
Abstract
The HIV-1 restriction factor SAM domain and HD domain-containing protein 1 (SAMHD1) blocks HIV-1 infection in human myeloid cells. Mutations in the SAMHD1 gene are associated with rare genetic diseases including Aicardi–Goutieres syndrome. However, it is unknown whether polymorphisms of SAMHD1 are associated with infection and natural control of HIV-1 in humans. Our objective was to determine whether the expression of SAMHD1 mRNA is affected by common single nucleotide polymorphisms (SNPs) in SAMHD1 and whether the SNPs are associated with HIV-1 infection status. Using a tagging SNP approach, we determined the association between eight tagging SNPs in SAMHD1 and the mRNA expression in B-lymphocyte cell lines from 70 healthy white donors. We identified one SNP (rs1291142) that was significantly associated with SAMHD1 mRNA expression, with minor allele carriers having 30% less mRNA levels (p=0.015). However, after analyzing the published genome-wide association study data of 857 HIV-1 controllers and 2088 HIV-1 progressors from the European and African-American cohorts, we did not find a significant association between SNPs in SAMHD1 and HIV-1 infection status, including SNP rs1291142 (p>0.05). We also observed 2- to 6-fold variations of SAMHD1 mRNA levels in primary B-lymphocytes, CD4+ T-lymphocytes, and CD14+ monocytes from five healthy donors. Our results suggest that common regulatory polymorphism(s) exist in the SAMHD1 gene that affects its mRNA expression in B-lymphocyte cell lines from healthy whites. However, polymorphisms of SAMHD1 are unlikely to contribute to the infection and natural control of HIV-1 in European and African-American individuals.
Introduction
P
HIV-1 restriction factors are host proteins that potently block viral replication in the cell. 4 –6 The well-characterized HIV-1 restriction factors include APOBEC3G (apolipoprotein B mRNA editing enzyme, catalytic polypeptide-like 3G), 7 TRIM5α (tripartite motif 5α), 8 and Tetherin (BST-2 or CD317). 9,10 Polymorphisms in the genes encoding HIV-1 restriction factors have been shown to associate with HIV-1 infection and AIDS progression in humans. For example, polymorphisms in the APOBEC3G and TRIM5α genes are associated with status and clinical course of HIV-1 infection, 11 –14 suggesting that these HIV-1 restriction factors contribute to HIV-1 infection and AIDS progression in vivo.
SAMHD1 is a cellular protein that restricts HIV-1 infection in human myeloid-lineage cells, such as monocytes, dendritic cells, and macrophages. 15,16 As a dGTP-regulated deoxynucleoside triphosphate (dNTP) triphosphohydrolase, SAMHD1 degrades dNTPs 17,18 and mediates HIV-1 restriction by decreasing the concentration of the cellular dNTP pool. 19 Mutations in SAMHD1 are associated with a rare genetic disorder known as Aicardi–Goutieres syndrome (AGS). 20 Patients with AGS have mutations in multiple exons of SAMHD1 that affect protein expression or in an intron that alters the splicing acceptor site of SAMHD1. 20 SAMHD1-deficient monocytes from one AGS patient have been shown to be highly susceptible to HIV-1 infection in vitro, 21 suggesting that SAMHD1 may block HIV-1 infection in myeloid cells in vivo. Moreover, a mutation in the splice acceptor site of an intron in SAMHD1 results in a shorter mRNA and an aberrant protein with an in-frame deletion, which is associated with cerebral vasculopathy and early onset stroke in Amish individuals. 22 However, it is unknown whether common regulatory polymorphisms exist in SAMHD1 and whether such polymorphisms are associated with infection and natural control of HIV-1 in humans.
In this study, we hypothesized that common SAMHD1 SNPs may affect its mRNA expression and such SNPs could be associated with HIV-1 infection status in humans. We identified an intron SNP in SAMHD1 that significantly affects the mRNA expression of SAMHD1 in B-lymphocytes from 70 healthy white donors. However, our analysis indicated that SNPs in SAMHD1 are not associated with HIV-1 infection status among 857 HIV-1 controllers and 2088 HIV-1 progressors from the European and African-American cohorts. 3 Our results suggest that polymorphisms of SAMHD1 are unlikely to contribute to the infection and natural control of HIV-1 in European and African-American populations.
Materials and Methods
Selection of intron SNPs in the SAMHD1 gene
Tagging SNPs in SAMHD1 were selected from HapMap
23
setting parameters for an minor allele frequency over 10% and linkage disequilibrium of 0.8 in the white population (Table 1). Parameters yielded eight tagging SNPs: rs4810362, rs6030346, rs6065494, rs6072746, rs6016783, rs7268811, rs12481143, and rs1291142. The sequence, reference genotype and variant, and allele frequency for each of these SNPs were determined using the SNP database on
SNP, single nucleotide polymorphism; MAF, minor allele frequency. Used tagging SNPs, subset of SNPs designed to predict other SNPs in the region, to capture other SNPs. This table is based on the search of the HapMap database (the catalog of human genetic variants 23 ).
