
Abstract

E
Comprehensive analyses of HIV-1-specific T cell responses in different cohorts of HIV-1-infected individuals have reported extensive heterogeneity in breadth and magnitude of HIV-1-specific CD8+ T cell responses, even in individuals with uniformly undetectable HIV-1 viral load such as HIV “elite controllers.” 9,10 In this context we hypothesized that elevated Treg frequencies may impact the ex vivo breadth and magnitude of the total HIV-1-specific T cell response, a research question unanswered prior to the current study.
To address this question we measured the frequency of CD4+CD25+FOXP3+ regulatory T cells in peripheral blood mononuclear cells (PBMCs) from 44 untreated HIV-1-infected individuals, including 18 HIV-1 elite controllers [HIV-1 viral load (VL) <50 HIV-RNA copies/ml; median CD4 count 849 cells/μl, IQR: 686–1209)], 21 individuals with chronic untreated HIV-1 infection (median VL 44,500 HIV-RNA copies/ml, IQR: 11,300–69,150; median CD4 count 430 cells/μl, IQR: 353–547), and 5 individuals with untreated acute infection (median VL 416,000 HIV-RNA copies/ml, IQR: 228,500–1,613,000; median CD4 count 480 cells/μl, IQR: 263–487) [gating scheme Fig. 1a, appropriate CD25 and FOXP3 Fluorescence Minus One (FMO) controls were used to define the CD25+FOXP3+ population]. We concomitantly screened these study subjects for HIV-1-specific CD8+ T cell responses using a panel of 222 HIV-1 optimal epitope peptides tailored to the patients' HLA-Class-I type in an IFN-γ-ELISpot assay. For a subset of 17 individuals additional ELISpot data on total T cell responses against the entire expressed HIV-1 genome were generated using a panel of 410 overlapping 15mer peptides spanning all HIV-1 proteins.
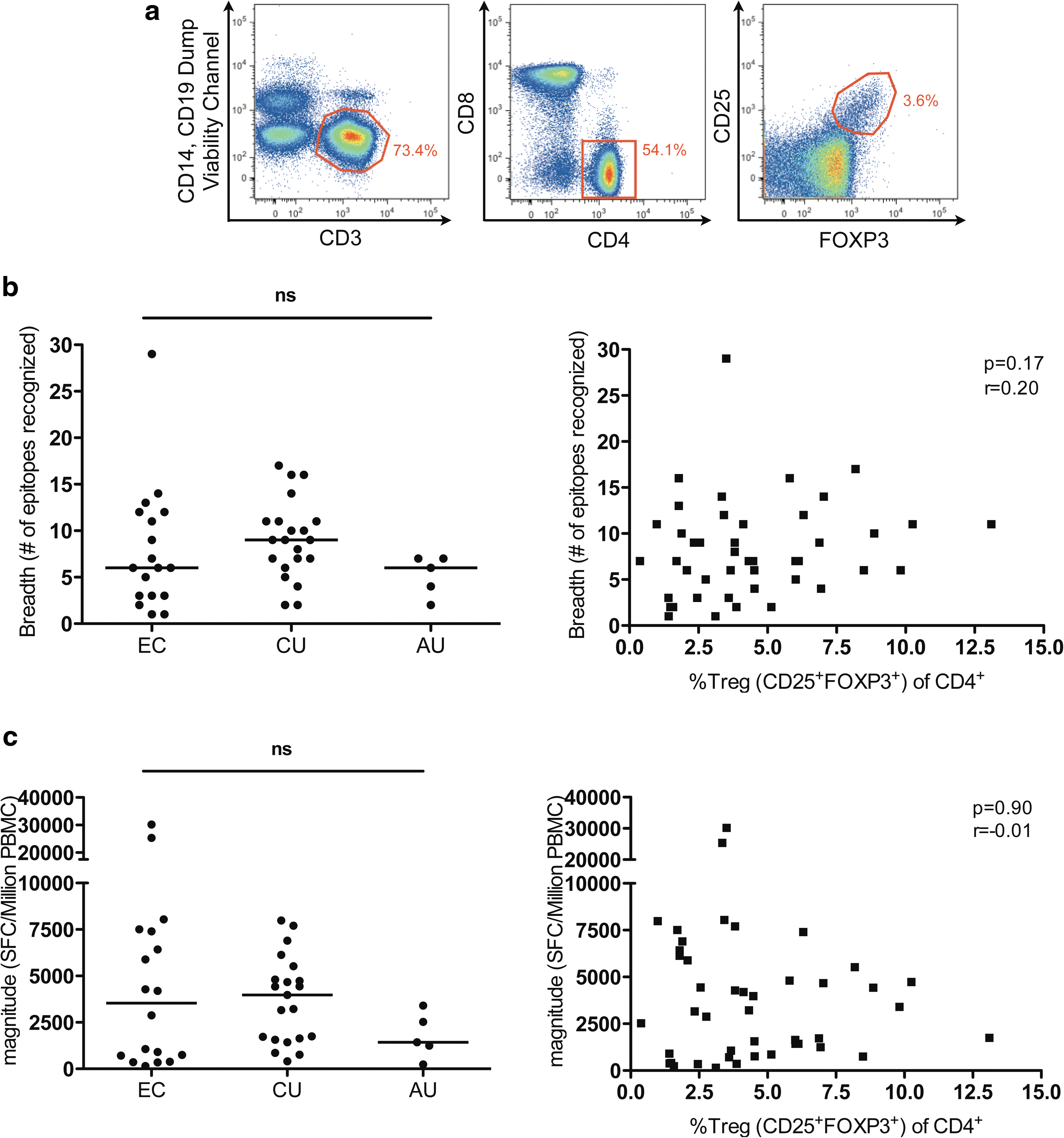
As anticipated, breadth (=number of epitopes recognized) and magnitude (=sum of total number of spot-forming cells/million PBMCs) were lowest in individuals with primary HIV-1 infection. 9 No statistically significant differences in breadth or magnitude were found between elite controllers and chronic progressors (Fig. 1b and c, left panels) and we observed a marked heterogeneity of breadth and magnitude of HIV-1-specific CD8+ T cell responses within the HIV-1 elite controllers, despite uniformly undetectable HIV viral load in line with our previously published data. 10
We next correlated the frequency of CD4+CD25+FOXP3+ Tregs to breadth and magnitude of the HIV-1-specific CD8+ T cell response in the 44 study subjects and found that neither breadth nor magnitude showed an association with Treg frequencies (Fig. 1b and c; right panels) or absolute Treg numbers (r=0.19, p=0.22 and r=0.03, p=0.81, respectively). To account for potential bias toward HLA alleles with many described optimal epitopes, we also generated an optimal epitope recognition score determined by the number of optimal epitopes recognized per individual divided by the number of epitopes described and tested for the respective HLA type. Again, there was no statistically significant difference in optimal epitope recognition scores between the study groups (data not shown) and no correlation with Treg frequency (r=0.18, p=0.24) or absolute Treg counts (r=0.10, p=0.49), respectively. Even in elite controllers differences in Treg numbers or frequency did not explain the heterogeneity in HIV-1-specific cellular immune responses.
