent to develop a vaccine that can protect against interclade isolates and then expand that strategy to address all of Group M. VLPs were used as the delivery mechanism for the ConB and PolyB envelopes. VLPs present envelopes in their native, trimeric structure and also facilitate immune responses to the Gag core. 46 These VLPs incorporate similar amounts of envelope on their surface so any differences in the immune responses generated are due to immunogenic differences and not dosage. 15
Both vaccines were immunogenic with the majority of recipients developing antibody titers against SIV Gag with both ConB VLP and PolyB VLP vaccines eliciting immune responses statistically above background levels (p<0.05, Student's t-test) (Fig. 1). The PolyB VLP vaccine trended toward eliciting stronger anti-Gag antibody responses than the ConB VLP vaccine (p-value=0.0676). Interestingly, these results indicate that the envelopes on the surface of the VLP affect the immunogenicity of the Gag core as both the ConB VLP and PolyB VLP vaccines were identical in this regard. It is currently unclear as to the mechanism of this enhancement, but it is likely linked to the immunogenicity of the envelopes themselves.
To measure the breadth of immune responses elicited by both vaccines, a panel of clade B viral isolates was selected for testing against the vaccine-elicited antibodies. These isolates were selected to represent a general sample of all clade B isolates. These isolates were chosen because they were isolated soon after sexual transmission, which is the route responsible for the majority of worldwide HIV infections. 28,36,41 Statistically significant differences were not found between the ConB VLP and PolyB VLP vaccines for any given single isolate. The PolyB VLP vaccine elicited higher titers and a significantly increase in breadth of envelope reactivity as measured by the Tokey's multiple comparison test. This statistical test takes into account the number of envelopes recognized, as well as the strength of that recognition. Antibodies elicited by the PolyB VLP vaccine may recognize an increased number of viral isolates because this vaccine presents a more diverse set of epitopes compared to the consensus VLP vaccine. A consensus VLP vaccine presents epitopes that are common to a large number of isolates, but the polyvalent VLP vaccine may present these same common epitopes in addition to presenting a number of unique B cell epitopes. Therefore, these unique epitopes may enhance the breadth of Env recognition to include isolates that do not contain the most common epitopes, but do contain the unique epitopes.
To determine the protective effect of ConB VLP and PolyB VLP vaccines, all groups were challenged with multiple vaginal exposures of SHIVSF162p4. This virus is heterologous to the ConB envelope and each envelope in the PolyB VLP vaccine (Table 3). Only one monkey vaccinated with the ConB VLP vaccine had no detectable virus following challenge. Similarly, three of four nonvaccinated monkeys had detectable virus with one monkey being resistant to infection. Two of the monkeys vaccinated with ConB VLP vaccine-infected NHPs had identical viral burdens as the naive group indicating a lack of protection by this vaccine. One of the ConB VLP-vaccinated monkey had a one log reduction in viral load and this animal had the highest anti-Gag antibody response, as well as the only anti-SF162 Env antibody responses within the ConB VLP group, which may account for the lower viral burden.
The PolyB VLP vaccine initiated a potentially protective effect with three out of four monkeys having no detectable virus following challenge (Fig. 4). The one PolyB VLP-vaccinated monkey that became infected had a 2-log reduction in viral titer over the course of infection. Most likely the high levels of anti-Gag and anti-SF162 Env antibody levels induced by the PolyB VLP vaccine were able to induce this protection (Figs. 1 and 2). Interestingly, the incomplete protection demonstrated by monkey #AH69 of the PolyB VLP vaccine group may be due to the lack of elicited anti-Env responses. Monkey #AH69 had no detectable antibody to any of the envelopes in the panel, including SF162. This was the only PolyB VLP-vaccinated monkey not to recognize any of the envelopes tested (data not shown), thus limiting its immune protection to anti-Gag antibodies only.