Isolation of primary B-lymphocytes, monocytes and CD4+ T-lymphocytes
Primary B-lymphocytes, monocytes, and resting CD4+ T-lymphocytes were isolated from fresh peripheral blood mononuclear cells (PBMCs) of five healthy blood donors using anti-CD19-, anti-CD14-, and anti-CD4-coated magnetic particles (BD Biosciences), respectively. The purity of the isolated primary cells (>98%) was confirmed by specific antibody staining and flow cytometry.
Genomic DNA and cDNA samples
Seventy Epstein–Barr virus (EBV)-transformed B-lymphocyte cell lines from healthy white donors were obtained from Coriell Cell Repositories as previously described. 24 Genomic DNA and total RNA were isolated from the B-lymphocytes and the RNA samples were treated with DNase I to avoid genomic DNA contamination as described. 24 Total cellular RNA samples from B cell lines, primary B-lymphocytes, monocytes, and CD4+ T-lymphocytes were used to synthesize cDNA using a reverse transcription kit as previously described. 24 All samples were tested by real-time PCR, showing no amplification after 40 cycles in cDNA preparation without reverse transcriptase.
Genotyping
We used allele-specific real-time PCR for SNP genotyping as previously described. 25 Three primers were designed for each SNP. A reverse primer was designed and then two separate forward primers were designed to recognize the reference genotype and allele variants of each SNP (Table 2). Real-time PCR was conducted with the CFX 96-Bio-Rad Real-time PCR Detection System using SYBR Green (Bio-Rad) and the following protocol: step 1 (initiation): 95°C for 3 min; step 2 (denature): 94°C for 30 s; step 3 (annealing): 60°C for 45 s; step 4 (extension): 72°C for 1 min. Steps 2–4 were repeated for 40 cycles. Genomic DNA was used as a template in real-time PCR. There were at least three cycle differences between primer pairs targeting reference or variant alleles.
This table contains two primers for quantification of cDNA and three primers for each of the SNPs. Primers for genotyping contain a reference genotype primer, an SNP allele primer, and either a reverse/forward primer that concurs with the others.
SAMHD1 mRNA quantification
The amount of SAMHD1 mRNA was quantified by real-time PCR using the primers shown in Table 2. β-Actin was used as a control (Forward primer: 5′-CCTGGCACCCAGCACAAT-3′; Reverse primer: 5′-GCCGATCCACACGGAGTACT-3′) for normalization. The condition of real-time PCR reaction was as described above and cDNA was used as a template.
Statistical analysis
The amount of SAMHD1 mRNA was calculated as the ratio of SAMDH1/β-actin amplified by 100,000. Since the data were not normally distributed, we transformed the data into a log10 scale. The HelixTree program (Golden Helix, Bosman, MT) was used for initial screening of multiple SNPs. If any positive SNPs were identified, statistical analysis was performed using Minitab software by fitting single linear regression models as described. 24,25 The goodness of fit was judged by residual plots and normal quartile plots, testing the fulfillment of linear regression assumptions, which included constant variance, normality, and independency.
Analysis of the datasets of HIV-1 controllers and progressors
Sample sizes of the analyzed data were 857 HIV-1 controllers and 2088 HIV progressors (total n=2945, Table 3). The two available datasets contain data from European HIV-1 controllers and progressors (comparing 516 HIV-1 controllers to 1196 HIV-1 progressors) and African-American HIV-1 controllers and progressors (comparing 341 HIV-1 controllers to 892 HIV-1 progressors). The analysis was performed using the online Ricopili tool on
This table is based on the International HIV Controllers Study
3
and the datasets are available at
Results
Polymorphism of SAMHD1 affects mRNA expression levels in B-lymphocytes from healthy white donors
Human SAMHD1 spans 59,532 bp of genomic sequence in 16 exons in chromosome 20 (position 34,954,059–35,013,590) and encodes a 626-amino acid protein. 20 Human peripheral blood leukocytes express high levels of SAMHD1 mRNA. 27 EBV-mediated transformation of B-lymphocytes is a standard protocol to generate immortalized cells to preserve genetic materials of individuals. 3,24 We used EBV-transformed B-lymphocytes from 70 healthy white donors as the source of SAMHD1 mRNA and measured the mRNA levels of SAMHD1 by real-time PCR methods. The mRNA levels of β-actin were also measured and used as a control. Our results indicated that SAMHD1 mRNA was expressed in B-lymphocytes at the level of ∼0.01% of β-actin mRNA. In 70 B-lymphocyte cell lines derived from different donors, the average level of SAMHD1 mRNA was 69±55 (mean±SD, arbitrary unit), with 50-fold interindividual variability (ranging from 7 to 355). These results suggest the expression levels of SAMHD1 mRNA in B-lymphocytes vary considerably among different individuals.
