
Abstract
Vitamin D3 is known to have an effect on the immune function. We investigated the immunomodulatory capability of vitamin D3 in HIV-infected patients and studied the expression of chemokine receptors on regulatory T cells (Treg). Vitamin D3-deficient HIV-1-seropositive subjects were treated with cholecalciferol (vitamin D3) at a dose of 800 IU daily for 3 months (n=9) or 25,000 IU weekly for 2 months (n=7). Peripheral blood mononuclear cells (PBMCs) were isolated and analyzed for skin-homing (CCR4 and CCR10) and gut-homing (CCR9 and integrin α4β7) marker expression on Treg, by flow cytometry, before and after supplementation. Serum 25(OH)D3 and parathyroid hormone (PTH) levels were determined at baseline and after the treatment period. Weekly doses of 25,000 IU cholecalciferol effectively achieved the optimal target serum 25(OH)D3 concentration of >75 nmol/liter (30 ng/ml) in HIV-infected patients. High-dose cholecalciferol supplementation differentially influenced skin-homing markers on Treg with an increased level of CCR10 expression and while a reduction in CCR4 expression level was observed together with a lower percentage of Treg expressing CCR4. For both dosing regimens, there were no significant differences in the expression of gut-homing markers, CCR9, and integrin α4β7. High-dose vitamin D3 supplementation is needed to reverse vitamin D3 deficiency in HIV-infected individuals and this results in modulation of skin-homing markers but not gut-homing markers expression on Treg. At a standard dose of 800 IU/day, vitamin D3 is not effective in achieving an optimal 25(OH)D3 concentration in patients with an underlying T cell dysfunction and is unable to exert any immunomodulatory effects.
Introduction
V
Human immunodeficiency virus-1 infection is characterized by progressive CD4+ T cell depletion and immune dysfunction. Regulatory T cells are potential targets of HIV given that they express CD4. 15 HIV entry into target cells also requires cellular expression of the chemokine receptors CCR5 and CXCR4 in conjunction with CD4. 16 The role of Treg in regulating T cell activation during immune responses to pathogens such as chronic viral infections is a subject of great interest. Their effects can be beneficial or detrimental hinging on the balance between attenuating HIV-induced immune hyperactivation and mounting an immune response to HIV and mucosal pathogens. Chronic HIV infection also seems to alter the distribution of Tregs resulting in an apparent increased proportion in the peripheral lymph nodes and mucosal lymphoid tissues. 17 –20 Various chemokine receptors such as CCR4, CCR9, CCR10, CD62L, and integrin α4β7 are known to dictate the homing capacity of T cells as well as Tregs. 21,22 Using CD4+CD25+ Tregs isolated from healthy subjects, Ji et al. demonstrated that HIV-1 binds to Treg leading to an up-regulation of CD62L and integrin α4β7 expression. 20 As a result, Tregs migrate to and accumulate in the peripheral lymph nodes and mucosal lymphoid tissues.
Vitamin D3 has been also shown to affect the homing capacity of the peripheral CD4+ T cell population. Sigmundsdottir et al. reported that 1,25(OH)2D3 induced the expression of skin-homing chemokine receptors, CCR10 on CD4+ T cells in vitro, at the expense of gut-homing markers, CCR9 and integrin α4β7. 23 Given that both vitamin D3 23 and HIV 17,18,20 can modulate the expression of chemokine receptors on Treg, it leads to the question of whether the homing capability of Treg can be augmented by vitamin D3 during HIV infection. We investigate how cholecalciferol (vitamin D3) in two different dosing regimen, 800 IU daily versus 25,000 IU weekly, can impact Treg homeostasis in HIV-infected individuals and evaluate the expression of skin- and gut-homing markers in vivo and in vitro.
