
Abstract
Women are now becoming the pivot of the epidemiological spread of HIV infection worldwide, especially in developing countries. Therefore, research to develop an efficient microbicide is now a priority for the prevention of HIV-1 acquisition in exposed women. However, recent disappointing failures in microbicide clinical trials revealed major gaps in basic and applied knowledge that hinder the development of effective microbicide formulations. Indeed, the inhibitory power of microbicide molecules may be affected by several physiological and immunological factors present in male and female genital tracts. Furthermore, mucosal crossing of HIV-1 to increase the ability to reach the submucosal target cells (macrophages, lymphocytes, and dendritic cells) may be modulated by supraepithelial factors such as seminal complement components (opsonized HIV-1), by epithelial factors released in the submucosal microenvironment such as antimicrobial soluble factors, cytokines, and chemokines, and by potent intraepithelial and submucosal innate immunity. The design of vaginal microbicide formulations should take into account an understanding of the intimate mechanisms involved in the crossing of HIV through the female genital mucosae, in the context of a mixture of both male and female genital fluids.
Introduction
A
The incidence of HIV is declining globally albeit slowly. 1 This decline in incidence is related to the reduction in risky behavior that has occurred in key sub-Saharan countries. 2 In these countries abstinence, monogamy, and a reduction in the number of sexual partners, and the use of barrier methods such as male and female condoms, are widely advocated. Condom use currently offers the best method to prevent HIV-1 transmission, but it requires partner consent. It seems that abstinence is not a realistic choice for many women, and consistent condom use remains low, especially in primary partnerships, as well as in couples expecting to have children. Likewise, research to develop biomedical approaches can then constitute an alternative strategy to prevent acquisition and transmission of HIV-1. Recently, postexposure and preexposure prophylaxis (PrEP) with antiretroviral drugs (ARVs), 3 treatment of sexually transmitted infections, 4 and male circumcision 5 have been shown to significantly decrease the likelihood of HIV transmission from HIV-positive individuals to their HIV-negative partners. However, before the implementation of circumcision programs, behavioral and social research is needed to find out how people perceive the practice and what messages are needed to persuade them, and to assess the risk of an increase in unsafe sex after circumcision.
Among newly designed preventive methods, ARVs have been highly effective in preventing perinatal transmission. Results of the iPrEx trial demonstrated that the daily oral administration of emtricitabine/tenofovir reduced the risk of HIV acquisition among men who have sex with men by 44%. 6,7 The Partners PrEP Study in HIV-serodiscordant couples from Kenya and Uganda showed that the daily oral administration of tenofovir or an emtricitabine/tenofovir combination reduced HIV-1 infection by 67% and 75%, respectively. 7 –10 Among biomedical approaches, microbicides hold the greatest hope of curtailing AIDS worldwide. Recently, the CAPRISA 004 clinical trial showed, for the first time, that a 1% vaginal gel formulation of tenofovir, a nucleoside analog reverse transcriptase inhibitor (NRTI), protected women from HIV-1 acquisition with an overall efficacy of 39% and 54% observed in women who were high adherers 11 (i.e., reported to use the gel more than 80% of the time).
Effective prevention strategies involving blockade of HIV-1 mucosal transmission by using topical compounds such as microbicides will afford women the ability to protect themselves with a method that may also serve as a contraceptive. These agents were described initially as “virucides” because the first compounds evaluated such as detergents and acid buffering agents acted directly on the viral particles in a nonspecific manner by disrupting the viral envelope. 12 The name now commonly accepted for such products is “microbicides,” reflecting their ability to offer protection not only against viruses, such as HIV-1 and genital herpes, but also against bacteria that could modulate HIV-1 transmission, such as Gonorrhea or Chlamydia. 12 Although the term microbicides has been entrenched in the field, it does encompass specific antiretroviral agents that are also described as “topical PrEP.” Effective microbicide formulations must be safe to use, chemically and physically stable, compatible with latex and other materials used in barrier devices, affordable, and acceptable to the end user. They will be formulated as rings, gels, foams, creams, impregnated sponges, suppositories, or films, for insertion prior to sexual intercourse.
The recent disappointing failure of large-scale vaccines 13 and the need to improve microbicide efficiency 14 –17 revealed that major gaps in basic science remain to the development of effective microbicide formulations. 18 Indeed, it seems to be difficult to translate in vitro and animal studies to clinical outcomes. 19 We will here review current hypotheses concerning the crossing of HIV-1 through the genital mucosa with the aim of developing a design for microbicide formulations.