To identify frequent polymorphisms in the SAMHD1 gene that may affect its mRNA expression, we selected eight tagging SNPs in SAMHD1 introns with minor allele frequencies over 10% (Table 1) and associated the genotypes with SAMHD1 mRNA expression. Interestingly, single SNP analysis revealed that one SNP (rs1291142) was significantly associated with SAMHD1 mRNA expression (Fig. 1). SNP rs1291142 is located in intron 15 of SAMHD1 and presents as GG for the reference genotype and GA or AA for variants. B-lymphocytes with a GA or AA genotype showed 0.67-fold (p=0.015) lower SAMHD1 mRNA levels than the B-lymphocytes homozygous for reference (GG) genotype (Fig. 1). By contrast, there was no significant association between the other seven SNPs and SAMHD1 mRNA levels. These results suggest that a regulatory polymorphism exists in the SAMHD1 gene, which can significantly affect SAMHD1 mRNA expression in B-lymphocytes from healthy whites.
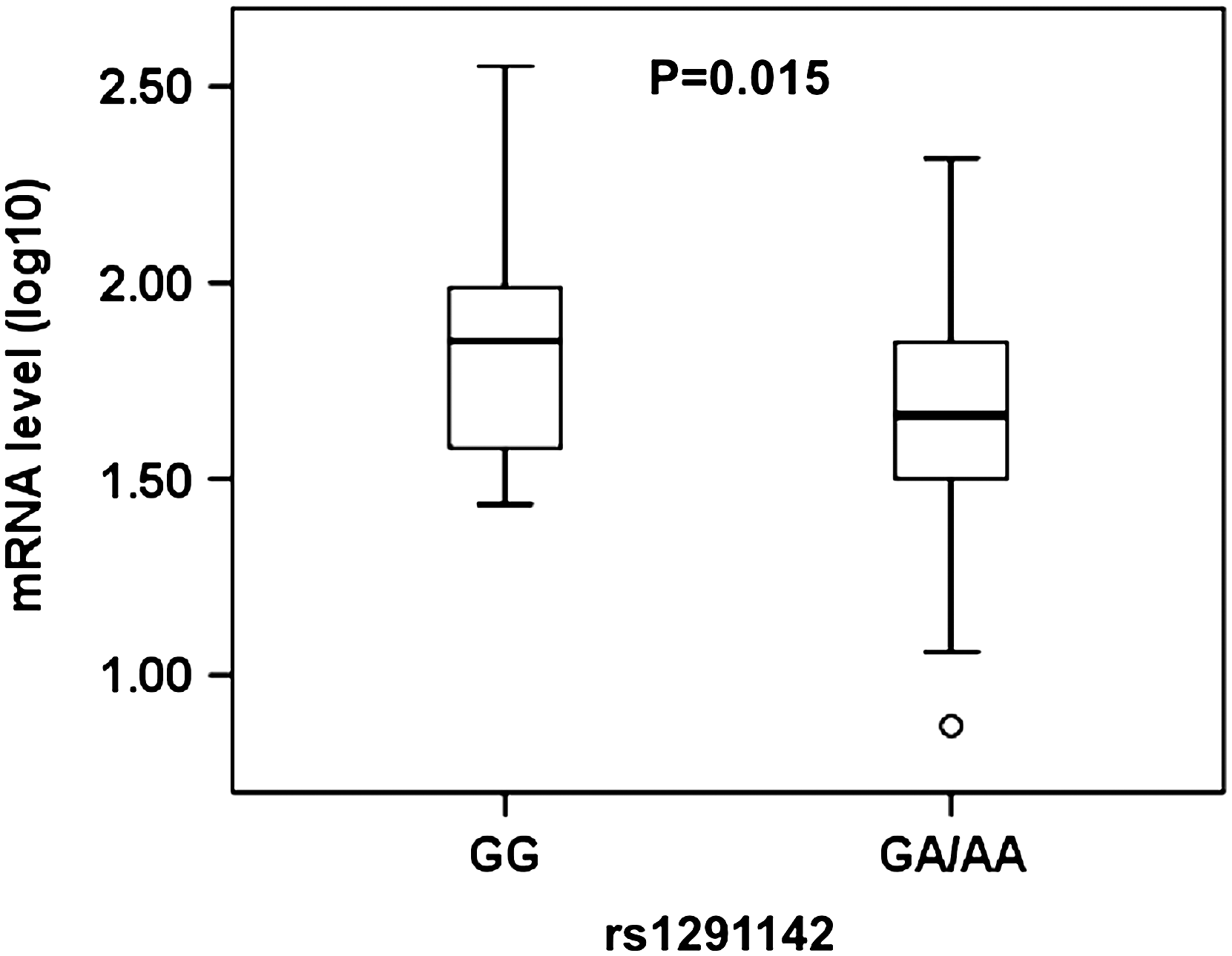
Polymorphism rs1291142 of SAMHD1 affects its mRNA expression levels in B-lymphocytes from healthy white donors (n=70). The mRNA levels of SAMHD1 in B-lymphocytes from 70 healthy white donors were measured using real-time PCR methods. The GA and AA carriers of SAMHD1 single nucleotide polymorphisms (SNPs) (rs1291142) show 0.67-fold lower SAMHD1 mRNA than those of GG carriers (p=0.015, two-sided, linear regression).