Materials and Methods
Patients
From April 2007 to August 2009, we conducted a pilot study in vitamin D3-deficient HIV-infected patients visiting the Radboud University Nijmegen Medical Centre, The Netherlands. Patients included were HIV-1 seropositive, vitamin D3 deficient, and above 18 years old. Vitamin D3 deficiency was defined as 25(OH)D3 levels below 25 nmol/liter (10 ng/ml). The low-dose study cohort (n=9) received a daily oral dose of 800 IU cholecalciferol for 3 months. The high-dose cohort (n=7) received a weekly oral dose of 25,000 IU cholecalciferol for 2 months. In the control group (n=7) recruited from February to May 2009 and followed through until May to August 2009, no supplementation was administered. At baseline and after the treatment period, blood samples were taken for peripheral blood mononuclear cell (PBMC) isolation preceding flow cytometry analysis and serum 25(OH)D3 levels. The study was approved by the Ethical Committee on Human Experimentation of the Radboud University Nijmegen. A written consent was obtained from all participants to the study. PBMCs were isolated as described below and subsequently used for flow cytometry analysis.
In vitro study
Buffy coats were obtained from healthy donors (Sanquin Blood Bank, Region South East, the Netherlands). Informed consent was obtained according to the Declaration of Helsinki. PBMCs were isolated by density centrifugation with Lymphoprep (Axis-Shield AS, Oslo, Norway) and LeucoSep (Greiner Bio-One, Frickenhausen, Germany). CD4+ T cells were purified from PBMCs by negative selection using monoclonal antibodies (mAbs) directed against CD8 (RPA-T8), CD14 (M5E2), CD16 (3G8), CD19 (4G7), CD33 (P67.6), CD56 (B159), and CD235a (GA-R2(HIR2) (BD-Biosciences, Erembodegem, Belgium) combined with sheep-antimouse-Ig-coated magnetic beads (Dynal Biotech, Invitrogen ASA, Oslo, Norway). The resultant CD4+ T cell fraction, typically of >90% purity, was incubated with anti-CD25-PE (M-A251, BD Biosciences, NY), anti-CD4-ECD (SFCI12T4D11), and anti-CD27-PC5 (1A4CD27)-conjugated antibodies (both from Beckman Coulter Corporation, Miami, FL). CD4+CD25hiCD27+ Treg were isolated from purified CD4+ T cells by high-purity flow cytometric cell sorting (Altra Flow Cytometer, Beckman Coulter, Miami, FL). The isolated CD4+CD25hiCD27+ Treg routine yield of >95% purity were stimulated with anti-CD3/anti-CD28 mAb-coated T cell expander beads (Dynal Biotech, Invitrogen ASA, Oslo, Norway) in 200 μl of culture medium (RPMI-1640 supplemented with glutamax, 0.02 mM sodium pyruvate, 100 U/ml penicillin, 100 μg/ml streptomycin) (all from Gibco, Paisley, UK) and 10% human pooled serum (HPS). Exogenous recombinant human interleukin-2 (rhIL-2) 12.5 U/ml (Proleukine, Chiron, Amsterdam, the Netherlands) was added to the cell culture. 1,25(OH)2D3 (Fluka Biochemika, Sigma-Aldrich, MO) was dissolved in absolute ethanol and made up to a concentration of 100 nM in each well. No solvent effect was apparent in control experiments. Cell cultures were performed in 96-well round bottom plates (Greiner, Frickenhausen, Germany) and incubated for 3–4 days in a 37°C, 95% humidity, 5% CO2 incubator.
Flow cytometry
Cells were phenotypically analyzed by five-color flow cytometry (Coulter Cytomics FC 500, Beckman Coulter, Fullerton, CA) using Coulter Epics Expo 32 software. Cells were washed with phosphate-buffered saline (PBS) with 0.2% bovine serum albumin (BSA) before being labeled with fluorochrome-conjugated mAbs. After incubation for 20 min at room temperature, in the dark, cells were washed twice to remove unbound antibodies and analyzed. For cell surface staining, the following mAbs were used: CD127 PC5-labeled or PC7-labeled (RDR5) (both from eBioscience); CD25-PE (M-A251), CD25-APC (2A3), CD49d-PE (9F10), and CCR4-PC7 (1G1) (all from BD Biosciences); CD4 ECD-labeled or PC7-labeled (SFCI12T4D11), CD4-PC5 (13B8.2), and CD27-PC5 (1A4CD27) (all from Beckman Coulter Corporation); CCR9-PE (112509) and CCR10-PE (314305) (both from R&D Systems); and CD27-FITC (M-T271; Dako). Appropriate isotype control mAbs were used for marker settings.
