
Abstract
This study compared HIV-1 genotypes shed over time (≤3.5 years) in the vaginal secretions (VS) and blood plasma (BP) of 15 chronically infected women. Analysis of predicted coreceptor tropism (CCR5=R5, CXCR4=X4) for quasispecies shedding revealed three patterns: (1) viral quasispecies shed in both VS and BP were restricted to R5-tropism at all time points, (2) quasispecies shed in VS were restricted to R5-tropism at all time points but X4 quasispecies were identified in the BP at one or more time points, and (3) quasispecies shed in matched VS and BP both contained X4-tropic viruses. Overall, the frequency of X4 quasispecies circulation in VS was 2-fold less than in BP and detection of X4 virus in VS was more likely to occur when X4 quasispecies comprised more than 50% of BP viruses (p=0.01) and when declines in blood CD4+ lymphocyte levels were the greatest (p=0.038). Additionally, the mean number of predicted N-glycosylation sites between matched VS and BP samples was strongly correlated (r=0.86, p<0.0001) with glycosylation densities in the following order (VS R5=BP R5 > BP X4 > VS X4). The X4 glycosylation densities may result from compartmentalization pressures in the female genital tract or the delayed appearance of these viruses in VS. Our results suggest that the presence of X4 virus in VS is associated with a threshold population of X4 quasispecies in BP, which are increasing during the HIV-induced failure of the human immune system.
Introduction
T
In more than 95% of recently HIV-1-infected individuals, only non-syncytium-inducing (NSI) variants, which typically use the CCR5 (R5) protein to enter CD4+ cells, are detected. 21 However, during disease progression approximately half of HIV-1 subtype B-infected individuals show the emergence of syncytium-inducing (SI) viruses, which primarily utilize the CXCR4 (X4) transmembrane protein as a coreceptor for cell entry. 21,22 Data from both genotypic and phenotypic studies suggest that NSI/R5 variants are more readily transmitted than SI/X4 strains. 23,24 A number of hypotheses have been offered to explain why SI/X4 viruses are not as readily transmitted via mucosal routes including that cotransmitted SI/X4 variants are more readily cleared by the host immune system, 21 or that selective expression of the R5 cell-surface protein on target cells in the genital mucosa favors the transmission of R5-tropic NSI viruses. 25 Little information has been reported regarding viruses in the sites relevant to most HIV-1 exposures worldwide, the genital tracts of infected men and women, 26 –29 where an increase in the relative abundance of NSI/R5-tropic viruses could indeed facilitate selective transmission of these variants. A cross-sectional genotyping study of a dozen women reported significant differences in the relative abundance of R5- and X4-tropic viruses between blood and vaginal secretions of some women, and X4-tropic viruses were detected only in the blood compartment for a majority of these women, although no longitudinal data were available. 14
The emergence of SI/X4 viruses in BP has been linked to a rapid loss of CD4+ cells and a faster progression to AIDS. 29 Studies have also shown that the appearance of SI/X4 viruses in peripheral blood has a transient nature. Specifically, data from a longitudinal study of nine males with slow or moderate disease progression indicated that SI/X4 viruses appeared in the blood during an intermediate disease stage and peaked just prior to transition into a late stage characterized by a decline of CD4+ cell counts to levels ≤200 cells/μl. 30 Previous studies have not specifically addressed the longitudinal presence or absence of SI/X4 viruses in the female genital tract, or compared a woman's immune status with the appearance of SI/X4 viruses within the vaginal compartment. Therefore, in this study we examined HIV-1 envelope sequences of cell-free viruses in vaginal secretions and blood collected longitudinally from 15 HIV-1-infected women to compare predicted coreceptor phenotypes of viruses shed in these two anatomical sites over time.
Materials and Methods
Study population
This analysis includes data from examinations of women enrolled in the Emory Vaginal Ecology (EVE) Study of HIV-1 infection. HIV-1-infected women were eligible to enroll in this study if they were 18–49 years of age, had a normal Pap smear within the previous 12 months, were expected to live at least 1 year, and if on antiretroviral therapy (ART), had been using the same antiretroviral(s) for at least 3 months before enrollment. Exclusion criteria were as described elsewhere. 31 Participants were requested to refrain from vaginal intercourse and the use of intravaginal medications for 72 h before their examinations.