Comparison of SAMHD1 mRNA levels in primary B-lymphocytes, monocytes, and CD4+ T-lymphocytes
To compare SAMHD1 mRNA levels in different types of immune cells, we measured SAMHD1 mRNA levels in primary B-lymphocytes, monocytes, and CD4+ T-lymphocytes isolated from PBMCs of five healthy blood donors using real-time RT-PCR. We compared the mRNA levels of SAMHD1 relative to β-actin controls in these samples. Our results indicated that the average SAMHD1 mRNA levels in primary B-lymphocytes are lower (p>0.1) than that in CD4+ T-lymphocytes and CD14+ monocytes from five donors. The average SAMHD1 mRNA level in primary CD14+ monocytes was the highest among these cell types (Table 4), although these differences are not statistically significant (p≥0.19). In addition, we observed 2- to 6-fold variations of SAMHD1 mRNA levels among different donors and cell types (Table 4). The analysis does not allow us to conclude that there is a difference in SAMHD1 mRNA levels among different cell types, although there may be a pairwise trend of lower SAMHD1 mRNA expression in B-lymphocytes.
Real-time RT-PCR was used to measure the mRNA levels of SAMHD1 and β-actin. The relative mRNA levels of SAMHD1 compared to β-actin controls are shown (arbitrary unit). Each sample was detected in triplicate.
Polymorphisms of SAMHD1 are not significantly associated with HIV-1 infection status of controllers and progressors from the European and African-American cohorts