25(OH)D3 and parathyroid hormone levels
Serum 25(OH)D3 was measured by high-pressure liquid chromatography (HPLC) with UV detection, after prior extraction on small SepPak columns as previously described. 4 Tritiated 25(OH)D3, collected from the HPLC system during passage of the UV peak, was used to correct for procedural losses. The within run precision was 2.6% at 69 nmol/liter and the between run precision was 6.2% at 69 nmol/liter. For parathyroid hormone (PTH) assays the ELSA-PTH assay by CIS BIO was used at first. This method was switched into the intact PTH assay performed by the Abbott Architect analyzer when the CIS BIO assay was no longer available. The Abbott Architect assay was recalibrated on the CIS BIO assay to give identical measurement results.
Statistical analysis
Statistical analysis was performed using the Statistical Product and Services Solutions package version 16.0. The Wilcoxon rank-sum test was used to compare differences between groups (unless otherwise stated). The level of significance was set at p≤0.05.
Results
We investigated the effect of low-dose versus high-dose cholecalciferol (vitamin D3) on serum 25(OH)D3 and PTH levels in HIV-infected subjects. There were three study groups: the high-dose cohort (n=7) received weekly oral cholecalciferol supplementation at a dose of 25,000 IU for 2 months, the low-dose treatment arm (n=9) was given 3 months of 800 IU/day cholecalciferol supplementation orally, and the control group (n=7) received no supplementation and was followed up from spring to summer. The patients' characteristics are presented in Table 1.
High weekly dose of 25,000 IU cholecalciferol achieves the optimal target serum 25(OH)D3 concentration in HIV-infected patients
The baseline median serum 25(OH)D3 and PTH levels in the high-dose cohort (n=7) were 30.0 nmol/liter and 7.7 pmol/liter, respectively. After weekly cholecalciferol supplementation at 25,000 IU for 2 months, the median serum 25(OH)D3 and PTH levels were 138.0 nmol/liter and 6.7 pmol/liter, respectively (Fig. 1A). In the low-dose cohort (n=9), the baseline median serum 25(OH)D3 and PTH levels were 23.8 nmol/liter and 7.2 pmol/liter, respectively. After 3 months of 800 IU/day cholecalciferol supplementation, the median serum 25(OH)D3 and PTH levels were 38.7 nmol/liter and 3.8 pmol/liter, respectively (Fig. 1B). The optimal target serum 25(OH)D3 of 75 nmol/liter was achieved in 86% (6/7) of the high-dose study cohort but only two subjects (22%) in the low-dose treatment group attained this target concentration.

Serum 25-hydroxyvitamin D3 [25(OH)D3] concentrations and parathyroid hormone (PTH) levels. Serum 25(OH)D3 and PTH concentrations in
From February to May 2009, seven HIV-infected subjects were recruited and no vitamin D3 supplementation was given to this group. They were monitored for the effect of seasonal variation of vitamin D3 status after 2 to 3 months. In spring, the median serum 25(OH)D3 and PTH levels were 33.0 nmol/liter and 6.6 pmol/liter, respectively, and in summer, the median serum 25(OH)D3 and PTH levels were 48.5 nmol/liter and 5.7 pmol/liter, respectively (Fig. 1C).
None of our patients was obese and this precludes BMI as a possible confounder from the analysis of serum 25(OH)D3 levels. We also found no association between patient's CD4 counts and serum 25(OH)D3 levels.
Modulation in the skin-homing capacity of Treg can be observed only in high-dose vitamin D3 supplementation
In the next set of experiments, we investigated whether vitamin D3 supplementation can modulate the homing capability of Treg in HIV-infected patients. For this purpose, we evaluated the skin-homing markers, CCR4 and CCR10, as well as gut-homing markers, CCR9 and integrin α4β7, using flow cytometry (Fig. 2).