This subpopulation of samples was chosen from the EVE study using the specific criterion that significant virus shedding was occurring in both blood and the genital tract at all three time points. This was done in order to detect and accurately analyze as many minor quasispecies in the genital tract as possible using our protocol. ART therapy at the time of the EVE study was still dual therapy in some early cases and only later was triple therapy in use. In addition, we have no data on ART adherence or ART failure due to resistance in the EVE study. Thus, the high level of virus shedding observed in this subpopulation of samples was likely due to women who were not on ART, suboptimal dual therapy ART, and possible nonadherence or failure of ART.
Specimens
Matched vaginal secretions (VS) and blood plasma (BP) were collected during clinical examinations at approximately ≥6-month intervals from 15 women enrolled in the EVE study. VS samples were obtained by introducing 10 ml of phosphate-buffered saline (PBS) into the vagina and collecting the pooled fluid in the posterior vaginal fornix. VS samples were tested for the presence of seminal fluid using an acid-phosphatase assay 31,32 and blood contamination was estimated by a method previously described. 31 Endocervical swabs were obtained to culture Neisseria gonorrhoeae and to test for Chlamydia trachomatis. Wet mounts were examined to detect Trichomonas vaginalis. Women subsequently diagnosed with genital tract infections were treated for those infections. Venous blood was collected at each examination in a CPT vacutainer tube that contained acid-citrate-dextrose anticoagulant. Samples were processed by previously described methods. 31 All VS and BP samples used in this study had detectable virus loads determined by a quantitative assay for cell-free HIV-1 RNA described elsewhere. 31,33
Cell-free HIV-1 RNA isolation, reverse transcription, and nested PCR
Cell-free HIV-1 was pelleted from 1-ml aliquots of cell-free VS and BP samples by centrifugation (105×g) for 30 min. RNA was extracted from the pellets using the E.Z.N.A. Total RNA kit (Omega Bio-tek, Doraville, GA) and eluted in 50 μl of water according to the manufacturer's protocol. RNA extractions of VS and BP samples were performed at separate times to prevent the possibility of cross-contamination of matched samples during this procedure.
One-tenth of the extracted RNA (5 μl) was reverse-transcribed for 30 min at 42°C using the GeneAmp RNA PCR kit (Perkin Elmer, Branchburg, NJ) and 50 pmol of an antisense primer (JH35R: 5′-CACTTCTCCAATTGTCCITCA-3′). 34 The resulting complementary DNA was used in an nPCR protocol that amplifies a 540-bp fragment in the C2V4 region of the gp120 envelope gene (7002–7541 of HXB2). 34 The broadly reactive primer sets (JH35R, JH44F: 5′-ACAGTRCARTGYACACATGG-3′, JH33F: 5′-CTGTTIAATGGCAGICTAGC-3′, JH48R: 5′-RATGGGAGGRGYATACAT-3′) and nPCR cycling conditions were previously reported to amplify the C2V4 region from HIV-1 subtypes B, C, D, and E. 34 Reverse transcription and nPCR amplification of VS and BP samples were performed at separate times to prevent the possibility of cross-contamination of matched samples during this procedure.
Three independent RT-nPCRs were performed and pooled for each VS and BP sample in order to obtain a more accurate representation of the cell-free HIV-1 RNA quasispecies. In addition, amplifiable copies of C2V4 template RNA were estimated from the highest dilution of the RNA extract (100 to 103) that produced a 540-bp amplicon detectable by ethidium bromide staining of a 1% agarose gel in the RT-nPCR reactions described above. Negative control RT-nPCR reactions were performed with water as an input template.
Cloning and sequencing of HIV-1 env RT-nPCR products
For each VS and BP sample, the pooled HIV-1 env C2V4 RT-nPCR products were gel purified and then cloned using the TOPO TA kit (Invitrogen, Carlsbad, CA) according to the manufacturer's protocol. Plasmid DNA was isolated from 10 clones for each sample using the QIAprep Spin Miniprep kit (Qiagen, Valencia, CA), and HIV-1 env C2V4 inserts were sequenced using Visible Genetics' OpenGene system. Sequence data were edited with the Sequencher 3.0 program (Gene Codes Corporation, Ann Arbor, MI), and an ∼345-bp fragment that included a portion of the C2 region, and the entire V3 and C3 regions (C2V3C3) was analyzed. For each sequence, the predicted coreceptor tropism of the V3 loop was determined using the PSSM program sinsi matrix (available at
Statistical analyses
Statistical analyses were performed using the Instat 3.0 software package (GraphPad Software, Inc, San Diego, CA). A p value ≤ 0.05 was considered significant. Observed distributions were examined by chi square tests. Mann–Whitney and t tests were used to compare the number of N-glycosylation sites between VS and BP sequences. The proportions of predicted R5 viruses in VS and BP samples were compared in binomial proportions tests. Spearman rank correlation coefficients were calculated to assess relationships between clinical data (e.g., CD4+ cell levels and viral loads) and predicted coreceptor distributions and N-glycosylation densities for VS and BP samples. Mann–Whitney tests and Kruskal–Wallis tests with Dunn's multiple comparisons post tests were used to compare clinical data among patient groups defined by experimental findings (e.g., predicted R5 and X4 viruses, number of N-glycosylation sites).