A previous genome-wide association study using the samples of HIV-1 controllers and progressors from the European and African-American cohorts has yielded two public data sets containing genome-wide SNPs of HIV-1 controllers
3
and
A total of 23 and 29 SNPs in SAMHD1, including seven of the eight SNPs that we tested in this study, were genotyped in the HIV-1 controllers and progressors from the European and African-American cohorts, respectively (Table 3 and Supplementary Tables S1 and S2; Supplementary Data are available online at
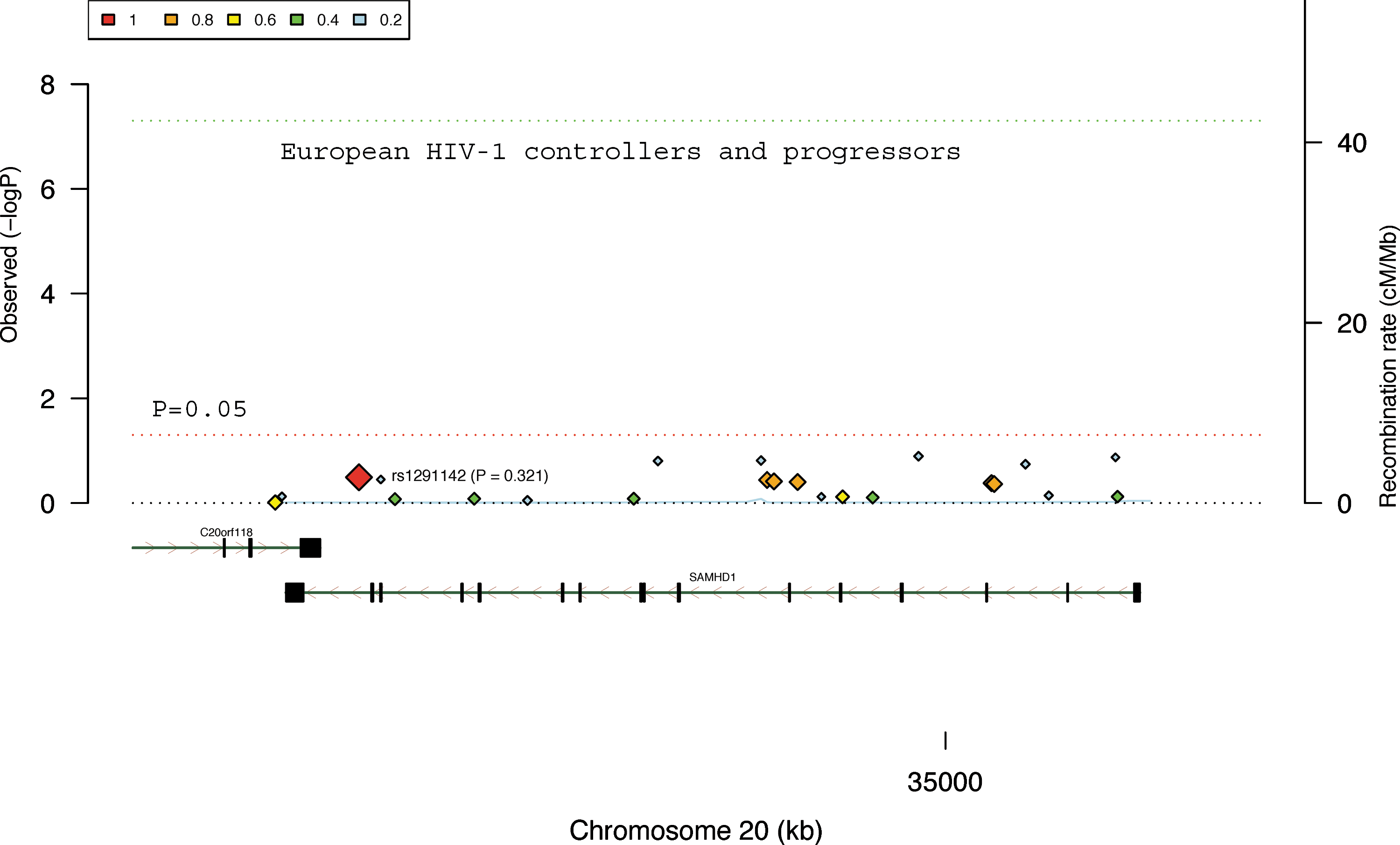
Polymorphisms of SAMHD1 are not significantly associated with HIV-1 infection status of HIV-1 controllers and progressors from the European cohort. The results were derived from the analysis of 23 available SNPs in SAMHD1 in 516 HIV-1 controllers and 1196 HIV-1 progressors. The color squares and the sizes of color diamonds represent the r 2 values of the linkage disequilibrium. The SAMHD1 gene, SNP (rs1291142), and its p value (0.256) are labeled. The red and green dot line represents p=0.05 and genome-wide significance (p<5×10−8), respectively. The x-axis indicates the location and organization of the genes. The blue line represents the gene recombination rate (y-axis on the right side).
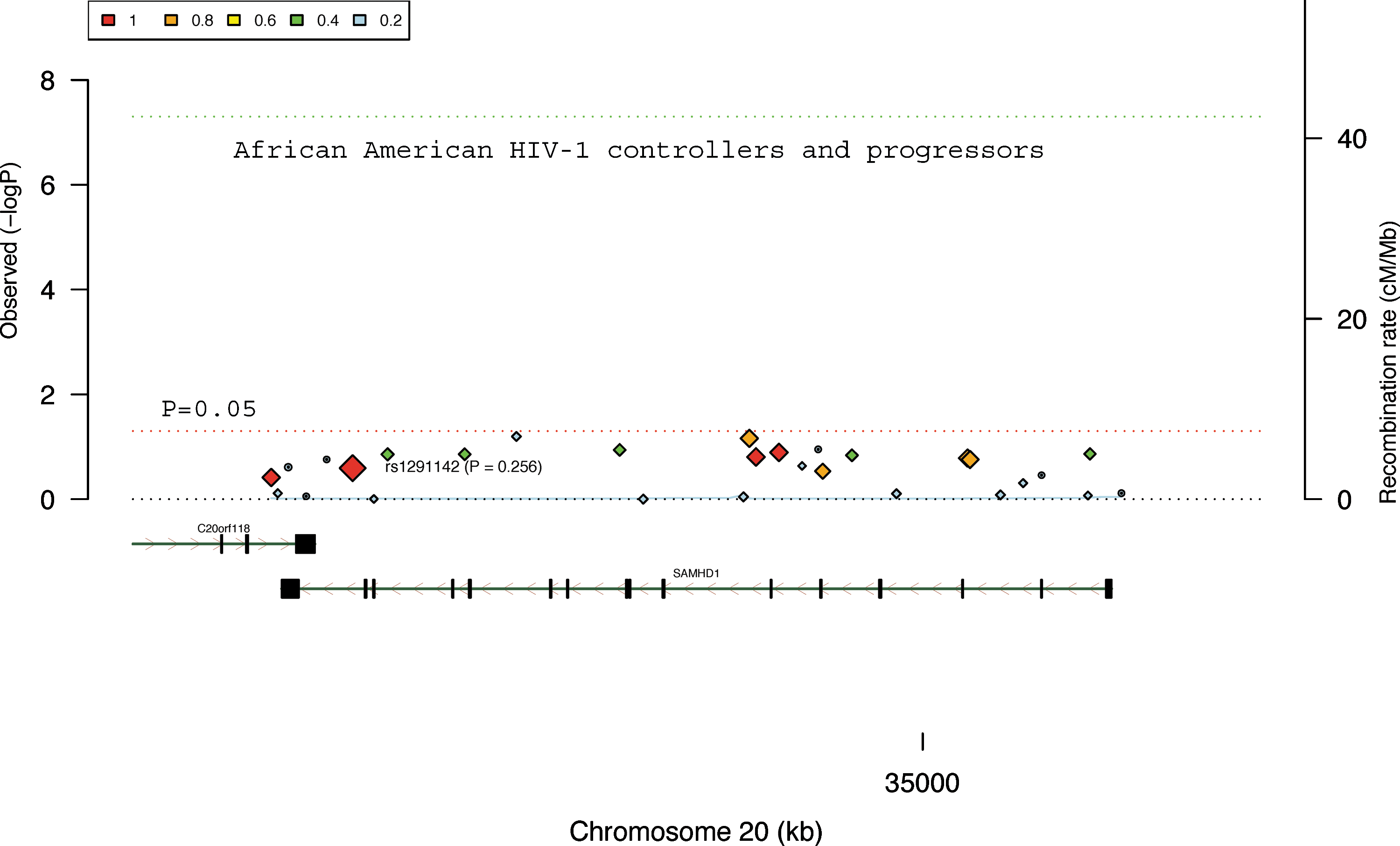
Polymorphisms of SAMHD1 are not significantly associated with HIV-1 infection status of HIV-1 controllers and progressors from the African-American cohort. The results were derived from the analysis of 29 available SNPs in SAMHD1 in 341 HIV-1 controllers