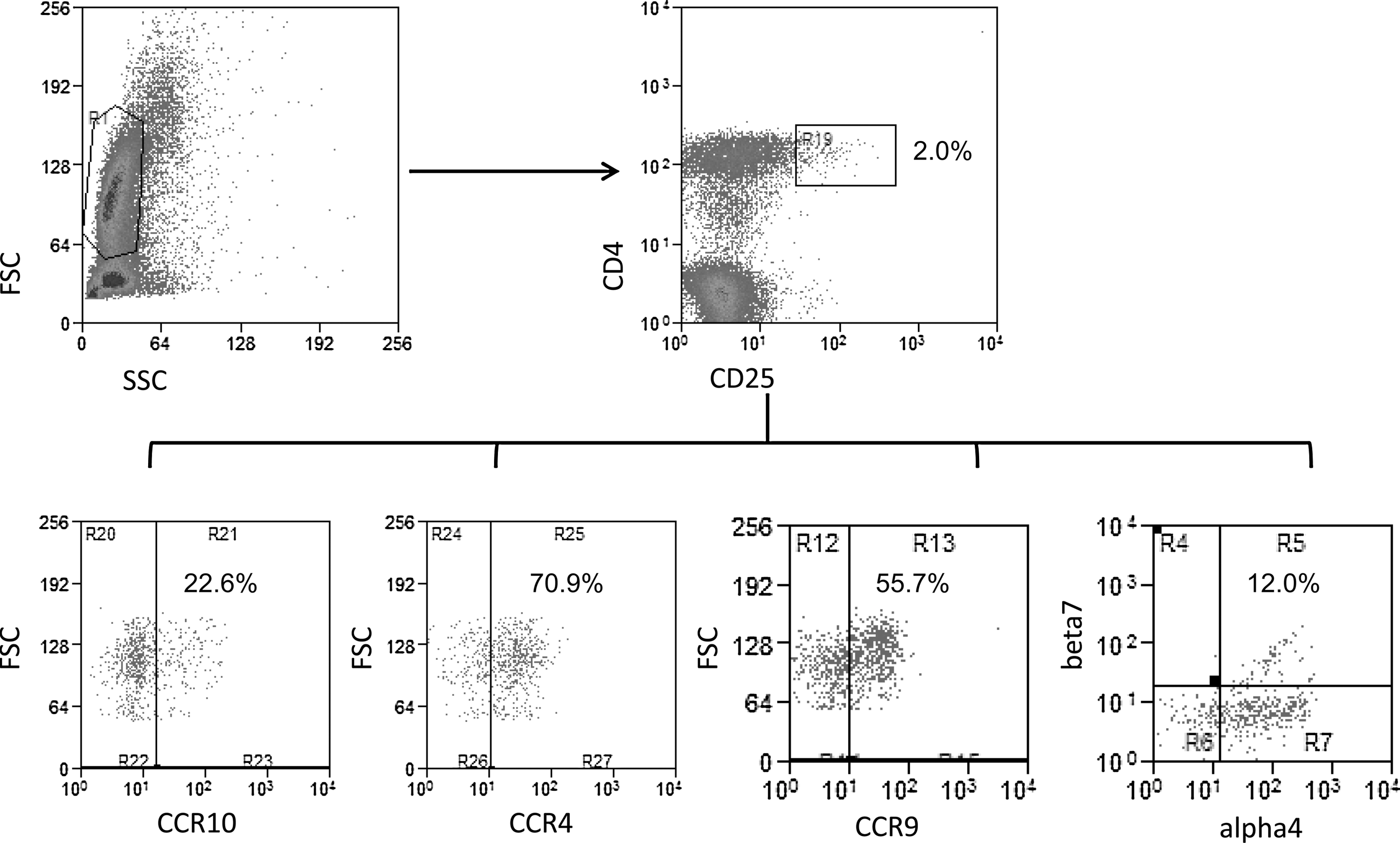
Representative flow cytometry dot plots for homing markers expression in HIV-infected patients treated with high-dose cholecalciferol. Representative dot plots show surface staining for markers as analyzed on peripheral blood mononuclear cells (PBMCs). The corresponding markers are indicated along the x-axis and the y-axis. Cells were first gated from the FSC (forward angle scatter) vs. the SSC (right-angle scatter) plot and subsequently the CD4+ and CD25high regulatory T cells were further analyzed for homing markers (CCR10, CCR4, CCR9, and integrin α4β7) as shown in the lower panel. The percentages are indicative of the cell population in the respective quadrants.