Nucleotide sequence accession numbers
The V3 nucleotide sequences reported in this study have been assigned GenBank accession numbers HQ822532–HQ 823431.
Results
Study participants
For this analysis we examined a total of 900 HIV-1 C2V3C3 env sequences from matched samples of VS and BP provided at 45 clinic visits of 15 HIV-1-infected women. For each woman, matched samples collected at three clinic visits were selected that spanned a total of 1.5–3.5 years. Fifteen of 30 (50%) intervals between clinic visits were 1 year with 4, 7, 3, and 1 intervals at 0.5, 1.5, 2.0, and 2.5 years, respectively (rounded to nearest half year increment; all intervals ≥6 months, Table 1). A neighbor-joining phylogenetic tree constructed from all 900 HIV-1 env C2V3C3 sequences showed that sequences from the VS and BP samples of each patient clustered together without interpatient contamination (data not shown).
CD4+
Time interval from first sampling time used in these studies (rounded to closet half year increment; all intervals between successive clinic visits ≥6 months.
HIV-1 RNA copies/ml BP.
Total HIV-1 RNA copies in cell-free VS sample.
λ=proportion of sequences from each sample predicted to use the R5 coreceptor.
Mean (SD) number of potential N-glycosylation sites in 10 sequences from each sample.
Significant difference in N-glycosylation densities between sequences obtained at indicated sampling date and those obtained at the following sampling time (Mann–Whitney tests, p≤0.05).
Significant difference in proportion of R5 and X4-tropic viruses or N-glycosylation densities between matched BP and VS samples (binomial proportions and Mann–Whitney tests, respectively, p≤0.05).
Proportion of sequences from each BP and VS sample predicted to use the R5 coreceptor (λ) and N-glycosylation densities for BP and VS sequences also reported.
BP, blood plasma; VS, vaginal secretions; ART, antiretroviral therapy; GT, genital tract; NA, not available.
Each VS and BP sample had a measurable HIV-1 gag RNA virus load that was determined by a QC RT-PCR assay, as described previously. 31 The median virus load for VS and BP samples was 6.0×104 total copies/VS (range, 1.1×103 to 1.4×107 total copies/VS) and 8.7×104 copies/ml (range, 8.3×102 to 1.6×106 copies/ml, Table 1), respectively. The contribution of whole blood to vaginal virus load did not exceed 2% in any of the VS samples. At the first collection date, five, four, and four women had peripheral blood CD4+ cell counts >400 (high), 400–200 (medium), and <200 (low) cells/μl, respectively (Table 1). CD4+ cell count data were not obtained for two women on the first collection date, although both women had high CD4+ cell counts at the time of enrollment in the study. Yearly peripheral blood CD4+ changes ranged from +68 to −139 cells/μl/year (median=–62 cells/μl/year), and by study end, one women had CD4+ cell counts in the high category, while six and seven women had CD4+ levels in the middle and low categories, respectively (the CD4+ cell count for the last collection time was not available for one woman). Four women were receiving antiretroviral therapy during all three clinic visits and another six women were receiving antiretroviral therapy at one or two clinic visits. Trichomonas vaginalis was detected at a single clinic visit of two women, and Chlamydia trachomatis was detected at one clinic visit of another woman (Table 1). Neither Neisseria gonorrhoeae nor seminal fluid was detected at any of the sampling times.
HIV-1 R5- and X4-tropic virus shedding in VS and BP
For each of the 10 clones sequenced from 45 matched VS and BP samples, HIV-1 coreceptor tropism was predicted using the PSSM program. Each matched set of VS and BP samples was categorized as being composed of only R5-tropic viruses (23/45), X4-tropic viruses in plasma only (12/45), or X4-tropic viruses in the VS and BP (10/45). Less than one-half (10/22) of the matched samples where X4-tropic viruses were identified in BP had X4-tropic viruses in the matched VS sample. X4-tropic strains were not detected in VS at any collection time without also being detected in BP.