and 892 HIV-1 progressors. The color squares and the sizes of color diamonds represent the r 2 values of the linkage disequilibrium. The SAMHD1 gene, SNP (rs1291142), and its p value (0.321) are labeled. The red and green dot line represents p=0.05 and genome-wide significance (p<5×10−8), respectively. The x-axis indicates the location and organization of the genes. The blue line represents the gene recombination rate (y-axis on the right side).
The results were derived from the analysis of the dataset at
The results were derived from the analysis of the dataset at
Polymorphisms of TRIM22 are significantly associated with HIV-1 natural control in controllers from the African-American cohort
To generate a positive control for association with HIV-1 infection and progression, we analyzed polymorphisms of the TRIM22 gene in the same datasets used for SAMHD1 SNP analysis as a positive control. A recent study has shown a significant correlation of TRIM22 mRNA levels associated with HIV-1 viral loads, suggesting that TRIM22 likely acts as an antiviral effector in vivo. 28 Our analysis revealed that an SNP rs11038741 in the TRIM22 gene is significantly associated with natural control of HIV-1 infection in HIV-1 controllers compared to progressors (linkage disequilibrium=1, p=0.00459) in the African-American cohort (Fig. 4), but not in the European cohort (data not shown). These results validated our correlation analysis of SAMHD1 SNPs and HIV-1 infection using the same dataset and methods.
