
Abstract
HLA-B*27:05 is one of the widely reported alleles associated with resistance to HIV, while HLA-A24, HLA-B7, HLA-B*07:02, HLA-B*35:01, HLA-B*53:01, and HLA-B40 are reported to be associated with susceptibility to HIV. Using a bioinformatics approach we attempted to predict potential HLA-B*27:05-specific HIV-1C epitopes that do not bind to susceptibility-associated HLA alleles based on our hypothesis that such epitopes have a greater probability of eliciting a protective immune response in the host. A consensus sequence was built for all proteins of Indian clade C virus. Epitopes specific to HLA-B*27:05 were predicted from the consensus sequence using two different bioinformatics methods to enhance the accuracy of the prediction. Epitopes that were also predicted to bind to any of the susceptibility-associated HLA alleles were excluded from the list. The short-listed epitopes were modeled using MODPROPEP to refine the prediction. Fourteen peptides were identified as epitopes by both sequence-based methods and were found to interact strongly with HLA-B*27:05 by molecular modeling studies. Five of the 14 epitopes were previously reported as immunogenic by other researchers, while the remaining nine are novel. The 14 epitopes have been repeatedly identified by three different methods indicating their potential as useful candidates for an effective HIV vaccine.
Introduction
I
HLA alleles influence the response of the host to the pathogen. HLA-A027 and HLA-A11, 8 specifically HLA-A*11:019 and HLA-B*27, 7,10 –13 particularly HLA-B*27:0514 and HLA-B*57 15 have been reported to be associated with resistance to HIV or slow progression to AIDS. Likewise, HLA-A24, 7 HLA-B7, 14 several alleles of HLA-B35-(HLA-B*35:01, 7 HLA-B*35:02, and HLA-B*35:03), 12 and HLA-B40, 8 specifically HLA-B40:06 9 and HLA-B*53, 12 have been reported to be associated with susceptibility to HIV infection or rapid progression to AIDS. Such differential host genetic associations with infection and disease prompted us to hypothesize that epitopes that could bind specifically to resistance-associated HLA alleles and not susceptibility-associated ones could provoke a robust protective immune response in the host. Such epitopes could form critical components of an effective vaccine against HIV.
HIV-1C is the most common clade responsible for the burden of AIDS globally. 16 Strains of HIV-1C circulating in India are known to be biologically distinct from those in other parts of the world. 17 We therefore chose Indian HIV-1C as a model for the present analysis. Gag and Nef proteins of HIV have been widely reported to stimulate cell-mediated immune responses, 5,6,18 but attempts to identify potential epitopes from the rest of the proteome of Indian HIV-1C isolates are limited. The present study was designed to screen the entire expressed genome of Indian HIV-1C for potential epitopes that bind to HLA-B*27:05, since it is one of the most widely reported HLA alleles associated with resistance to HIV, having experimentally solved crystal structures that could be used for modeling studies.
Materials and Methods
Building of consensus sequence for HIV-1C proteins
A total of 1,769 sequences of all expressed proteins (Gag, Nef, gp120, gp41, p31-integrase, p51RT, protease, rev, tat, vif, vpr, and vpu) of Indian HIV-1C were downloaded from the Los Alamos HIV sequence database 19 (as of June 1, 2010). Multiple sequence alignment was performed using Clustal W2 20 with default parameters. The Consensus Maker from the Los Alamos online resource for HIV 21 was used to construct a consensus sequence for each protein.
Prediction of HLA-B*27:05-specific epitopes
Possible epitopes specific to HLA-B*27:05 were predicted from the consensus sequence of each protein of Indian HIV-1C using the Propred1 22 and IEDB consensus method. 23 The threshold value was set at 3 in the ProPred1 and 5% in the IEDB consensus method. Predicted epitopes that also bind to HIV susceptibility-associated HLA alleles were excluded from further analysis. Those peptides identified as epitopes by both methods were selected for molecular modeling studies.