Transport of HIV-1 from the Female Genital Tract Toward the Lymph Nodes
Details concerning HIV-1 mucosal transmission have not been determined in humans but are essential for the design of effective vaginal microbicides. In the inferior female genital tract, cervicovaginal mucosal tissues include the columnar monolayer epithelium of the endocervix and the squamous multilayer epithelium of the vagina and ectocervix. The monolayer zone of transformation (between ectocervix and endocervix) and endocervix were described as potential mucosal sites of HIV entrance. 20,21 Recently, noninvasive imaging combining serial single-photon emission-computed tomography (SPECT/CT) and magnetic resonance (MR) analysis showed a radiolabeled cell-free and cell-associated HIV-1 surrogate distribution with the highest intensity in the vaginal pericervical area, without detectable signal in the uterus. 22 However, these are not the exclusive site of entry, as SIV has been transmitted to hysterectomized animals and HIV-1 in the congenital absence of a cervix. 21 Since women with HIV-1 detected in their cervical secretions were shown to have a thinner cervical epithelium and lower number of cellular layers, 23 we cannot exclude the possibility that exposure to HIV-1 impairs mucosal epithelial integrity. 24
The mechanism by which the virus crosses a tight layer of epithelial cells is still not known, and five potential mechanisms were thought to mediate HIV-1 mucosal transmission. These mechanisms included direct infection of epithelial cells, rapid transcellular transport through epithelial cells or “transcytosis,” epithelial transmigration of infected donor cells, uptake by intraepithelial dendritic cells, and circumvention of the epithelial barrier through physical breaches. 25 Since there is no evidence in vivo of transient infection of epithelial cells, 26,27 HIV-1 may cross a monolayer of epithelial cells by transcytosis, as suggested by experiments showing a link between protection of macaques against vaginal challenges and local production of antibodies that specifically inhibit HIV-1 transcytosis. 28 However, the extent to which these individual mechanisms contribute to mucosal HIV-1 infection is a matter of intensive investigation. A clear understanding of early events in sexual transmission, the immunological microenvironment, the target cells, and the type of transmitted viruses indeed constitutes the foundation for rationally designing products able to block specific viral entry pathways.
Role of genital inflammation in facilitating HIV-1 transmission
In macaques, SHIVSF162P3 vaginal inoculation was followed by proinflammatory cytokine production in cervicovaginal lavages (CVL) and plasma 29 and recruitment of CD4+ T cells that were necessary for mucosal viral spread and dissemination to the systemic compartment. 30 Notably, elevated genital cytokine concentrations, such as tumor necrosis factor (TNF)-α, interleukin (IL)-1β, IL-6, and IL-8, were associated with higher viral load set points, replication and shedding of HIV, and CD4 depletion, suggesting that strategies aimed at reducing genital inflammation may limit disease progression. 31 The magnitude of cytokine induction in CVL seems to be higher in infected macaque when compared to uninfected macaques. 29 Concentrations of cytokines/chemokines in cervicovaginal fluids may also be regulated by pregnancy and the menstrual cycle (e.g., IL-1β, IL-10, IL-6, and IL-8) and/or bacterial vaginosis and sexually transmitted infections (STIs) [Nesseiria gonorrhoeae, Chamydia trachomatis, Trichomonas vaginalis, Candida albicans, Mycoplasma hominis, Peptostreptococcus ascharolyticus, or herpes simplex virus-2 (HSV-2)]. 32,33 However, the number and distribution of immune cells in the cervicovaginal mucosa were found to remain constant throughout the menstrual cycle, indicating that sex steroid hormones modulate the metabolic activity of immune cells that are already present in the tissue. 34,35
Interestingly, a window of most frequent transmission of virus was estimated in the late luteal phase when progesterone levels are high and when local immunity may be low, 36 supporting findings concerning the higher susceptibility to HIV in women during progesterone-dominated periods including pregnancy and contraceptive use. Indeed, progesterone is known to inhibit cytotoxic T lymphocytes (CTLs) and natural killer (NK) cells, decrease and alter glycosylation of IgG and IgA antibodies, modulate cytokine production, and up-regulate HIV-1 receptors at the surface of CD4+ T cells. 37,38 In multiple studies in humans and macaques, high progesterone levels were associated with a higher viral burden, increased viral shedding, 39,40 and therefore accelerated HIV-1 disease progression. 41
However, activation of the mucosal innate immune system is necessary to control early and local viral replication. Indeed, it was suggested that the lack of a protective effect of rDC-SIGN gel may be due to DC-SIGN binding to the virus, thereby reducing viral antigenic exposure to both innate and adaptive immunity, which lowered antigenic exposure and then the magnitude of early proinflammatory mucosal responses. 29 Several lines of evidence suggest an important role for specific polyfunctional T cell responses in immune control of HIV infection. 42 Nevertheless, the induction of HIV-specific CD8+ effector cells was not clearly related to a better control of genital HIV-1 shedding. 31 Several studies showed that by reducing the inoculum dose used in a mucosal challenge infection model, the time to the first positive peripheral plasma SIV RNA level was lengthened. 43 Furthermore, two recently published studies showed that the combined use of a microbicide and a T cell-based vaccine limited the early mucosal transmission of HIV-1, 44,45 suggesting that a vaccine established sufficient immunity for control of SHIV or SIVmac251 infection if assisted by a microbicide molecule that expands the potential window of vaccine effectiveness by slowing local virus expansion and replication.