Longitudinal analysis of the predicted coreceptor phenotypes shed in VS and BP of the 15 women showed three distinct patterns over time. In one pattern, only predicted R5 viruses were shed in both VS and BP for all clinic visits of five women (Fig. 1A). In a second pattern, predicted X4 strains occurred only in BP of five women as a mixture with R5-using viruses at one or more sampling times (Fig. 1B). In the last pattern, predicted X4 viruses were found (alone or as a mixture with R5 strains) in both VS and BP samples of five women (Fig. 1C).

Schematic showing the relative abundance of predicted R5 and X4 viruses being shed in vaginal secretions (VS) and blood plasma (BP) samples over time for each woman. The top and bottom halves of each bar (separated by a dashed white line) represent VS and BP viruses, respectively, and solid and patterned bars indicate predicted R5 and X4 coreceptor usage, respectively. Women are grouped as those who had
The above analysis of coreceptor detection at clinic visits suggested that the presence of X4-topic virus in VS was associated with concurrent circulation in BP. Relative amounts of R5- and X4-tropic viruses were estimated from the sequences of the 10 clones from each VS and BP sample and are expressed by the proportion of R5 virus in a sample (λ=number of predicted R5-tropic clones/10 total clones; λ=1 and 0 indicate all R5 and all X4-tropic viruses in a sample, respectively). Further analysis revealed a stochastic association between levels of X4 virus circulating in blood and detection in VS. Namely, X4 virus detection in VS was more likely to occur when X4 viruses comprised more than 50% of BP viruses (λ<0.5, p=0.01) and even more so when they comprised more than 80% of BP viruses (λ<0.2, p=0.002).
Immune status and HIV-1 coreceptor usage of viruses shed in VS and BP
Significant differences in yearly peripheral blood CD4+ decreases were found between women who shed exclusively R5 viruses and those that shed X4 viruses in BP only or in both VS and BP. For this analysis, yearly decreases in peripheral blood CD4+ cell levels (determined from all available data including data from clinic visits prior to or between the three collection times used in this study) were compared between women who shed exclusively R5 viruses in both VS and BP (Fig. 1A) and those who shed X4 at any collection time (Fig. 1B and C). Yearly peripheral blood CD4+ decreases were greater for women who shed predicted X4 viruses at any collection time (median loss=–2 and −79 CD4+ cells/μl/year, for women with R5 only and X4 at any time, respectively, p=0.038, Fig. 2). Differences between the starting blood CD4+ cell levels for these two groups were not statistically significant. This analysis of blood CD4+ cell level decreases excluded the two women noted above (patient ID 35 and 50) who shed predominantly X4 viruses in their VS and BP samples, but also had very low CD4+ cell levels (≤40 cells/μl) for all their clinic visits. Viral loads and antiretroviral therapy were not associated with the shedding of X4 viruses in VS or BP samples.