Molecular modeling of epitopes with HLA-B*27:05
Short-listed epitopes were modeled with HLA-B*27:05 (PDB ID: 1OGT by Hülsmeyer et al. 24 ) using MODPROPEP. 25 MODPROPEP is a web server that helps to model protein-peptide complexes, specifically with MHC proteins and kinases. Residue-based statistical energy function, Miyazawa and Jernigan pair-potential matrix incorporated in the MODPROPEP was chosen for modeling the peptide in the MHC binding pocket. Among the modeled peptides, nonamers that had a better energy score than that found complexed with the HLA-B*27:05 molecule in the crystal structure (1OGT) were considered to be better binders to HLA, and hence probably better immunogens.
Results and Discussion
The diversity of HIV has made it difficult to come up with an effective vaccine for AIDS. Given the fact that strains of HIV-1C from India are distinct from those in other parts of the world, we undertook a comprehensive analysis for immunodominant epitopes in HIV-1C circulating in India. As it has become increasingly evident that region/subtype-specific vaccines are required for HIV, the study aimed to identify suitable candidates for an effective vaccine against Indian HIV-1C. Other investigators have also attempted to identify immunogenic CTL epitopes from Gag and Nef proteins of HIV-1C. 5,6,18 The epitopes identified by them were longer than nine amino acids. While it is thought that class I MHC are known to bind to peptides 8–12 amino acids in length, 26 most of the minimal epitopes reported are 9-mers. 27 We therefore attempted to predict minimal epitopes from all proteins of Indian HIV-1 subtype C, and have identified highly potential candidates using in silico methods.
Mapping of epitopes in pathogens in vitro is a time-consuming and expensive procedure, and bioinformatics tools can be effectively employed to hand pick a set of highly potential ones that can be further tested using in vitro and in vivo models. Propred1 is one of the widely used programs for prediction of HLA class I-specific epitopes, having a sensitivity of 66–78% and specificity of 80–81%. 22 This tool was initially employed to predict epitopes that specifically bind to HLA-B27:05. As usage of more than one predictive method simultaneously is known to increase the accuracy of the results, 28 we used an additional bioinformatics tool (the IEDB consensus method) 23 to identify epitopes. This tool is a consensus algorithm for the prediction of CTL epitopes based on four different methods, and it is reported to be superior to any of the individual methods on which it is based. 29 Epitopes that were predicted by both methods were further subjected to molecular modeling.
HLA-B*27:05-specific epitopes
HLA-B*27 is reported to be associated with resistance to HIV or slow progression of AIDS in HIV-infected individuals by several investigators, 7,10 –14 and cytotoxic T lymphocyte responses, activated by HLA-B27, are implicated in playing a protective role against HIV infection. 30 The frequency of the HLA-B27 allele in the Indian population varies widely between 1% and 29% across different regions. 31 Among the HLA-B*27 alleles, HLA-B*27:05 is one of the most characterized and its three-dimensional structure is available 24 for molecular modeling studies. Association of HLA-B*27:05 with resistance to HIV implies that epitopes that bind specifically and strongly to the HLA-B*27:05 molecule could provoke a robust protective response in the host.
Two hundred and eighty-eight nonamers were predicted as potential binders to HLA-B*27:05. Of these, 92 nonamers were also predicted to bind to HLA molecules that are known to be associated with susceptibility to HIV infection or rapid progression to AIDS. These nonamers were excluded and the remaining 196 nonamers were identified as epitopes specific to HLA-B27:05 by the first method, viz. ProPred1. Furthermore, 96 of these 196 epitopes were also found to be HLA-B*27:05-specific in the second round of analysis using the IEDB consensus method. 23 These were taken up for further analysis. They were modeled onto the peptide binding site of HLA-B*27:05 using the three-dimensional structure available in the Protein Data Bank (PDB). The binding energy of the nonamer bound to the HLA allele present in the PDB was taken as the threshold for short-listing the predicted nonamers as better binders to HLA-B*27:05, and therefore highly probable immunogens. Fourteen nonamers scored better than the control nonamer. These data are summarized in Table 1. Envelope proteins were found to contain a maximum number of epitopes (five from gp41 and two from gp120), followed by gag and protease (two epitopes each), and one each from RT, vpu, and vpr. The details of the short-listed epitopes are given in Table 2.