The ability of mucosal and plasma IgA to inhibit HIV-1 transcytosis across the mucosal epithelium may represent an important mechanism for protection against the sexual acquisition of HIV-1 infection. 47 While virosome-based vaccines that elicit mucosal Env-specific antibodies correlated with protection, 48 no mucosal IgA was detected in vaginal washes from vaccinated monkeys in this study. We also examined ADCVI activity induced by ConB VLP and PolyB VLP vaccines. Both ADCVI and percent ADCC killing prechallenge and postchallenge have been correlated with reduced acute viremia and chronic viremia. 48,49 However, we observed no significant ADCVI activity in monkeys vaccinated with either vaccine.
No prechallenge neutralizing antibodies were detected, but nonneutralizing plasma antibodies have been associated with protection against viral challenge. 50,51 These nonneutralizing antibodies are capable of inhibiting HIV-1 infection and replication in macrophages and immature dendritic cells. 52 These cells are the first cells exposed to the virus during mucosal transmission and play key roles in the early establishment of infection. 53,54 By blocking infection and transmission to these cells, the PolyB VLP vaccine may elicit immune responses that either completely block infection or significantly reduce the inoculating dose. The development of nonneutralizing antibodies to Env as a correlate of protection is supported by recent findings from the RV144 trial where human vaccinees with high antibody titers to the V1/V2 region of Env received protection from vaccination. In addition, vaccinees with low plasma IgA binding to Env received protection from vaccination, but those with no protection responses received less or no protection. 55
Upon the establishment of infection, all vaccinated and unvaccinated monkeys had similar immune responses. The rate and strength of these immune responses were identical across all groups and were observed only in those animals that had measurable vRNA levels. These immune responses were generated in response to the viral infection alone and were not primed or affected by the previous vaccination. All postchallenge immune responses also correlated with viral burden throughout the disease course further indicating that they were dependent on the infection alone. These results indicate that vaccination with either the ConB VLP or PolyB VLP vaccine did not affect postchallenge immune responses.
Importantly, this study demonstrates for the first time in a direct comparison that a polyvalent Env vaccine strategy is capable of eliciting a greater breadth of humoral immunity than a vaccine based on a consensus Env. This greater humoral breadth may lead to an increased ability to protect against a heterologous viral challenge as compared to a consensus vaccine. Due to the small numbers of animals per group and a 75% infection rate within the naive population, protection cannot be statistically demonstrated in this study. Future studies utilizing a 100% infection rate and/or greater n values will be needed to definitively test protection. The observed apparent infection rate within the PolyB vaccine group is likely due to the increased levels of anti-Gag and anti-SF162 Env antibody responses that were generated by the PolyB VLP vaccine. Future work is needed to induce stronger anti-Env immune responses prechallenge. The virus-like particle platform is limited in the amount of Env on its surface. It is likely that priming the immune response by vaccinating with an Env only vaccine followed by boosting with a VLP will facilitate the generation a stronger anti-Env responses that may be enriched for neutralizing antibodies. 3 The current vaccine strategy also did not induce cellular responses (data not shown). Cellular immune responses may not block the initial infection, but they are capable of clearing already infected cells, likely further decreasing the viral burden. To accomplish this either a larger dose of purified VLP or a viral-vector-delivered VLP may be needed. 56 The PolyB VLP vaccine used in this study demonstrates that a polyvalent vaccine is the more effective strategy to generate a broadly reactive humoral immune response.
Footnotes
Acknowledgments
This research was supported by an award from the National Institute of Health/National Institute of Allergy and Infectious Diseases R01AI068507 to T.M.R. and AIDS fellowship Training Award AI065380. The authors thank Meredith Hunter and Preston Marx for their technical assistance and advice. The following peptides and the following reagents were obtained through the AIDS Research and Reference Reagent Program, Division of AIDS, NIAID, NIH: TZM-bl cells (also called JC57BL-13) (#8129), HIV-1ADA (#416), HIVIg (#3957, from NABI and NHLBI).
Author Disclosure Statement
No competing financial interests exist.