Based on recent data suggesting differential susceptibility of HIV-1-specific CD8+ T cell epitopes restricted by “nonprotective” versus “protective” HLA-class I alleles (specifically HLA-B57 and HLA-B27) to Treg suppression, 11 we next investigated if associations between Treg frequencies and epitope-specific responses could be identified in our data set after stratification by expression of HLA-B27/B57 (n=16) or lack thereof (n=28). However, neither breadth nor magnitude correlated with Treg frequencies in either group. We also did not observe overall lower Treg frequencies in individuals with HLA-B57 or HLA-B27, possibly mediated by preferential killing of Tregs by HLA-B27/B57-restricted cytotoxic T lymphocytes, as previously suggested. 11
Lastly, among the 17 individuals comprehensively screened with 410 overlapping peptides for T cell responses against the entire expressed HIV-1 genome no association between Tregs and breadth and magnitude of the total HIV-1-specific T cell response was found.
Taken together these results suggest that frequencies of regulatory T cells in the peripheral blood do not directly correlate with the breadth and magnitude of HIV-1-specific T cell responses as measured by IFN-γ secretion. Our data assessing CD8+ T cell function on the single epitope level by IFN-γ ELISpot confirm and extend previous reports by Owen et al., in which HIV-1-specific T cell immunity was assessed by multiplex cytokine and chemokine analysis in the supernatant of HIV-1-stimulated PBMCs, and similarly no correlation between Treg numbers and HIV-1-induced cytokine production was observed. 12 While cytokine secretion in these two studies did not appear to be impacted by Treg numbers, our data do not rule out that other T cell effector functions such as HIV-1-specific proliferation or cytotoxicity are more closely associated with Treg frequencies. In fact, the differential impact of Treg suppression on proliferation compared to cytokine secretion has been suggested in a recent study. 11
While these observations provide a detailed investigation of Tregs and HIV-1-specific T cell responses in peripheral blood, recent data suggest that mucosal and lymphoid tissues may be more enriched in regulatory T cells. 4,13,14 It is therefore possible that the frequencies of tissue-based Tregs, e.g., in gut-associated or other lymphoid tissue, may have a more direct association with breadth and magnitude of tissue-related HIV-1-specific T cell responses, which warrants further study.
Footnotes
Acknowledgments
We thank all individuals who participated in this study as well as the Ragon Institute Clinical Platform for critical support with cohort coordination and specimen acquisition. This work was supported in part by research funding from the Elisabeth Glaser Pediatric AIDS Foundation (Pediatric HIV Vaccine Program Award MV-00-9-900-1429-0-00 to MMA), MGH/ECOR (Physician Scientist Development Award to M.M.A.), NIH NIAID (PO1 AI074415, KO8 AI074405 and AI074405-03S1 to M.M.A.), and the Doris Duke Charitable Foundation (M.A.). These studies were furthermore supported by the Bill and Melinda Gates Foundation, the Terry and Susan Ragon Foundation, and the Harvard University CFAR (P30AI 060354).
Author Disclosure Statement
No competing financial interests exist.