The MJ energy score of the bound HLA-B*27:05 peptide was −39.01 kcal/mol. Preferred amino acids in anchor sites (P2 and P9) are underlined. Epitopes with preferred amino acids in P2 and P9 and also reported previously to elicit CD4 and/or CD8 responses are given in bold. Epitopes with preferred amino acids in both P2 and P9 are given in italic.
Epitoes conserved identically in more than 50% of the total sequences (58.46% to 91.67%).
Epitoes conserved in more than 50% of the total sequences (56.90% to 83.23%) with one amino acid difference.
We further examined if any of the short-listed epitopes bind to HLA molecules other than those associated with susceptibility to HIV. We found that each of the short-listed HLA-B*27:05-specific epitopes bound to one or more (up to a maximum of eight) HLA alleles (Table 2). The findings that the 14 short-listed epitopes bind strongly to HLA-B*27:05 but not to any of the known susceptibility-associated alleles, and that they interact promiscuously with other HLA types, prompt us to speculate that these epitopes could be highly significant contributions for the design of a CTL-targeted vaccine against the Indian HIV-1C strains.
Conservation of epitopes
HIV is known to evolve by undergoing escape mutations in CD8+ T cell epitopes.
32
CTL escape mutants transmit from one host to another,
33
but are likely to revert to the wild type in individuals with HLA mismatch.
34
Emergence of escape mutations is determined by various factors, viral as well as human, and is thus a slow process.
33
Experimentally defined HIV CTL epitopes are actually concentrated in relatively conserved and potentially important functional regions of the genome
35,36
and it is believed that the emergence of escape mutations may exact a cost on viral fitness.
37
–40
Conservation of the short-listed epitopes was examined by comparing the sequences of the predicted epitopes with the corresponding sequence in an infectious HIV-1C clone from India.
41
The amino acid sequences of 12 of the 14 epitopes were found to be conserved in the infectious clone. One epitope from gp120 (1:MRVRGILRN) varied from the primary infectious clone by one amino acid at the sixth position, and another epitope from gp41 (260:LRDFILVTA) varied by two amino acids at positions 6 and 8. We also examined the extent of conservation of the short-listed epitopes among Indian HIV-1C protein sequences employed to develop the consensus sequence using the “epitope conservancy analysis” tool available from
Screening of host mimics
It is reported that epitopes that mimic human peptides are tolerated by our immune system while peptides that do not resemble human peptides induce an immunogenic response in the human host. 43 Further, Maksyutov et al. 44 suggested that epitopes shared by HIV and the human host are likely to be implicated in the immunopathogenesis of AIDS through induction of cross-reacting effectors of the immune system or would cause autoimmune reactions. An inverse relationship has been reported between HIV epitope's similarity with human proteins and their immunogenicity. 45 Thus, the human immune system preferentially responds to antigenic sequences that are different from human peptides, and hence removal of host-mimics from among the list of epitopes could be crucial while designing an efficacious vaccine. The short-listed epitopes were compared with the human proteome (Homo sapiens, taxid: 9606) using BLASTP (with parameters automatically adjusted for short sequence search) 46 to look for the presence of identical peptides. None of the 14 short-listed epitopes were found to be maximally conserved in the human peptides. We extended our analysis to evaluate whether peptides sharing eight or less numbers of residues exist in humans. However, none of epitopes was observed to have similar host peptides sharing eight residues. Five of these epitopes differed by two amino acids, and two epitopes differed by three amino acids. Epitopes of HIV differing by a minimum of two residues from human peptides have been previously demonstrated to stimulate immune response against HIV. 45 The uniqueness of these epitopes implies that they are likely to be recognized by the human immune system as foreign, resulting in the induction of an immunogenic response in man.