A significant increase in CCR10 expression (mean fluorescence intensity, MFI) by Treg was seen in the high-dose cohort after treatment (Fig. 3A). In the same study group, a decrease in the skin-homing chemokine receptor, CCR4 expression level (MFI) was observed together with a lower percentage of Tregs expressing CCR4 (Fig. 3C). However, in the low-dose treatment group no change in both skin-homing markers could be observed after 3 months (data not shown).
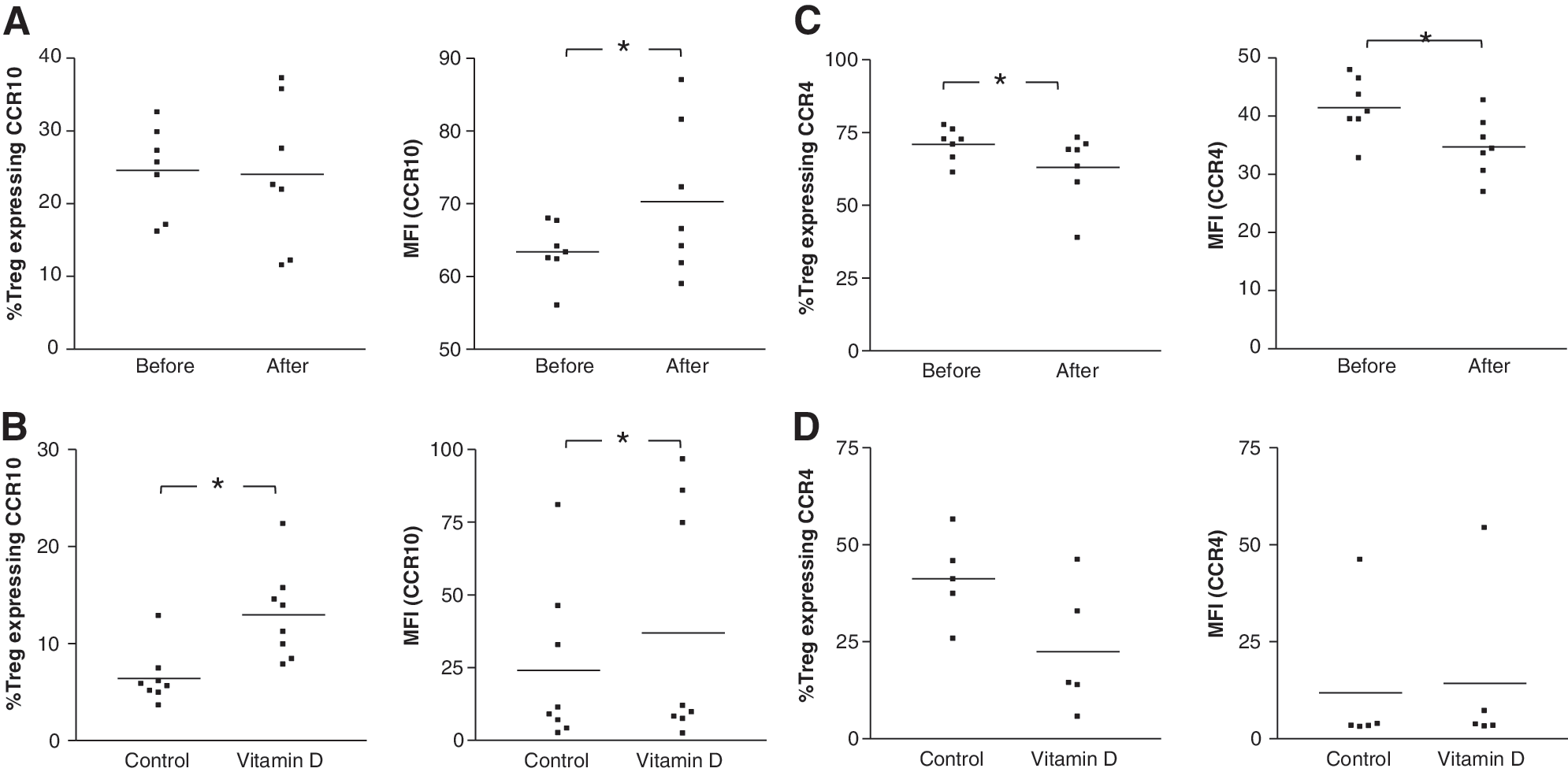
In vivo and in vitro effects of vitamin D3 on skin-homing markers CCR10 and CCR4 expression by regulatory T cells. Percentage and level of expression (mean fluorescence intensity, MFI) of skin-homing marker
We verified our in vivo findings by performing the analysis on regulatory T cells isolated from healthy volunteers and tested the in vitro effects of vitamin D3 using a supraphysiological dose of 1,25(OH)2D3. Treg were stimulated with anti-CD3/anti-CD28 mAb-coated microbeads in the presence of exogenously added rhIL-2 and 100 nM 1,25(OH)2D3 and analyzed after an incubation period of 4 days. The percentage of Tregs expressing CCR10 was significantly increased (Fig. 3B). A significantly elevated level of expression of CCR10 by Treg was also apparent. On the other hand, there was no significant effect observed for the expression of CCR4 in vitro (Fig. 3D).