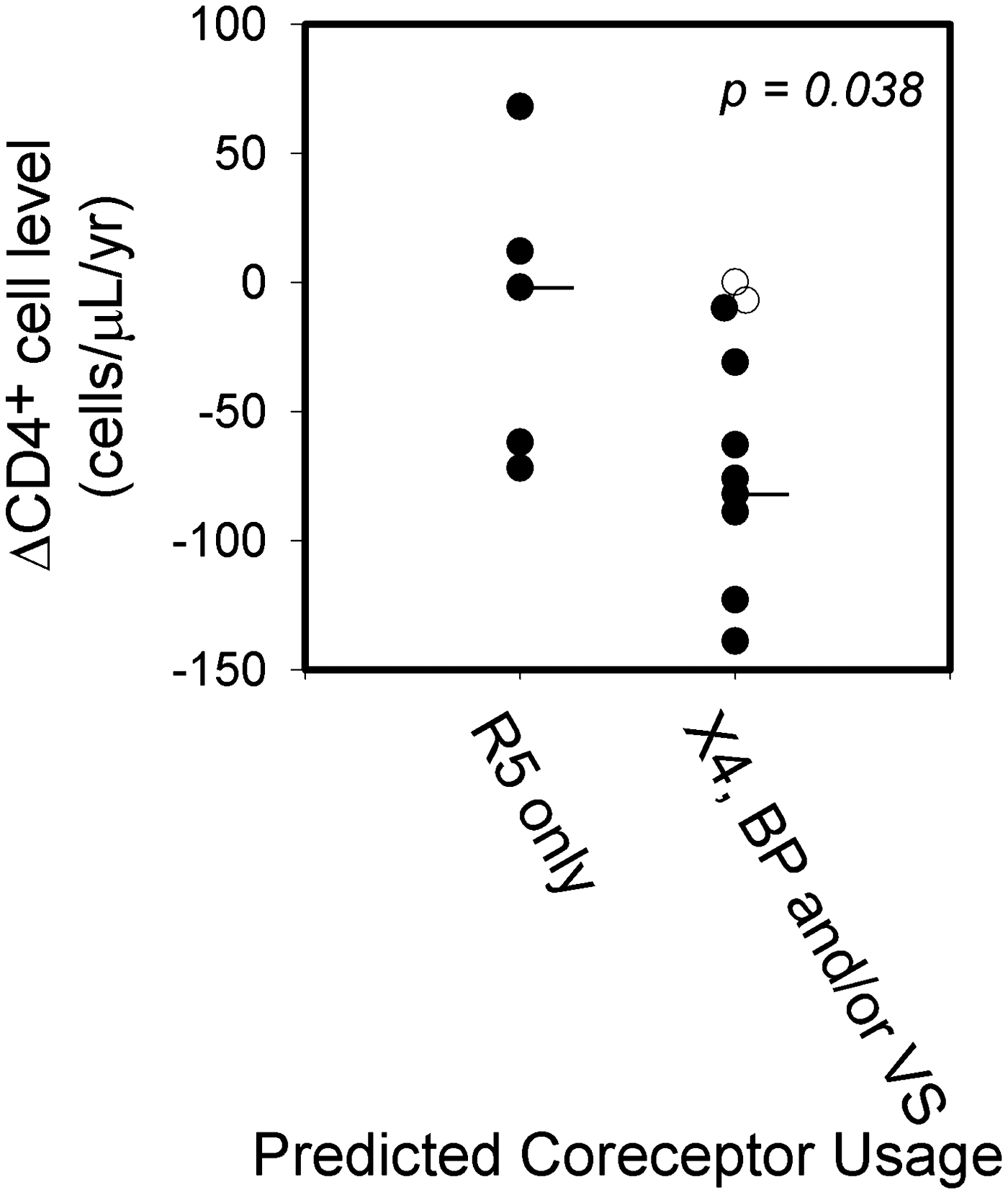
Comparison of changes in CD4+ cell levels for women who shed predicted R5 viruses only, or X4 viruses in VS and/or BP samples for ≥1 sampling times. Horizontal lines indicate median change in CD4+ cell level/year for each group. Two women (patient IDs 35 and 50 with CD4+ cell level changes shown as unfilled circles) were excluded from this analysis because their CD4+ cell levels were very low for all sampling dates (≤40 cells/μl).
A cross-sectional examination of our data using the relative amounts of R5 and X4 viruses in each sample indicated that greater amounts of X4-tropic viruses were associated with lower blood CD4+ cell levels for both VS and BP samples. Although modest, the rank correlations between blood CD4+ cell levels and λ values for VS and BP (r=0.61, p<0.001; r=0.45, p=0.003, respectively) indicate that greater amounts of R5- and X4-tropic viruses are associated with higher and lower blood CD4+ cell levels, respectively, in both anatomical compartments.
Despite the correlation between the relative abundances of R5- and X4-tropic viruses in VS and BP (VS and BP λ values, r=0.69, p<0.001) and the evidence that the abundances at both sites appear to be influenced by a common factor (CD4+ cell level), significant differences between VS and BP λ values (≥ 0.4, p≤0.05) were found for seven (16%) matched sample pairs of four women (Table 1). No significant distinctions in blood CD4+ cell levels, viral loads, or use of antiretroviral therapy were found between these seven matched samples and the remaining VS and BP pairs.
N-Glycosylation densities of HIV-1 shed in VS and BP
Previous data suggest that N-linked carbohydrate moieties on the HIV-1 envelope may be important in virus infectivity and viral escape of host immune responses. 37 To examine N-glycosylation densities, we identified affected N-X-S/T sites in predicted amino acid sequences of VS and BP. 38 The number of potential N-glycosylation sites ranged from 5 to 10 sites (median=8) for VS sequences, and 5 to 9 sites for BP sequences (median=8). Over time, changes in the mean number of N-glycosylation sites (calculated from the 10 sequences from each sample) between consecutive sampling times ranged from 0 to 1.5 for VS samples (median=0.3) and from 0 to 1.0 for BP (median=0.3). These changes were correlated between matched VS and BP samples (r=0.72, p<0.0001, data not shown), but they were not significantly associated with fluctuations in blood CD4+ cell levels or viral loads. Overall, the mean numbers of N-glycosylation sites between matched VS and BP samples were strongly correlated (r=0.86, p<0.0001). Even so, in comparisons of these mean N-glycosylation densities between paired samples, significant differences were found for six sampling times (13%) of five women (Table 1). No significant differences in blood CD4+ cell levels, viral loads, or the use of antiretroviral therapy were found between these six clinic visits and the remaining sampling times.