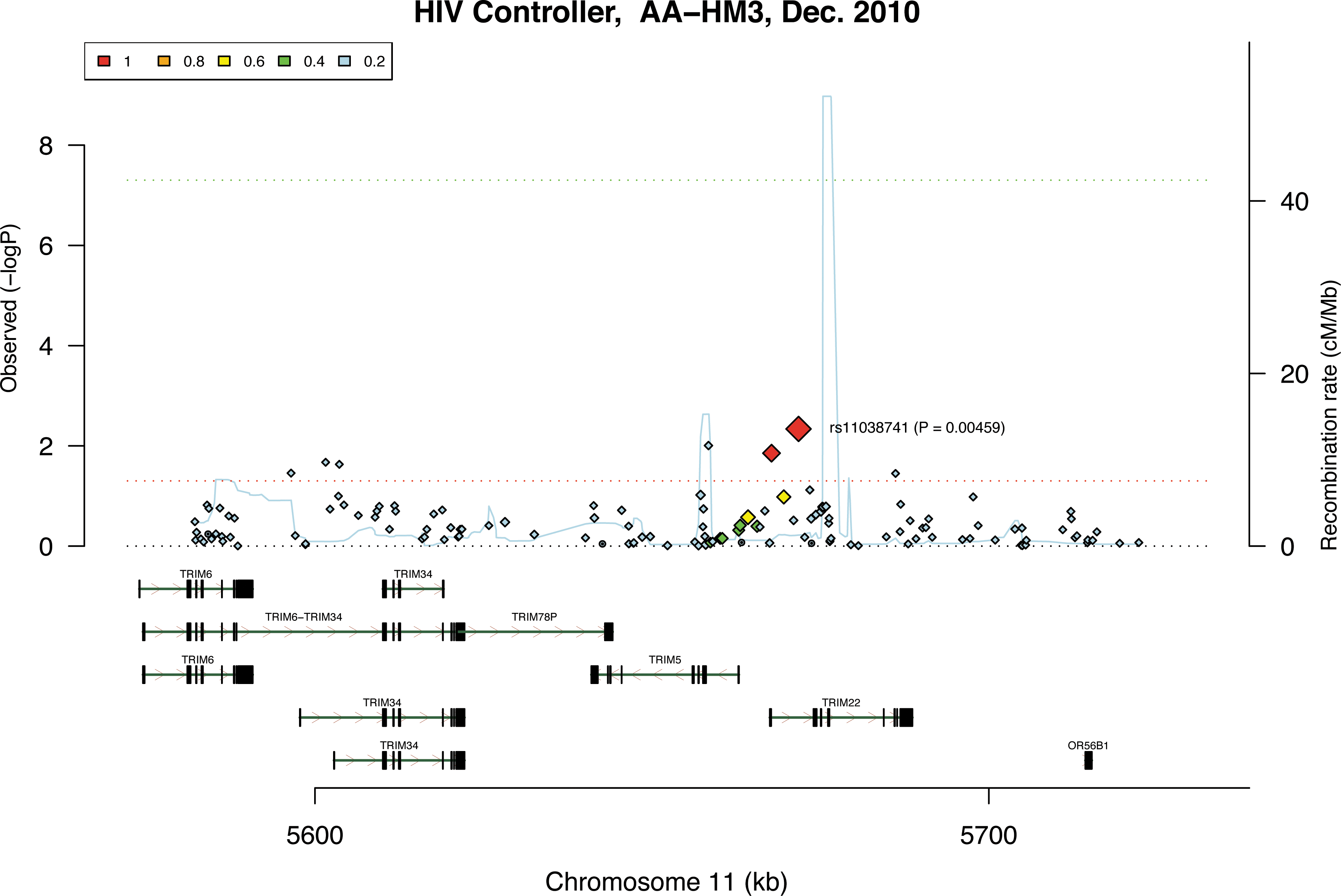
Polymorphisms of the TRIM genes are significantly associated with natural control of HIV-1 infection in HIV-1 controllers from the African-American cohort. The results were derived from the analysis of 133 available SNPs of the TRIM family genes in 341 HIV-1 controllers compared to 892 HIV-1 progressors. The color squares and the sizes of color diamonds represent the r 2 values of the linkage disequilibrium. The TRIM genes and SNP rs11038741 (p=0.00459) are labeled. The red and green dot line represents p=0.05 and genome-wide significance (p<5×10−8), respectively. The x-axis indicates the location and organization of the genes. The blue line represents the gene recombination rate (y-axis on the right side).
Discussion
Given the important role of SAMHD1 in HIV-1 infection in vitro and that several rare exon and intron mutations of SAMHD1 are associated with AGS and other genetic disorders, 20,22 it is conceivable that common regulatory polymorphisms of SAMHD1 that affect its mRNA expression may play an important role in controlling HIV-1 infection in vivo. Indeed, our results indicated that SNP rs1291142 significantly affected SAMHD1 mRNA expression in B-lymphocytes from 70 healthy white donors, suggesting that a common polymorphism exists in SAMHD1 and can regulate its mRNA expression in B-lymphocytes. However, SNP rs1291142 and other SNPs in SAMHD1 are not associated with HIV-1 infection status. The effect of this SNP on SAMHD1 mRNA expression is limited with a 0.67-fold difference, which may not be significant enough to exert the effect on SAMHD1 protein expression and HIV-1 restriction in vivo. Other host factors (such as microRNA) may also affect mRNA expression, since there were 50-fold interperson differences among 70 healthy white donors.