Vitamin D3 has no effect on gut-homing markers in vivo and in vitro
There were no changes in the frequency and expression levels of the gut-homing markers CCR9 and integrin α4β7 with either low daily dose (data not shown) or high-dose pulsed therapy with vitamin D3 in HIV-infected patients (Fig. 4A and C). Similarly, we looked at the expression of these markers in vitro, using Tregs isolated from healthy volunteers and stimulating these cells with anti-CD3/anti-CD28 mAb-coated microbeads and tested the effect of 100 nM 1,25(OH)2D3. Parallel to our in vivo observation, vitamin D3 has no effect on CCR9 and integrin α4β7 expression in vitro (Fig. 4B and D).
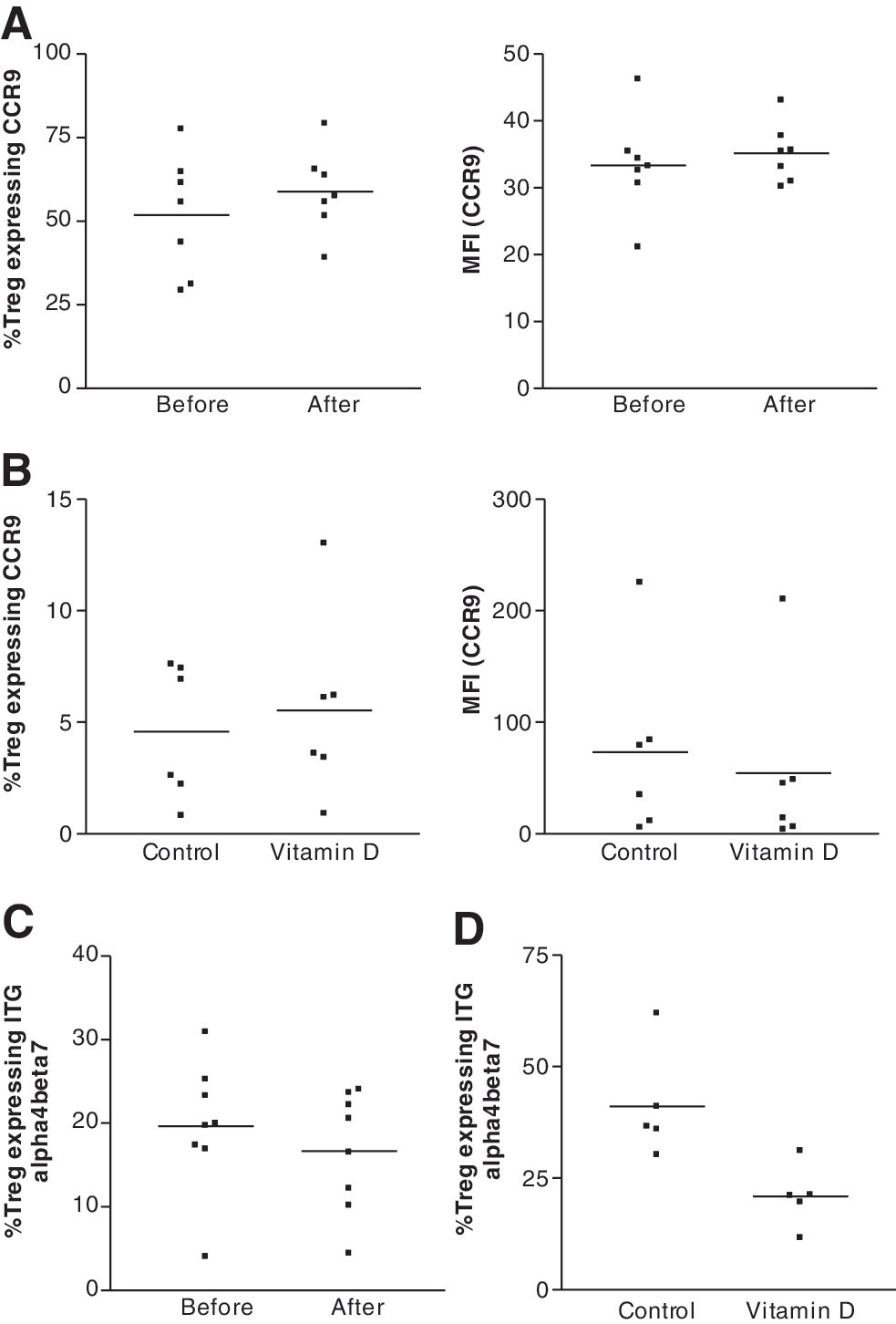
Effects of vitamin D3 on gut-homing marker CCR9 and integrin α4β7 expression by regulatory T cells in vivo and in vitro. Percentage of Treg expressing
Discussion
In this study, we used two different doses of vitamin D3 supplementation to investigate its effect on regulatory T cell trafficking in patients infected with HIV-1. We employed the standard recommended dose of 800 IU/day 24 versus a high-dose regimen of 25,000 IU/week cholecalciferol (vitamin D3) in this study. In the high-dose study cohort, skin-homing marker CCR10 expression on Treg was increased while CCR4 expression and the percentages of CCR4-expressing cells were decreased. On the other hand, there were no notable differences in the expression of either the skin- or gut-homing markers in the low-dose treatment group. The lack of effect seen in the low-dose cohort could be related to the lack of treatment efficacy with a standard dose of 800 IU cholecalciferol per day in this specific patient population. We saw that only 22% (2/9) of the patients in this cohort responded with a significant drop in PTH level and achieve serum 25(OH)D3 level of >75 nmol/liter (30 ng/ml). This effect is similar to that achieved by UV exposure during summer in the study region (Fig. 1C).
High-dose pulsed therapy seems effective in correcting vitamin D3 deficiency in 86% (6/7) of HIV-infected patients. The median serum 25(OH)D3 was elevated by 3-fold, from 33.0 nmol/liter to 138.0 nmol/liter after 2 months. There is limited knowledge on the immunological influence of vitamin D3 supplementation in HIV-infected patients or the effective dose in this specific group of patients. In a cohort of 56 children (age 6 to 16) randomized to receiving a bimonthly supplementation of 100,000 IU cholecalciferol (with daily doses of calcium) or placebo, serum 25(OH)D3 concentrations were higher in the treated group but no differences in CD4 count or viral load over a 12-month period were observed. 