Proinflammatory environment may be induced by interactions between epithelial cells and viral particles
Epithelial cells constitute the main cell population in the initial contact with HIV-1 during its crossing through mucosae. As the initial barrier against viral entry, epithelial cells may hamper the crossing of the virus by their intrinsic integrity and solidity. Indeed, the disruption of the reproductive tract epithelium, as observed with nonoxynol-9, may facilitate HIV infection and serve as a portal for human HIV transmission. 46 However, the intrinsic mucosal properties also seem to be important since the penetrability of the rhesus macaque mucosa to the virus was reported to be dependent on the origin of mucosal tissues. 47 Indeed, it was found that the rectal route requires the smallest quantities of virus to infect macaques, followed by the vaginal route and then the oral route, reflecting the risk order of sexual HIV-1 acquisition among humans. 47 Moreover, the tightness and the integrity of the epithelium change markedly during the menstrual cycle, 30 as reported by the enhancement of SIV vaginal transmission observed in progesterone pretreated macaques. Furthermore, the viral gp120 seems to alter the integrity of the mucosal epithelial barrier, which then allows virus translocation by disrupting tight junctions by a mechanism that is TNF-α dependent. 24 With the lowest tightness, single epithelial monolayers, such as the endocervix and rectal mucosae, may support rapid transcellular transport or “transcytosis” from the apical to basolateral pole of the cell, and thus provide viral access to the cells of the submucosa. We have previously reported that less than 1% of HIV-1 can cross through the epithelial monolayer by transcytosis. 48
Vaginal epithelial cells may also modulate HIV-1 transmission by secreting monokines or proinflammatory factors such as thymic stromal lymphopoietin (TSLP) that enhance the maturation, activation, or recruitment of HIV-sensitive submucosal cells. 49 We have shown that CCR5-tropic HIV-1 interacting closely with the apical pole of epithelial cells induced the basolateral release of macrophage-attracting chemokines and proinflammatory cytokines. 50 In agreement with our data, inflammatory responses (including IL-1β, TNF-α, IL-10, and IL-12) in genital secretions are found to be correlated with infection with multiple HIV-1 variants, suggesting that the virus may use epithelial cells to ensure its own spread. 31 Notably, a proinflammatory mucosal environment may modulate the expression of HIV receptors at the surface of submucosal target cells. 51,52 More recently, it was shown that glycerol monolaurate, a widely used antimicrobial compound with inhibitory activity against the production of MIP-3α and other proinflammatory cytokines, can inhibit in vitro mucosal innate and inflammatory responses to HIV-1 and SIV, and in vivo it can protect rhesus macaques from acute infection even after repeated intravaginal exposure to high doses of SIV. 53 Overall, this study is consistent with studies showing that the establishment of a systemic infection requires a local expansion of a small founder population of infected cells in the cervicovaginal mucosa. 54,55 Altogether, these findings highlight the necessity of developing microbicide molecules that would prevent the early replication of the virus in mucosal tissues.
Submucosal replication of HIV-1
Early after HIV exposure, a small population retained the virus in the submucosa; this event was followed by local replication, which then resulted in expansion of virus replication mediated by the inflammatory environment. 53 Langerhans cells (LCs), defined by the cell surface expression of CD1a and langerin, are found within the epithelium. CD11c+ DCs, CD68+ macrophages, and CD4+ T cells are localized at the intraepithelial level and beneath the epithelial layer in the submucosa. 56 –59 Infected DCs, LCs, and macrophages have been observed shortly after sexual transmission, 53,57,60 –63 strongly suggesting that these innate immune cells are initial targets for HIV-1. Thus, identification of cell receptors mediating virus attachment may be important to specifically prevent the initial infection of these HIV-1 target cells. Some lectins such as the mannose receptor (MR), the DC-specific intercellular adhesion molecule-grabbing nonintegrin (DC-SIGN; CD209), and the DC immunoreceptor (DCIR) were identified as attachment receptors for HIV-1 on specific DCs and macrophage subsets. 64 –66 In addition, attachment of HIV-1 on epithelial cells 50,67 and submucosal target cells (T cells, 68 macrophages, 69 and DCs) was found to be mediated by glycosaminoglycans (GAG) such as syndecan. 70 Several studies showed that syndecans together with other lectins or host proteins, by acting as attachment receptors, facilitate HIV-1 entry.
Some of these attachment receptors also mediate de novo virus production. Indeed, DC-SIGN, MR, and DCIR promoted infection of innate immune cells as well as transmission of HIV-1 to target T cells, while langerin prevented infection through viral degradation. 71,72 Therefore, the surface expression of specific lectins influences target cell infection as well as virus internalization, and may participate in the involvement of innate immune cells in HIV-1 transmission through mucosae. Gringhuis et al. demonstrated that virus production by DC required some defined innate signaling events. 73 The first signal resulted from the interaction between HIV-1 genomic RNA and Toll-like receptor 8 (TLR8) that then promoted transcription initiation from the integrated provirus. The second signal, mediated by the close interaction between HIV-1 glycoprotein gp120 and the C-type lectin DC-SIGN, was necessary to activate the kinase Raf-1 and to lead to transcription elongation and productive virus expression. Immature DCs (iDCs) in the mucosa are among the first cells that capture HIV-1 and play a critical role in the early dissemination of HIV-1 to CD4+ T cells following sexual transmission. 60,74 Blocking the interaction between HIV-1 and DC-SIGN may also prevent viral transmission.
We have previously demonstrated a synergistic activity between entry inhibitors and an RT inhibitor against CCR5-tropic HIV-1 infection, 75 suggesting that a combination of microbicide molecules would be a most promising strategy to block the viral cycle at different stages. 76 However, it remains very speculative to currently identify the best antiretroviral strategy to block the initial capture of HIV-1 by DCs, and, in addition, its transmission from DCs to CD4+ T cells and even its replication by DCs. 77
Role of innate immunity
During inflammatory responses, neutrophils are one of the first innate immune subsets to be recruited to the site of infection where they generate inflammatory responses. 78 Neutrophils were identified in the cervix from both HIV-infected and uninfected women. 79 A correlation was found between cervical neutrophil numbers and concentration of IL-1β, TNF-α, IL-10, and IL-6, suggesting that the HIV-induced increase of those cytokines in vaginal fluids is related to the activation of neutrophils. 80 Their role in mucosal HIV transmission needs to be further investigated.