SNP rs1291142 itself is unlikely to be functional; it may be in high linkage disequilibrium with functional SNPs, because it is located in the middle of intron 15 of SAMHD1 and is not a splicing acceptor or donor site. However, we cannot completely rule out the possibility that SNP rs1291142 itself is functional, because SNPs in introns can modulate RNA levels of some human genes by affecting RNA elongation. 24,25,29 It is unclear whether this SNP affects SAMHD1 mRNA expression in primary B-lymphocytes or other cell types from healthy donors or from HIV-1 infected individuals, since the regulation of polymorphisms on the expression of other genes has been shown to be tissue specific. 25,29 Given that SAMHD1 is a myeloid-cell-specific HIV-1 restriction factor, 15,16 it is interesting to test whether the same polymorphism affects SAMHD1 mRNA expression in myeloid cells.
Previous studies indicated that SAMHD1 mRNA expression is upregulated by the infections of adenovirus, rabies virus, and respiratory syncytial virus in human cells. 30 –32 It is unknown whether EBV infection of B-lymphocytes may affect the expression of SAMHD1 mRNA in the cell. Our real-time PCR-based SAMHD1 cDNA quantification was based on the relative levels of SAMHD1 and β-actin mRNAs, which represented the relative levels of SAMHD1 mRNA among all 70 individuals' samples. Furthermore, our real-time PCR data suggest that the average level of SAMHD1 mRNA expression in EBV-transformed B-lymphocytes was lower than that in primary B-lymphocytes. Thus, our correlation analysis between the SNP genotypes and the mRNA expression levels of SAMHD1 was valid. However, it is possible that the regulation of SAMHD1 mRNA expression in EBV-transformed B-lymphocytes is different from that in other cell types. For example, HIV-1 target cells including myeloid cells in vivo may be subjected to certain regulations of SAMHD1 expression after HIV-1 infection, which may override the effect from SAMHD1 polymorphisms.
Our analyses based on available datasets suggest that SNPs in SAMHD1 are not significantly associated with HIV-1 infection and the natural control of HIV-1 infection in European and African-American individuals. It would be interesting to examine whether SNPs in SAMHD1 are associated with HIV-1 infection and natural controlling of AIDS in different ethnic populations, such as Asians and Hispanic populations. Of note, SNP rs7268811 in intron 6 of SAMHD1 that we analyzed in the healthy white donors was not found in HIV-1 controllers and progressors from the published European and African-American cohorts (Tables 5 and 6 and Supplementary Tables S1 and S2). It is possible that this SNP has a different allele frequency and heterozygosity among different ethnic populations.
Polymorphisms of other genes in addition to the HLA class I genes can be associated with natural control of HIV-1 infection. For instance, our analysis revealed that an SNP (rs11038741) in the TRIM22 gene is significantly associated with natural control of HIV-1 infection in HIV-1 controllers compared to progressors in the African-American cohort (Fig. 4). Of note, a recent in vitro study indicated that TRIM22 significantly inhibits HIV-1 replication in cells, likely by interfering with viral transcription. 33 Additional studies on the association of TRIM22 with the type 1 interferon response and viral control during primary HIV-1 infection indicate concordance between type 1 interferon and TRIM22, but not human TRIM5α expression in PBMCs, suggesting that TRIM22 likely acts as an antiviral effector in vivo. 28 Collectively, these studies and our analyses suggest that TRIM22 is an HIV-1 restriction factor associated with HIV-1 natural controlling in African-Americans.
SAMHD1 is a novel HIV-1 restriction factor since its restriction function appears to be myeloid cell-type-specific and HIV-1 has not evolved a viral antagonist to counteract SAMHD1-mediated restriction. 34 In contrast, Vpx of less pathogenic HIV type 2 (HIV-2) and certain simian immunodeficiency viruses (such as SIVsm lineage) can induce proteolytic degradation of SAMHD1 and relieve SAMHD1-mediated viral restriction in human myeloid cells. 15,16 The lack of Vpx in HIV-1 may benefit HIV-1 as an immune evasion strategy for the escape from immune surveillance initiated by the infected myeloid cells. 35,36 Further studies of the function and mechanism of SAMHD1-mediated HIV-1 infection can help in understanding the pathogenesis of primate lentiviruses and may aid in the development of more effective interventions against HIV-1/AIDS.
Footnotes
Acknowledgments
We thank the members of the Wu laboratory for helpful discussions. We thank the International HapMap Consortium for the HapMap database and the International HIV Controllers Study for the HIV controller datasets. We thank Drs. Stephan Ripke and Paul McLaren at Massachusetts General Hospital and the Broad Institute for their advice on using the Ricopili datasets. This work was supported in part by grants (AI078762 and AI098524) to Li Wu from the NIH and by the program of Public Health Preparedness for Infectious Diseases (PHPID) of The Ohio State University. Danxin Wang was supported by an NIH grant (GM092655).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.