13 In another study carried out in 40 patients with multiple sclerosis (MS), the efficacy of low-dose cholecalciferol (800 IU or less) and high-dose ergocalciferol or vitamin D2 (50,000 IU for 7–10 days and subsequently once or twice weekly) was being compared. 25 Parallel to our observation, the low-dose cholecalciferol treatment was unable to reverse the deficiency in MS patients while switching the patients to a high dosage of vitamin D2 resulted in a significant increase in their serum 25(OH)D3 level. Of note, these patient cohorts share two things in common with ours; both have the same geographic location (northern hemisphere) and both have an underlying immunopathology of a different nature. The fact that we did not detect any significant differences in chemokine receptors expression by Treg in the low-dose cohort implies that it would be worthwhile using a higher dose to study the immunomodulatory effects of vitamin D3 in patients with an underlying immune dysfunction. In fact, it has been suggested that a higher 25(OH)D3 level (>100 nmol/liter or 40 ng/ml) may be required to sustain the immune function. 26
Homing receptors are selectively expressed and regulated in different T cells subset. 21,27 Skin-homing T cells express CLA and by their binding to vascular E-selectin initiates the T cell rolling interactions on vascular endothelium, the first step in the migration process. Migration to the skin also involves CCR4 for the transition of blood to the dermis and CCR10 from the dermis to the epidermis, and interactions of integrin α4β1 with VCAM and of LFA-1 with ICAM-1. In human T cells, 1,25(OH)2D3 has been reported to down-regulate CLA expression but not CCR4, CCR6, and CCR7. 28 In vitro, it has been shown that addition of 1,25(OH)2D3 after cells were activated resulted in an induction of CCR10 and inhibition of CLA expression. 29
In the present study, high-dose cholecalciferol supplementation in HIV-infected individuals differentially augments the expression of skin-homing markers, CCR4 and CCR10, on Treg. In vivo vitamin D3 up-regulates CCR10 expression and depresses CCR4 expression as well as the frequency of CCR4-expressing Treg. Parallel to our in vivo observation, CCR10 expression on Treg was elevated in vitro while the effect on CCR4 was not significant. The eventual result could still be a preferential homing of Treg to the skin as influenced by 1,25(OH)2D3. We should consider that even though both CCR10 and CCR4 are implicated in lymphocyte trafficking to the skin, there may be a certain degree of overlap in the two pathways with either alone being sufficient for skin homing. 