
Abstract
Sexual transmission of HIV is the major cause of spread of HIV in Africa and the Third World and is an unmet medical need. Recently, microbicides have attracted attention because they allow females to protect themselves and their offspring. We are exploiting one of the four retroviral enzymes, the ribonuclease H, RNase H, as a novel approach for a microbicide. It is the only enzyme of HIV not yet targeted by antiretroviral therapy. The enzyme is linked to the reverse transcriptase (RT) and hydrolyzes the RNA moiety of RNA–DNA hybrids. The RNase H is located inside virus particles and normally functions during viral replication inside cells. Here we show that activating the RNase H prematurely inside the virus particles destroys the viral genome and abrogates viral infectivity. The antiviral compound consists of a synthetic oligodeoxynucleotide (ODN), which creates an artificial RNA–DNA hybrid substrate for the RNase H inside the particle. The compound was analyzed in mouse models including humanized SCID mice and the vagina of mice. Infection was reduced up to 1000-fold or could be completely prevented. The compound is suitable as microbicide or to prevent mother-to-child transmission.
RNase H during Viral Replication
R
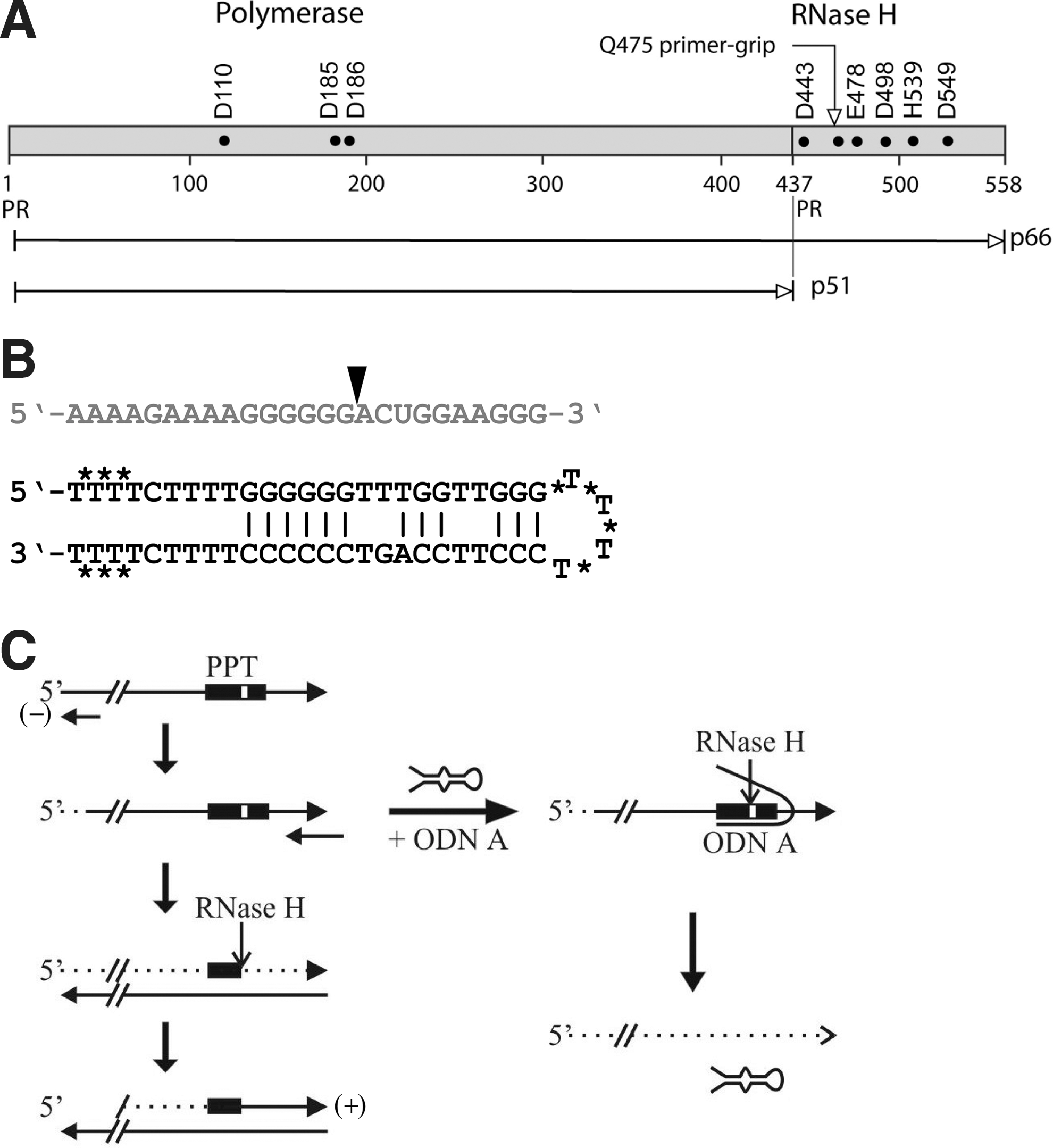
The RNase H carries out several functions. Its main role during viral replication is the hydrolysis of the viral RNA after it has been reverse-transcribed to cDNA by the RNA-dependent DNA polymerase activity of the RT. The RNase H then helps to supply primers for the RT to initiate second-strand DNA synthesis. All DNA polymerases require RNA primers for initiation of DNA synthesis. The transition from RNA to double-stranded DNA requires synthesis of two DNA strands and two primers. The first one is the cellular tRNAlys3 and the second one is the polypurine tract (PPT) RNA. 1,5 The tRNA primer is derived from the host cell and binds to the primer binding site (PBS) with the help of a melting protein, the viral nucleocapsid protein NCp7. 