Early stages of viral infections were also associated with local recruitment and activation of other effectors of innate immunity, i.e., NK cells and DCs. The maturation and homeostasis of DCs seem to be controlled by a crosstalk between DCs and NK cells. During contact between activated NK cells (aNK) and iDCs, HMGB1 was found to promote the maturation of iDCs and the induction of IL-12-dependent T-helper-1 responses. 81 HMGB1 could act as an alarmin, a danger signal to alert the innate immune system to initiate the host defense, promoting the global immune activation reported in HIV-1 patients. 82 Following infection of iDCs with HIV-1, DCs were no longer susceptible to NK-dependent IL-12 polarization, and thus were no longer able to induce a Th1 response, which constitutes a strategy used by the virus to escape to the immune system. In addition, NK-dependent DC maturation and survival were associated with increased production of the HIV-1 p24 antigen and expression of viral DNA by DCs. 81 Interestingly, significantly high levels of HMGB1 were released in the cervicovaginal secretions of HIV-1-infected or HSV-2-infected women, 83 suggesting that coinfection with HSV-2 may increase HIV-1 transmission. In addition, a positive correlation between HMGB1 concentrations and disease progression was found, 84 suggesting that HMGB1 plays a central role in the global immune activation observed in HIV-1 infection.
Following HIV capture, extensive in vitro and in vivo studies have shown that the transfer of the virus from DCs and/or LCs toward CD4+ T cells may have an important role in HIV transmission. 56,85 Indeed, Perreau et al. showed that the failure of a recent vaccine was due to the induction of the maturation of DCs by the vaccine, which was associated with better activation of CD4+ T cells by DCs, which in turn efficiently replicated HIV-1. 86 Transmission of the virus from DCs to CD4+ T cells seems to be mediated through a tight synapse by a DC-SIGN-dependent mechanism 87 or through exosome traffic. 88 In fact, exosomes are antigen-bearing vesicles released by DCs. They constitute a way for DCs that have captured antigen to transfer it to other DCs for presentation and to subsequently elicit efficient specific T cell responses. 89 Similarities in composition and size between exosomes and HIV particles as well as in their colocalization in DC compartments suggest that the development of a blocking reagent against this kind of transfer would be very difficult.
We and others found that the microenvironment may differentiate macrophages into distinct populations that would facilitate the dissemination of the virus and the emergence of reservoirs. An interferon (IFN)-γ-induced Th1 environment 52 and IL-13-induced Th2 environment 90 resulted in the differentiation of macrophages into subsets that archived HIV-1 within a latent stage and resulted in the persistence of the virus by either favoring the recruitment of HIV-1-sensitive T cells or enhancing the viral production by these recruited T cells. However, differentiated macrophages in the presence of the Th2 cytokine IL-4 dramatically enhance the spread of HIV by capturing, integrating, producing, and transferring (toward CCR4+ IL-13+ T cells) higher levels of HIV-1. 90 High levels of lipopolysaccharides (LPS) were found in the serum of HIV-1-infected patients. LPS-treated macrophages released high doses of IL-10 that dramatically decrease the activation of CD4+ T cells by a PD-1/PDL-1-dependent mechanism. 91 Taken together, these data suggest that the possible modulatory properties of microbicide formulations on mucosal innate immunity may, by itself, favor or decrease HIV replication. However, to our knowledge, no evaluation of the effect of microbicides on the antiviral function of cells on innate immunity has yet been proposed in the preclinical evaluation of microbicide formulations.
In spite of these recent advances in the understanding of the role of innate immunity in the control of HIV-1 replication, further work is needed to determine the role of other innate immune effectors such as NK T cells and γδ T cells. Indeed, γδ T cells may play an important role in mucosal protection against microorganisms. 92 These cells are also able to release Th1 and Th2 cytokines that may modulate the activation of CD4+ T cell responses. 93 γδ+ T cells located in cervicovaginal tissues were found to release anti-HIV factors such as RANTES, MIP-1α, and MIP-1β that may prevent SIV infection. 94 Notably, their frequency was up-regulated by the treatment of macaques with GM-CSF and heat shock proteins, suggesting that mucosal immunization with heat shock proteins to activate γδ + T cells may also be a promising strategy.
Factors Modulating HIV Transmission
The nature of the transmitting virus
The selection of single or limited variants during transmission across the cervical/vaginal mucosa is still not clear. It was suggested that the mucosa exerts a selection pressure on the transmitted/founder (T/F) virus or that a competitive selection among transmitted variants occurs during the crossing from the mucosa to the systemic compartment. 95 There are several challenges to the characterization of T/F virus properties including identification of acutely infected individuals, development of an optimal robust and validated set of models to generate T/F molecular clones, and development of sensitive in vitro assays to detect phenotypic differences that could impact transmission fitness in vivo. 96 Pseudoparticles expressing the viral envelope glycoprotein 97,98 found in viruses isolated weeks to month after infection 99 were used to study viral properties associated with mucosal transmission. At the mucosal level, an immune response against the virus may occur rapidly, 100 and induce mutations in the viral envelope leading to viral escape, suggesting that “early”-isolated viruses may differ from true T/F viruses. 101 However, a limited viral evolution prior to the peak of viremia was observed, 97 suggesting a finite window of potential vulnerability of HIV-1 to specific engineered antibodies in microbicide formulation. A recent study involving the analysis of HIV-1 variants of transmitter–recipient pairs indicated selection for a transmitting virus variant with a set of essential biological properties, such as fewer glycosylation sites, more compact envelope domains, and a histidine residue at position 12 of the leader sequence of the envelope protein, 102 and in some cases, enhanced sensitivity to neutralization by CD4 binding site monoclonal antibodies. 103