In cross-sectional examinations of N-glycosylation densities and CD4+ cell levels, a greater range of the number of VS and BP N-glycosylation sites was found for the low (<200 cells/μl) and high (>400 cells/μl) CD4+ cell level categories compared to the range observed for CD4+ cell counts of 200–400 cells/μl. Also, N-glycosylation densities among the three CD4+ cell level categories showed significant differences in both VS and BP, with the 200–400 cells/μl category having a higher number of N-glycosylation sites than the low or high CD4+ cell level categories (Fig. 3A).
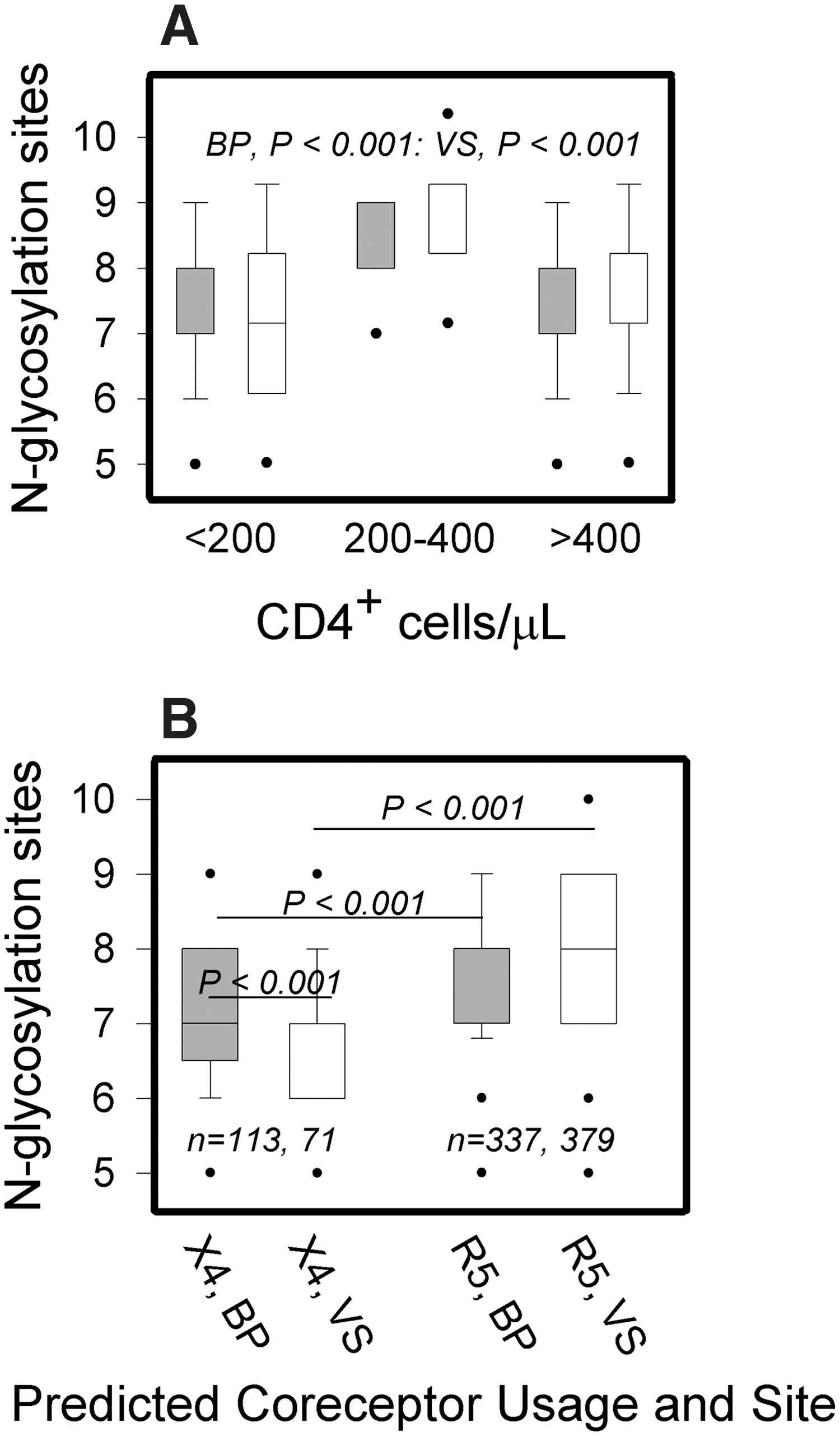
Cross-sectional analyses of the potential N-glycosylation densities of HIV-1 C2V3C3 sequences.