6 –8 The second primer is part of the viral RNA and resists hydrolysis by the RNase H during first-strand DNA synthesis. 9 The PPT RNA forms nonclassical structures with the DNA transcript, which protects it from cleavage by the RNase H. 10 It is the primer for initiation of the second-strand DNA synthesis by the DNA-dependent DNA polymerase of the RT. The two primers are then removed by the RNase H after they are no longer needed for DNA synthesis. Thus the RNase H not only generates primers, but also removes them.
The RNase H cleavage occurs at five different steps during replication. For removal of the two primers, cuts occur at RNA–DNA junctions opposite to the DNA strand. 1,5,11 Cleavage at the PPT is highly specific, because it will lead to the dinucleotides required for integration by the viral integrase into the host DNA genome. 1,5 The RNase H itself cleaves nonspecifically, but sequence specificity is dictated by the polymerase domain and the distance between the two active centers between the RT and the RNase H. This distance is 18 nucleotides and, consequently, the length of the primer at the PBS is 18 nucleotides, as is the cleavage site at the PPT. 9 The RT and the RNase H act in concert, whereby the RNase H functions as a processive exonuclease and a sporadic endonuclease. 12 The RT induces cleavage in particular at certain G-rich sequences, which leads to stuttering or stalling of the RT and cuts by the RNase H. The cleavage products of the RNase H are distinct from those of many other nucleases in that they consist of ribonucleotides with 3′-hydroxyl and 5′-phosphate ends. The linker region between these catalytic domains is an inactive, degenerated RNase H domain, which may have arisen by a duplication event. 13 The RT has been targeted by nucleoside RT inhibitors (NRTIs) or the nonnucleoside RT inhibitors (NNRTIs). 5 Despite its importance in the retroviral life cycle, the RNase H is the only enzyme of HIV not yet targeted by antiretroviral therapy.