Several studies showed that T/F and chronic viruses used CD4 and CCR5 with equal efficiency, 104 whereas other studies showed that the overall replication efficiency of T/F viruses was significantly lower in monocyte-derived macrophages compared to prototypic "highly macrophage-tropic" virus strains. 105 Notably, these T/F viruses replicated more efficiently in CCR5+ CD4+ T cells than in macrophages. 106,107 Finally, a viral subpopulation is archived as proviral DNA in the female genital tract early in primary infection, suggesting that HIV-1 variants from the male donor are selected in the female mucosal site during male-to-female transmission of HIV-1. 108
Furthermore, R5 and X4 viruses were found to cross through a tight monolayer of epithelial cells with equal efficiency. 94 Contrary to SHIVHxBc2, which replicates only in macrophages, the T lymphocyte restricted SIVmac239-induced systemic infection in vaginally challenged macaques, 109 indicating that there is no correlation between the tropism of the virus and its capacity to replicate and to induce systemic infection in the macaque model. Since most of the HIV-1 sequences in semen are CCR5 tropic 110,111 and transmitting donors are mostly asymptomatic, it was expected that transmitting HIV-1 would be predominantly CCR5 tropic. Other studies have also shown that epithelial cells were able to specifically sequester X4 viruses and to likely promote the preferential transmission of R5 viruses. 48,112 Saba et al. found that productive infection of cervicovaginal T lymphocytes with R5 viruses is much more efficient than with X4 viruses, suggesting that HIV-1 gatekeeping mechanisms exist in the cervicovaginal mucosa, preventing X4-HIV-1 from replicating as efficiently as R5-HIV-1. 113 Interestingly, the diversity of viruses transmitted intravaginally seems to be more restricted than those transmitted through the intravenous route, suggesting that host mucosal factors are mainly implicated in this selection. 114
Altogether, these current data utilizing a large number of envelope constructs strongly suggest that the mucosal bottleneck in the vagina is not the result of selective transmission of viruses with highly efficient CD4 or CCR5 use or with increased efficiency of entry into particular CD4+ CCR5+ mucosal cell subsets. 104 Therefore, it is possible that the mucosal immune system acts on the selection of HIV-1 variants and that the selection of viral variants occurs at the regional draining lymph nodes after crossing mucosal tissues. Further studies with cell-associated SIV and semen in a macaque model would be useful in defining the pattern of cell-associated virus transmission compared to cell-free virus transmission, and the impact of semen on the selection of R5 variants in the mucosa and of its subsequent transmission.
In addition, it is still unclear whether infection occurs via a cell-associated or cell-free mechanism. Both cervicovaginal secretion and sperm contain HIV-1-infected T cells and macrophages that are able to produce viral particles. Notably, a study of four transmission pairs from a well-characterized cohort of recently HIV-infected men who have sex with men revealed that in all these cases, the virus establishing infection in the recipient originated from the seminal plasma, suggesting that cell-free rather than cell-associated viruses are the origin of sexually transmitted HIV-1. 115 In addition, transmission via cell-associated SIV was found to be much less efficient than cell-free virus. 116,117 However, in vivo experiments showed that infected cells reached to cross through vaginal epithelial cells. 118 In this study, cells from the spleen and not isolated from sperm were used to infect female macaques through the vaginal route. Cells from sperm and spleen may differentially express receptors such as those implicated in adhesion, viral attachment, and migration of cells, which may modulate their transmigration. Upon budding of the virus from host cells, viral particles incorporated cellular proteins such as ICAM-1, HLA-DR, CD40, CD40L, and CD86, and this enhanced their infectivity. 119 Thus, the composition of the surface viral particle appears to influence the selection and the transmission of the most replicative variants.
Further analyses are required to characterize the cells that are able to transmigrate and to define the source of the seminal plasma virus. In addition, the viral load in the ejaculate needed for productive mucosal infection is not known yet.
HIV modulating factors found in the female genital tract
Epithelial cells from the genital tract produce several biological factors including a hydrophilic surface layer of glycoproteins and glycolipids, called the glycocalyx, and thick hydrophobic glycoprotein mucus 120 that may act as an initial barrier. Cervicovaginal mucus obtained from donors with lactobacilli dominant flora efficiently trapped HIV-1. 120 Hydrogen peroxide-producing (H2O2 +) Lactobacillus, a key regulator of the vaginal ecosystem, was found to decrease HIV-1 replication mostly through direct effects, whereas lactic acid was found to suppress bacteria associated with bacterial vaginosis. 121 However, cervicovaginal fluids and semen showed significant H2O2-blocking activity. 122 Similar to inflammatory-associated diseases, bacterial vaginosis and diseases that alter epithelial integrity increased HIV-1 transmission. Microbicides and other topical applications prevented vaginal transmission of HIV, indicating that an efficient microbicide could preserve the vaginal flora. 123
Others antimicrobial compounds were produced such as secretory IgA or natural IgG antibodies and lactoferrin. Recently, it was found that following immunization and HIV reexposure, IgG and IgA antibody-secreting cells were significantly correlated with ADCC killing, ADCVI activity, and inhibition of transcytosis. 124 Upon reexposure to antigen, strong anamnestic antibody responses were detected with the rapid increase in antigen-specific circulating antibody levels, which was associated with weaker viral replication. However, some natural antibodies were found to enhance the capture of HIV-1 by dendritic cells via the Fc receptor, resulting in increased viral production, which constitutes a new way for the virus to use the immune system for own advantage. 125 The activity of these HIV-1-enhancing factors may be unbalanced by released mucosal antiretroviral factors, including RANTES, elafin, secretory leukocyte peptidase inhibitor (SLPI), histatins, statherin, lysozymes, and cystatins. Efficient microbicide formulations should not affect the abundance and/or effectiveness of these mucosal antiviral factors.