A previous study of N-glycosylation sites in R5- and X4-tropic viruses reported strain-specific differences. 39 In our study, a cross-sectional examination of all sequences showed that R5 viruses had higher N-glycosylation densities than X4 viruses (medians=8.0 and 7.0, respectively, p<0.0001). Further analysis revealed VS X4 sequences had significantly fewer sites than BP X4 viruses to yield the following order of N-glycosylation densities: BP R5 ∼ VS R5 > BP X4 > VS X4 viruses (Fig. 3B). However, when these analyses were performed individually for each of the five patients who had predicted X4 viruses in both her VS and BP samples, this trend in N-glycosylation densities was significant only for patient 035.
Discussion
Many studies of HIV-1 transmission report detection of R5-tropic viruses only in individuals newly infected through heterosexual contact. 21,23,24 Our study sought to determine if this predominance of R5-tropic sexual transmission could be due in part to a relative scarcity of X4-tropic virus shedding in genital secretions compared to that in the blood of chronically infected women. We also wanted to know if differences in predicted glycosylation densities between VS and BP reported in cross-sectional studies 14 were maintained over time. Finally, we sought to know if predicted viral tropism and glycosylation patterns in HIV from VS and BP were affected differently by changes in the immune status of chronically infected women.
Our analysis of the predicted amino acid sequences of genomic viral RNA indicates that R5-tropic viruses were the most common variant in VS samples but that X4 strains were also present at 10 (22%) study visits and comprised all the VS quasispecies sampled at five (11%) of those visits. Although X4 viruses were present in blood without being detected in matched VS samples, they were detected in VS only when X4 viruses were present in BP. This suggests that shedding of X4 viruses in female VS is preceded by that in peripheral blood. In fact, our detection of X4 shedding in VS was more likely to occur when X4 viruses were 50% or more of the blood quasispecies in a matched BP sample. In addition, our results and a previous study 14 showed that when there was a significant difference in the ratio of R5/X4 viruses between matched VS and BP samples, the proportion of X4 viruses was always higher in BP. However, it is likely that neither the X4 nor R5 viruses circulating in peripheral blood become a significant proportion of the cell-free HIV-1 that is shed in a woman's genital secretions. We have previously reported that local virus replication in the female genital tract, not transudation of cell-free virus from blood plasma, is the major source of HIV in women's genital secretions. 19 In addition, we determined that less than 5% of a vaginal virus load during intermenses is due to whole blood mixing with vaginal secretions. 40 –42
As HIV-1 disease evolves, coreceptor switching from R5 to X4 tropism in blood is common among subtype B HIV-1-infected individuals. 12 The R5-to-X4 coreceptor switch is often a precursor of accelerated clinical disease progression accompanied by a large decline in peripheral blood CD4+ cell counts. 12,29,30,43 In our study, there was an overall correlation between VS and BP R5/X4 virus ratios and CD4+ cell counts. In addition, our longitudinal analysis showed that women who had greater declines in CD4+ cell counts over time were more likely to shed X4 viruses in VS as well as BP. These results suggest that the immune dysregulation during HIV-1 disease progression that allows for increased shedding of X4-tropic viruses in the female genital tract is similar to that in blood.
The mechanism for coreceptor switching to X4 virus shedding in women's genital secretions is not known. Reports that indicate local virus replication as the major source for most of the HIV shed in the female genital secretions 19,40,42 predict that infected cells replicating X4 virus in the mucosa are necessary for X4 shedding in the female genital tract. Thus, these X4-infected cells could arrive from peripheral blood as part of the transient population of CD4+ lymphocytes with mucosal homing receptors that infiltrate the genital mucosa on a continual basis. 44,45 This concept is further supported by observations that virus-infected cells trafficking from peripheral blood is the major proviral pool of infected cells in the genital mucosa and that mucosal proviral evolution is not compartmentalized from that in blood over time. 15 In contrast, previous cross-sectional studies report molecular evidence of limited HIV RNA compartmentalization in women's genital tissues 14 that could possibly support an independent evolution of X4 viruses. However, the evidence that X4-tropism evolves in the thymus and not other tissues such as lung, lymph nodes, and spleen 46 suggests that the female genital tract would also likely not support an independent evolution of X4-tropism.