Epitopes already reported by others
The set of short-listed epitopes was cross-checked with the HIV Molecular Immunology Database
47
to see if any of them were already reported by others. We found two (Gag271:VRMYSPVSI and protease 56:VRQYDQIPI
Interaction profile of HLA-B*27:05 and potential epitopes
By a comprehensive analysis of all known natural ligands of HLA-B27 and their amino acid composition, Lopez de Castro et al. 49 found that a large number of epitopes contained arginine at position two (P2), and a basic, aliphatic, or aromatic amino acid in position 9 (P9). In general, amino acids in position 2 and the extreme C-terminus of the epitope are generally reported to be critical for binding to most class I MHCs and these positions are referred to as anchor positions, 50 while amino acids in the other positions function as auxiliary anchor residues. 48 We analyzed the binding interactions of 14 short-listed epitopes with the HLA-B*27:05 molecule. Interestingly, we observed that 13 of the 14 epitopes had amino acid arginine at position 2. This residue was found to interact with the maximum number (five) of residues in the HLA molecule (Fig. 1b and c). To investigate the binding efficiency of the epitopes in the binding groove of HLA-B*27:05, a representative solid ribbon model and electrostatic surface model were employed (Fig. 1a and d). Our observations support the fact that arginine at position two is critical for the interaction of the epitope with HLA-B*27:05, as previously reported by other investigators. 49,51,52 Stewart Jones et al. 52 demonstrated that mutation in arginine at P2 eliminates binding of the epitope with HLA-B27, resulting in immune escape and progression to AIDS. We also observed that 10 of the 14 short-listed epitopes had either a basic, aliphatic, or aromatic amino acid in the C-terminus (P9). The amino acid at P9 interacts with two amino acids in the HLA molecule, namely LYS146 and ASP 77 (Fig. 1b and c). Collectively, eight epitopes were observed to have arginine in P2 and either a basic, aliphatic, or aromatic amino acid in P9, highlighting their significance as immunogenic CTL epitopes in HIV.