Several studies suggested that active immune regulatory mechanisms, rather than intrinsically attenuated innate immune responses, underlie the low levels of immune activation characteristic of resistance to HIV infection in humans 126 and in the macaque model. 127 Compared with HIV-unexposed individuals, it was reported that people who have been repeatedly exposed to HIV but are resistant to infection have reduced frequencies of activated T cells and elevated frequencies of regulatory CD4+ T cells (Treg). 126 A recently published paper suggests that a combination of a modest vaccine-induced CD8+ T cell response in the context of immunoregulatory suppression of T cell activation protected against vaginal HIV transmission. 128 This study highlighted the need to induce a protective adaptive immunity against the virus with a suppressive environment preventing virus replication in activated mucosal cells susceptible to HIV. 128 Moreover, the soluble factor PD-1 was able to negatively modulate immune activation by limiting the activation of both CD8+ T cells 129 and CD4+ T cells, 91 indicating its potential use as a therapeutic/preventive target. In addition, the release of plasma proapoptotic factors [such as TNF-related apoptosis-inducing ligand (TRAIL), Fas ligand, TNF receptor type 2 (TNFR-2), and microparticles] early after infection was shown to have a suppressive effect on T cells, B cells, and macrophages. 130 The release of products of cell death from mucosal cells and the subsequent immunosuppression following HIV-1 transmission could potentially narrow the window of opportunity to block HIV amplification in mucosal tissues. 130 A comparison of the nonpathogenic and pathogenic models also revealed other factors that may contribute to regulating immune activation, including (1) IDO, an immunosuppressive molecule produced by macrophages and DCs regulating the proliferation of T cells and antagonizing IFN activity, 131 (2) ADAR, an adenosine deaminase-suppressing IFN-stimulated gene (ISG) expression, 127,132 and (3) TIM3, a marker of T cell exhaustion, and its ligand galectin-9, a proapoptotic ISG. 133 Therefore, the activation of mucosal immunoregulatory mechanisms constitutes a new area of investigation to prevent early events in HIV-1 mucosal transmission.
HIV-1-inducing factors found in sperm
It has been reported that semen may enhance HIV infection. 134 HIV-1 concentrations in the semen are one of the predictive factors of HIV-1 transmission. Semen contains cell-free and cell-associated HIV-1 and then represents the main vector of HIV-1 dissemination. Within hours of coitus, macrophages, DCs, granulocytes, neutrophils, and memory CD8+ T cells are recruited into the endometrial stroma and lumen. 135 An accompanying increase in GM-CSF, IL-6, IL-8, and IL-1 was also induced in cervical tissues, which is consistent with seminal-plasma-induced cytokines acting to control leukocyte recruitment and activation in the female genital tract. 136,137 Indeed, seminal TGF-β was shown to stimulate GM-CSF production and inflammatory cell recruitment in the murine uterus. 138 In all semen samples from healthy men, IL-8, IL-7, SDF-1a, and MCP1 were present in high concentrations whereas IL-5, IL-6, IL-13, IL-17, IFN-α, RANTES, MIP-1β, IL-1α, IL-1α, G-CSF, and M-CSF were detected at low concentrations. 139 MIP-1α, IL-2, IL-10, IL-12, TNF-α, IFN-γ, and GM-CSF are detected in a minority of semen samples. 139
Some cytokines activated regulatory T cell responses, such as TGF-β, which is known to facilitate preparation of the female reproductive tissues for pregnancy.
140
This regulatory property may limit HIV replication. However, it has also been shown that TGF-β can potentiate the recruitment of immature CCR5+ Langerhans cells and can then facilitate the transport of viruses through epithelial cells toward the lymph nodes.
141
These studies suggest that as a first step, proinflammatory factors found in semen and those induced in the female genital tract following HIV-1 exposure may facilitate recruitment of HIV-1 target cells and their subsequent infection. In addition, they indicate that as a second step, the local activation of immunosuppressive mechanisms may limit dissemination of the virus as well as the induction of a protective immune response, facilitating the escape of several viral strains and their subsequent dissemination. Cytokines may also activate the local replication of the virus in mucosal target cells (dendritic cells, macrophages, and T cells) soon after its crossing through the vagina. To block the replication of HIV-1 in the first cells infected in mucosal tissues, RTIs have demonstrated efficacy as microbicides.
142
Compared to NRTIs, non-NRTIs (NNRTIs) seem to be more potent, in part because of their long half-life due to their capacity to bind irreversibly to RT, disrupting the binding of substrates to the active site of the enzyme. Recent examples of NNRTIs in clinical evaluations as microbicide candidates include dapivirine and UC781 (
In addition to free viruses and infected leukocytes, spermatozoa-associated virions are also one of the major sources of infectious virus in semen. Heparin sulfate moieties, expressed in spermatozoa, were found to play an important role in the capture of HIV-1 and to mediate efficiently the transmission of HIV-1 to DCs, macrophages, and T cells. 144 Notably, at low values of extracellular pH, similar to those found in the vaginal mucosa after sexual intercourse, the binding of HIV-1 to the spermatozoa and the consequent transmission of HIV-1 to DCs were strongly enhanced, suggesting that spermatozoa may act in the early transmission of HIV-1 infection. In addition, opsonization of free HIV-1 particles by complement components found in semen enhanced in vitro infection of macrophages and DCs and also the viral transfer to CD4+ T cells. 50 We have recently shown that HIV-1 opsonization may modulate in vitro the efficiency of candidate microbicides to inhibit HIV-1 infection of mucosal target cells, as well as its crossing through the mucosa. 145 Sperm contains other HIV-1-inducing proteins such as fibril-forming prostatic acidic phosphatase (PAP), 134 basic polyamines (spermin and spermidin), and free nucleotides (dNTP) that may protect HIV-1 virions from the threat of acid inactivation in the vaginal tract and enhance the transcription of viruses or TGF-β1.