In addition to similar coreceptor tropism profiles between VS and BP, we also found a strong correlation for N-glycosylation densities between matched VS and BP samples. Moreover, changes in N-glycosylation densities over time were correlated between matched VS and BP samples. These results suggest that N-linked carbohydrate density in PB and VS is usually affected by one or more common factors. Our data indicated that immunological integrity could be a factor; both VS and BP viruses had lower N-glycosylation densities at lower blood CD4+ cell levels (<200 cells/μl) compared to more moderate levels (400–200 cells/μl) as reported in a previous cross-sectional study. 14 Since N-linked carbohydrates on the viral envelope are implicated in the resistance to antibody-mediated neutralization, 14,47,48 the decline of host immunity (CD4+ cell counts <200 μl) may have decreased the selective pressure for maintaining these glycosylation sites as discussed previously. 39,47 Yet we were surprised that viral glycosylation density was also lower at high CD4+ cell levels (>400 cells/μl) compared to those at moderate levels (400–200 cells/μl). However, data from Bunnik et al. 49 showed that predicted env glycosylation densities in proviral DNA from peripheral blood mononuclear cells of infected persons continued to increase for as long as 3.5 to 8 years after seroconversion in chronically infected persons and peaked after their CD4+ levels were <400 cell/μl. Thus, our data expand on this previous study to show that glycosylation densities of HIV shed in both VS and BP of chronically infected women continue to increase during the early and middle phases of HIV-induced immune suppression.
Despite the evidence that N-glycosylation densities of HIV-1 in both VS and BP are associated with immune status, significant differences in the carbohydrate densities of matched VS and BP samples did occur in a small percentage of clinic visits for 33% of the women in our study. At some of those visits, VS and BP glycosylation differences could be explained by corresponding differences in their predicted coreceptor tropism. At the remaining visits where glycosylation densities were different but coreceptor tropism was similar, there was no trend for glycosylation being higher in VS or BP. In addition, differences in VS and BP glycosylation densities were transient; none of the women sustained a significant glycosylation difference for all three clinic visits and only one woman did for two visits. Thus, the lack of persistent glycosylation differences we observed between HIV shed in VS and BP suggests that mucosal immune pressures against virus in the women's genital mucosa were not driving HIV glycosylation densities independent of that in peripheral blood over time. However, our finding of lower glycosylation densities for X4 viruses in VS compared to BP may indicate that X4 virus shedding in the female genital tract undergoes selection for fewer N-glycosylation sites. However, the fact that most X4 virus in VS was detected later in disease progression when overall glycosylation densities were lower suggests that the level of overall immune dysfunction and not a compartmentalized mucosal selection mechanism was the dominant factor of this difference.
In summary, our longitudinal study of predicted HIV-1 coreceptor usage and envelope glycosylation density supports the concept of a dynamic process whereby subpopulations of infected cells in peripheral blood continually migrate to mucosal tissues, clonally expand, and shed virus in a woman's genital secretions. 50 Although our longitudinal study was not designed to use currently accepted genotype compartmentalization analyzes for VS and BP viruses, 51,52 our results support other longitudinal studies that showed a lack of sustained compartmentalized evolution of HIV-1 in the genital mucosa of women. 53 In fact, our results and those in a previous study that reported a pattern of similar antiretroviral therapy-associated resistance mutations between viruses in VS and PB over time 54 provide further evidence that mucosal tissues are continually being repopulated with HIV-infected cells migrating from peripheral circulation. As the samples selected for this study contained high virus loads in both VS and BP, future longitudinal studies will also need to examine the coreceptor tropism of virus in women with low virus loads. The presence of significant levels of X4 tropic virus in VS during mostly late-stage disease suggests that these genital tract viruses are available for sexual transmission. However, it is unknown whether these genital tract viruses are less transmissible or that decreased sexual activity due to high-level morbidity during late stage disease is responsible for their lack of transmission. Moreover, as R5-tropic entry inhibitors such as Maraviroc come into wider use therapeutically and possibly as a microbicide agent, it will be important to determine if they significantly increase X4 virus genital shedding in persons failing therapy and if those X4 viruses are more infectious and more pathogenic following sexual transmission.
Footnotes
Acknowledgments
The findings and conclusions in this report are those of the authors and do not necessarily represent the views of the Centers for Disease Control and Prevention or the Department of Health and Human Services.
Author Disclosure Statement
No competing financial interests exist.