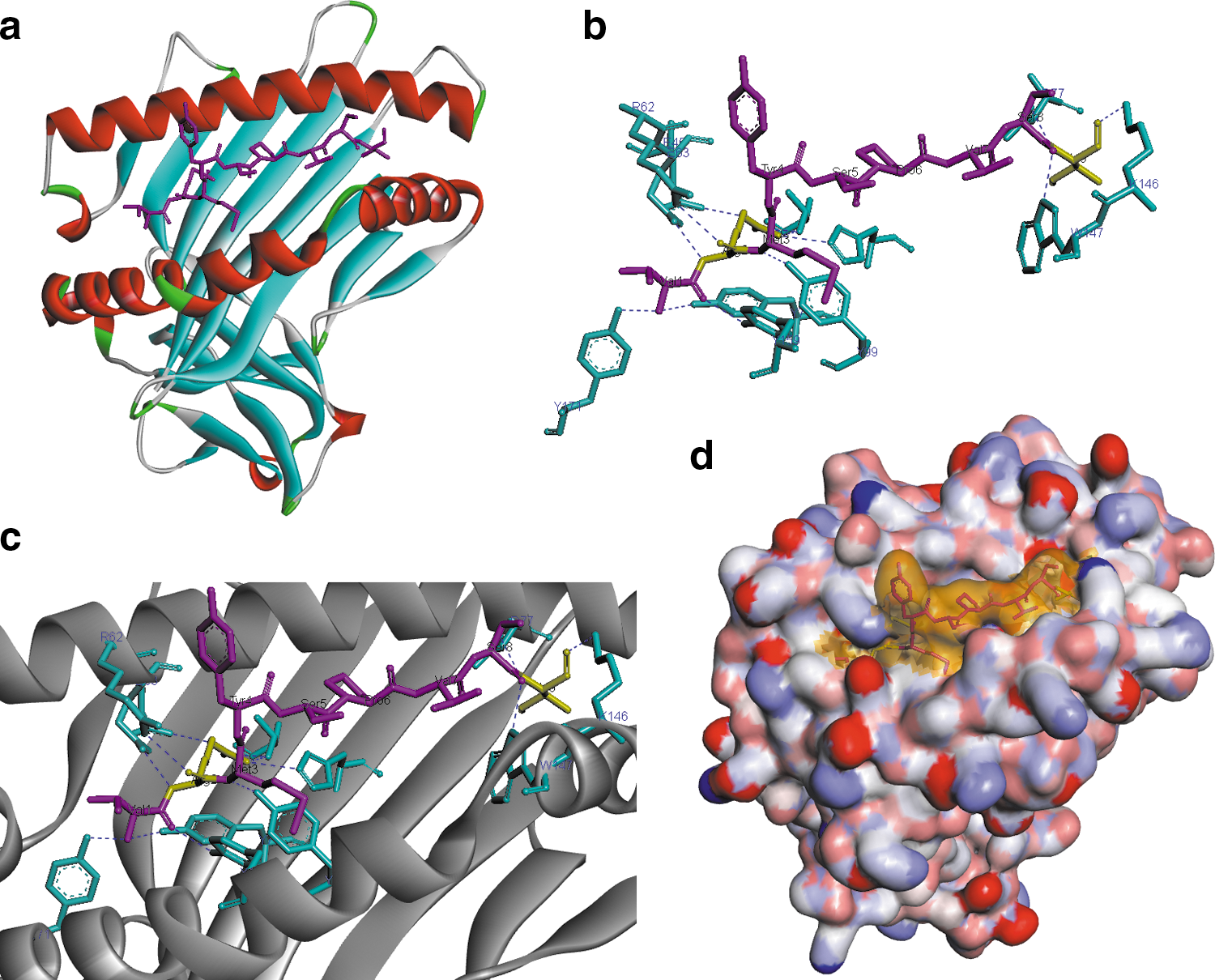
A representative epitope (Gag 271:VRMYSPVSI) modeled onto HLA-B*27:05 using MODPROPEP.
Further analysis revealed that two epitopes, one from Gag and one from protease (Gag271:VRMYSPVSI and Prot56:VRQYDQIPI), that were identified as potentially immunogenic epitopes not only by us but by other investigators as well, 47 also contain the preferred anchor residues at P2 and P9 that hold the binding specificity for HLA-B27. They are also reported to be polyfunctional in nature, and activate both CD4 and CD8 T cells. Furthermore, these two epitopes are present in at least the two major subtypes of HIV-1 (subtypes B and C). 47 Thus, the above mentioned epitopes may be considered highly significant candidates that warrant further investigation.
Genetic diversity in HIV has been the greatest stumbling block to the development of a successful vaccine. It has become evident that the strategy for effective vaccine development should aim at targeting individual subtypes or subtypes that are genetically closely related. Since host genetics plays a vital role in resistance or susceptibility to diseases, it would be significant to consider resistance-associated factors while designing a protective vaccine for HIV. In this scenario, the set of short-listed epitopes targeting HLA-B*27:05 would prove to be useful components for an effective vaccine for HIV-1 subtype C strains in India. However, for the vaccine to be useful for all individuals in a given population, other epitopes capable of inducing a protective immune response in those with other HLA alleles also need to be included. The strategy employed in this study can be used to identify epitopes specific to other HLA alleles associated with resistance to the pathogen. For the benefit of individuals who have susceptibility-associated HLA alleles, epitopes specific to the other HLA alleles present in such individuals should be identified.
Footnotes
Acknowledgments
We acknowledge the Indian Council of Medical Research, New Delhi, India for providing the Bioinformatics facility to our institute.
Author Disclosure Statement
No competing financial interests exist.