30 It has been shown that T cell accumulation within the skin was not impaired in CCR4-deficient mice. 31,32 Lehtimaki reported that Treg are able to infiltrate the skin of CCR4−/− mice as evident by an increased number of Treg and enhanced Foxp3 mRNA in an oxazolone-induced inflamed skin model. 31 Hence even though we observed a depressed CCR4 expression alongside an elevated CCR10 expression on Treg in HIV-infected subjects supplemented with a high weekly dose of cholecalciferol, it is tempting to speculate that the net effect of vitamin D3 may be a preferential homing of Treg to the skin.
The integrin α4β7 and CCR9 are expressed by T cells that migrate to the gut. As previously described, retinoic acid and vitamin D3, which are selectively produced in the gut and skin, respectively, differentially modulate the expression of gut- and skin-homing markers. 33 Retinoic acid has been associated with the up-regulation of CCR9 and integrin α4β7 expression while reducing the expression of CCR10 on T cells. On the other hand, 1,25(OH)2D3 does not affect the gut-homing markers but increased CCR10 expression. 34 Here, we did not observe any in vivo or in vitro effect by 1,25(OH)2D3 on the expression of CCR9 and integrin α4β7. Of note, it has been demonstrated that the HIV-1 envelope protein gp120 binds to α4β7. 35 As a result, CD4+CCR5hiα4β7 hi T cells are highly susceptible to infection by HIV-1 and gp120 engagement of α4β7 is thought to facilitate infection at the point of transmission.
In light of the importance of vitamin D3 in regulating the immune function, interventions to improve vitamin D3 status in HIV-infected patients continue to warrant attention. Our findings serve as a caution to clinicians that treatment with a standard dose of 800 IU per day cholecalciferol may be inadequate to reverse the deficiency in HIV-infected individuals or to serve any immunomodulatory effects. The effect from a high weekly pulsed-dose therapy with vitamin D3 in HIV-infected individuals is suggestive of the induction of skin-homing potential on Treg. From this pilot study, it would seem justified to employ a higher dose of vitamin D3 in order to determine if and how vitamin D3 can exert immunological effects in HIV-seropositive individuals. On the other hand, these observations warrant future validation in a larger population.
Footnotes
Acknowledgments
The authors thank Ronald van Beek for technical assistance and Rob Woestenenk for his assistance in high-purity flow cytometric cell sorting. This work was partially supported by the Nutricia Research Foundation, The Netherlands. M.G.N. was supported by a Vici grant of the Netherlands Organization for Scientific Research.
Author Disclosure Statement
No competing financial interests exist.