However, sperm also contains antimicrobial compounds such as SLPI, 146 β-defensins, and natural antibodies against DC-SIGN or CCR5. 51,87 Further work is needed to identify all these semen-activating factors, which will then provide an alternate strategy to limit HIV-1 transmission by specifically blocking all these HIV-1-inducing factors.
Conclusions
Well-defined mechanisms of HIV-1 mucosal transmission are an essential component of rational microbicide development. Despite years of basic science, methods by which HIV-1 crosses through epithelial cells, identification of HIV-1 target cells that replicate the virus locally in the female mucosal tissue, and characterization of cells that disseminate the virus are still not well defined (Fig. 1).
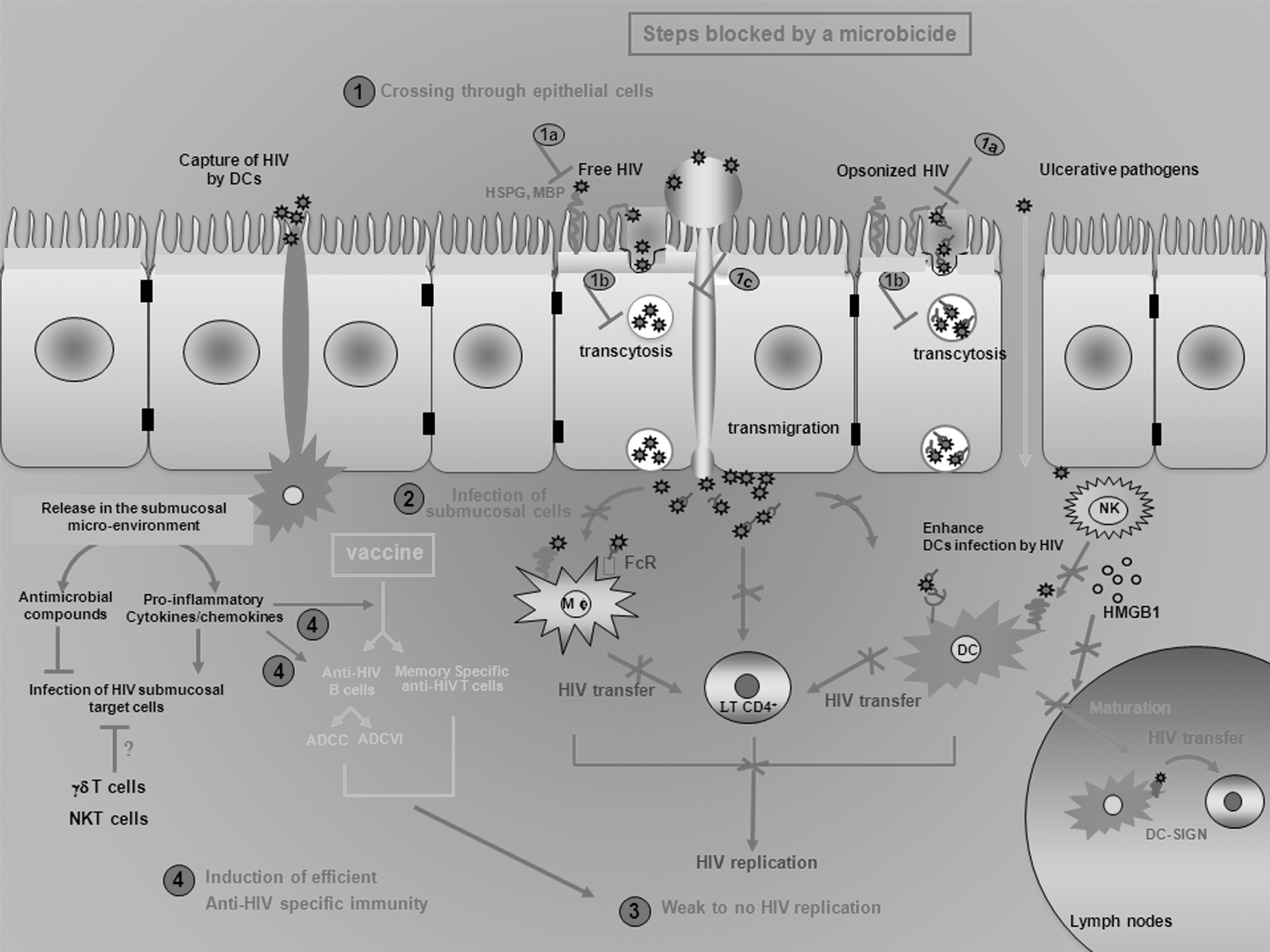
Microbicides would delay mucosal viral replication or decrease the infectious dose of the virus, which helps the induction of efficient and specific anti-HIV mucosal responses. Five potential mechanisms are thought to mediate HIV-1 mucosal transmission: direct infection of epithelial cells, transcytosis, epithelial transmigration, uptake by intraepithelial dendritic cells, and circumvention of the epithelial barrier through physical breaches. Mucosal crossing of HIV and further reaching of submucosal HIV-sensitive cells may be modulated by supraepithelial factors, such as seminal complement component (opsonized HIV), and by epithelial factors released in the submucosal environment, such as antimicrobial factors, cytokines, and chemokines, and by the submucosal innate immunity. Once HIV-1 has reached the submucosal environment, numerous HIV target cells may support the local replication of the virus [macrophages (Mϕ), DCs, and CD4+ T cells]. This HIV replication may be dramatically increased by several cells of the innate immune system (neutrophils and NK cells). More specifically, NK cells may promote viral dissemination by enhancing HIV replication by DCs and their migration to the lymph nodes where they may transfer the virus to T cells. To prevent HIV mucosal transmission, a microbicide would act on the HIV crossing through mucosae (1) by limiting its attachment of the virus to epithelial cells (1a) and/or their migration (1b and 1c), but also on the local replication (3) by inhibiting infection of submucosal cells (2). If the microbicide is not 100% efficient, the delay of viral replication would be efficient to the activation of the adaptive immune system. Cytokines and chemokines released by HIV-activated innate cells may elicit the induction of anti-HIV CD4+ and CD8+ T cells, and the release of specific antibodies that may neutralize directly the virus or induce antibody-dependent cell-mediated cytotoxicity (ADCC) and antibody-dependent cell-mediated viral inhibition (ADCVI). The time of induction of these anti-HIV-specific responses may be shortened by a prevaccination of individuals that activates efficient memory-specific responses. However, there is no HIV vaccine currently available and poor understanding of the subtle orchestration of the numerous viruses, cell factors, and HIV target cells involved in HIV mucosal transmission renders the design of an effective microbicide formulation difficult. DC-SIGN, dendritic cell-specific intercellular adhesion molecule-3-grabbing nonintegrin; MBP, mannose-binding protein; NK, natural killer; NKT, natural killer T cells.