RNase H—a Universal Enzyme
The RT-RNase H structures are slightly different in the retroviruses of different species, so they cannot be used easily as models for HIV drug development. For instance, the murine viral RT consists of a single polyprotein of 80 kDa, and the avian
RNases H are not restricted to retroviruses, but are universally required for supply of primers for cellular DNA synthesis in almost all forms of life. They belong to the five most conserved and oldest protein structures in the biosphere. 14 RNase H-like enzymes consists of five beta-sheets and several (three to five) alpha-helices, comprising in total about 150 amino acid residues. 15 Structural analyses have demonstrated that many enzymes have RNase H-like structures. The relatedness was not detected by comparison of the primary sequences but structural similarities, requiring crystallization. A hallmark of the RNase H-related enzymes is four conserved amino acid residues, DEDD (D443, E478, D498, D549), which are separated by other amino acid residues. These amino acid residues coordinate divalent cations, Mg2+ or Mn2+, required for enzyme activity. The family has been designated as the RNase H family or integrase family. 16
The RNase H domains are often associated with other domains that supply important additional functions, such as hybrid binding domains, supplied by the RT to the RNase H, DNA binding domains for the integrase, etc. 16
Of special interest is the similarity between retroviral RT-RNase H and the enzymes involved in antiviral defense, the Argonaute proteins, which mediate RNA interference by small interfering RNAs (siRNAs). Argonaute proteins consist of two domains, PAZ and PIWI, with PIWI having an RNase H-like structure and activity. 17 The PAZ and PIWI domains are required for siRNA-mediated silencing, which can even be induced to perform functions similar to the viral RT-RNase H depending on the choice of the divalent ions. 18 Thus composition and function of viruses and the antiviral defense machinery are related, which suggests a long history of coevolution. 18
There are two main types of cellular RNases H, RNases H1 and H2. The retroviral RNase H resembles an RNase H1, which is present in all forms of life. Eukaryotic RNase H1 is necessary during embryogenesis and mitochondrial DNA replication and is mainly localized in the cytoplasm. 19,20 Eukaryotic RNase H2 is more complex, consisting of three subunits, RNase H2A, H2B, and H2C, with only one of them, H2B, exhibiting enzymatic activity. The three subunits cooperate. 19,21 The RNase H2 is essential in eukaryotes, and mutated forms cause a human autosomal recessive genetic disorder, the Aicardi–Goutières Syndrome (AGS). 22 This enzyme can remove single ribonucleotides within a double-stranded DNA, 22 suggesting a role in the removal of misincorporated ribonucleotides as a kind of proofreading. It also removes RNA in RNA–DNA hybrids, so-called R loops, newly transcribed RNA that can loop back into the double-stranded DNA, for maintenance of genome integrity. 19,22 The RT and several RNases H have been identified in bacteria, 23 in yeast, and in Caenorabditis elegans, and RTs were also found in Archaea. The bacterial RNases H have been designated HI and HII. 23
RNase H Is Required for Viral Infectivity
The highly conserved structures of RNases H allowed predictions for inactivating mutations of conserved amino acid residues required for enzymatic activity. Based on the sequence of the Escherichia coli RNase H, a conserved histidine residue, H539, was selected for site-directed mutagenesis to N or D (Fig. 1A). 24 C-terminal deletions of 19 or 21 nucleotides also abolished infectivity. 25 Mutation of Q475 to E reduced endonuclease activity and processivity. 12 This residue is part of a critical primer-grip region important for hybrid binding and cleavage specificity. 15 This demonstrates the importance of the retroviral RNase H for replication and validates it as a target for drug design.
Oligodeoxynucleotides as Antiretroviral Compound
We selected the extended PPT as the target for an oligodeoxynucleotide (ODN). The PPT is highly conserved in HIV-1, with 69% of 2094 isolates being 100% identical in the sequence, including the related SIV. 26 The ODN is in total 54 nucleotides, consisting of an antisense arm and a second arm forming a hairpin-loop structure (Fig. 1B). Both strands are connected by a linker consisting of four deoxythymidines. The linker and the three terminal nucleotides at each end are linked by phosphorothioate bonds to increase stability against nucleases. The antisense arm is fully complementary to the target retroviral RNA. The partially complementary, hairpin-loop structure of the ODN is essential because a fully complementary ODN would not open at a target RNA. Formation of an RNA–DNA hybrid is thermodynamically favored over a DNA double strand. 27
We originally designed the ODN as a triple helix-forming ODN. The triple helix involves Hoogsteen bonds in addition to Watson–Crick hydrogen bonds, which are preferably formed at polypurine stretches, involving TAT or GCG trinucleotides, in which G is protonated (G+). They do not easily form inside a cell because the nuclei usually do not provide the acidic milieu needed for protonation of G. We cannot fully exclude that the ODN also forms triple helices with the viral target RNA, which would enhance inhibition of HIV, albeit by a different mechanism. However, the cleavage of the RNA indicated that the antisense effect dominated and the second arm of the ODN protected it during uptake. 27 Two ODN strands can form G cages, where four Gs are coordinated by a central potassium ion. The G clusters allow stacking of several layers of G cages. These form a stable structure, which may protect against nuclease attacks and enable uptake through the viral or cellular membrane without detergent. 27
High sequence specificity of the ODN was proven with two HIV isolates with slightly different PPTs. Exchange of two nucleotides out of the 25 nucleotide antisense strand significantly reduced the antiviral effect of the ODN. 26
Microbicides against HIV
Microbicides are an unmet clinical need in fighting the disease, and could render women capable of protecting themselves against HIV infection. 28 Topical microbicides, mainly administered as vaginal gels, have been intensely studied. The development statuses of current and potential topical microbicides against HIV are listed in Table 1. Initially investigated compounds based on surfactants aimed to physically destroy the retroviral membranes. One promising compound, nonoxynol-9 (N9), was active in vitro and also showed promising results in the feline model for HIV, but was ineffective as a gel-based topical microbicide in a Phase III clinical trial, and even increased susceptibility to HIV infection. 29 –31 Two Phase III clinical trials with another surfactant-based topical microbicide, C31G, were discontinued due to lower-than-expected incidence rates, and showed no efficacy as well. 32,33
NRTIs, nucleoside reverse transcriptase inhibitors; NNRTIs, nonnucleoside reverse transcriptase inhibitors.