In women, technical challenges associated with obtaining genital tissue limit our understanding of the important role of the cervicovaginal environment in the induction of innate and adaptive immunity to prevent HIV-1 infection. Invasive biopsy approaches to isolate immune cells from mucosal tissues and approaches for sampling mucosal tissues from the genital tract, such as cervical lavages and cervical cytobrushes, are not sufficient to explore details of HIV-1 mucosal transmission. Recently, Louissaint and colleagues demonstrated the feasibility of using radiolabeled HIV-free or cell-associated particles to determine the distribution of HIV-1 and semen surrogates after simulated intercourse without disrupting the vaginal contents and inducing local inflammation. 22
Several studies showed clearly that cervicovaginal and seminal factors may modulate transmission of the virus. It seems that the presence of high levels of proinflammatory cytokines/chemokines in mucosal tissues may favor replication of the virus and is correlated with disease progression. However, induction of anti-HIV-specific responses needs the activation of innate effectors that release proinflammatory monokines. It will then be a challenge to differentiate the proinflammatory response that facilitates the initial viral seeding/spread and the proinflammatory response resulting from the activation of the innate immune system that elicits specific antiviral responses. In the macaque model, the reduction of mucosal T cell activation by immunosuppressive mechanisms combined with a modest vaccine-induced CD8+ T cell response would allow the antiviral CD8+ T cells to control mucosal SIV replication before the infection could disseminate. 128 These latter findings suggest that an HIV-induced proinflammatory environment constitutes the first step in the local amplification of the T/F viruses, which would help its escape from the mucosal immune system induced more recently in a second step (emergence of new virus variants, infection of the cellular reservoir, and migration of infected cells from mucosal tissues to lymph nodes).
A recent proof of concept study also demonstrates that the combined use of a microbicide to lower the initial viral spread and a vaccine to elicit a strong mucosal immune response holds promise as a preventive strategy to limit HIV-1 mucosal transmission. 44 Other studies raised the possibility of combining microbicide molecules, which would (1) decrease the toxicity of each compound and their adverse effects, (2) have potential synergistic activity against distinct and complementary pathways used by HIV-1 to disseminate, (3) decrease the risk of selection of antiretroviral drug-resistant HIV-1 strains, and (4) be more advantageous not only against HIV-1 but also against other pathogens (such as genital herpes). 75,147 –149 Since gels containing nonspecific compound (such as polyanion-based microbicides and buffering agents) showed modest potency in phase III efficacy trials, 16,17 product development has recently turned to microbicides containing highly potent HIV-specific antiretroviral compounds, including entry inhibitors and RT inhibitors. Thus, several entry inhibitors under development as microbicide candidates work to block the CCR5 chemokine receptor, such as the CCR5 antagonists maraviroc and L-167, 150,151 and the specific interaction between gp120 and CD4, such as the Cyanovirin-N lectin, the L-644 peptide, and the BMS-378806 compound. 152,153
CCR5 antagonists may be more efficient in blocking the transmucosal crossing of CCR5-tropic HIV-1 strains, which are considered to be the primary viruses to be transmitted, due to their mucosal selection and their concentration within the genital secretions. 25 Based on this, CCR5 antagonists could constitute another very efficient mucosal barrier against the virus, acting synergistically with the other physiological existing barriers. However, in addition to CCR5-blocking agents, the RT inhibitors blocking the conversion of viral RNA to proviral DNA, and then preventing the infection of target cells and the subsequent establishment of reservoirs, may also be used as possible microbicides, 142 acting after cell entry in a more classical antiviral process. Both CCR5 antagonists and antiretroviral molecules targeting the HIV replication cycle are likely to be complementary and could be associated. Further studies are required to compare the effectiveness of several combinations of topical microbicides with partially effective preventive strategies (male circumcision, treatment of curable STIs, and use of antiretroviral drugs as prevention).
Footnotes
Author Disclosure Statement
No competing financial interests exist.