Polyanion-based microbicides interact with the positively charged region of the HIV surface protein gp120, thus blocking retroviral infectivity. While also showing antiviral activity in vitro, such compounds, namely PRO2000, Carraguard, and cellulose sulfate, were ineffective in Phase III clinical trials, and may also even increase the risk of HIV infection. 34 –36 One further, polyanion-based microbicide, SPL7013, is currently under investigation in a Phase I/II clinical trial. 37
Another strategy pursued with topical microbicides aims to maintain the low pH milieu of the vagina to prevent retroviral infection. One compound, BufferGel, was ineffective in a recent Phase II/IIb clinical trial. 38 Another pH-buffering compound, Acidform, has shown some effectiveness in a Phase I/II clinical trial, however, without statistic significance. 39
The failures of this first generation of unspecific microbicides lead to the investigation of HIV-specific compounds as microbicides. In particular, the NRTI tenofovir formulated as a 1% topical gel has been investigated as anti-HIV-1 microbicide and showed significant reduction of HIV-1 acquisition by an estimated 39% in the CAPRISA 004 double-blinded randomized trial for prevention of HIV-1 acquisition in South African women. 40 This study is presently repeated. Other HIV-specific compounds, NNRTIs, fusion inhibitors, or CCR5 coreceptor antagonists are presently in preclinical or early clinical development (for details, see Table 1). 41 –47
The long list of failures, with the promising exception of the tenofovir microbicide, calls for new and innovative microbicidal approaches against HIV. None of the compounds mentioned above targets the RNase H. RNase H inhibitors have recently been investigated in vitro, but have not yet been tested in animal studies. 48 They may suffer from cross-reactivity against cellular RNases H, making them toxic to cells. We are exploiting the RNase H in a different way, by activating instead of inhibiting the enzyme inside virus particles in a sequence-specific manner. We found that ODNs are capable of destroying infectivity of HIV particles before infection occurs, making this an interesting candidate for a microbicide. This is supported in vitro as well as by animal studies.
We used the ODN to treat cell-free virus particles and demonstrated that ODNs were able to enter the virus particles and induce cleavage of the viral RNA in artificially created hybrids. This prevents viral replication (Fig. 1C). We modeled the ODN into a schematic RT-RNase H model and the crystal structure (Fig. 2A and B). Treatment of viral particles with ODN resulted in a significant reduction of viral RNA as well as infectivity. 27 Furthermore, we analyzed 19 primary patient's isolates from the Hospital in Zurich as well as primary isolates including multidrug-resistant ones from Rwanda, Africa, a total of 10 isolates, which were provided by the NIH. 49 Cell-free virions were treated with ODN in the plasma and subsequently the HIV RNA levels were determined by qRT-PCR, as well as infectivity on primary human peripheral blood cells. Infectivity of viruses was reduced 1000-fold in 30% of the cases within 4 h of treatment with ODN. It may be possible to reduce mother-to-child transmission by ODN infusion to mothers during labor prior to delivery to reduce infection of the newborn during birth. This would also be possible in the case of resistance of the mother to other drugs.

Mouse Models
To perform small animal studies we used mice infected with the oncogenic murine retrovirus, the spleen focus forming virus (SFFV), which causes tumors of the spleen. Its PPT is not identical to the one from HIV, and the ODN was designed specificly for its PPT. 50 We applied the ODN by intravenous as well as intraperitoneal routes. Treatment with increased concentrations of ODN allowed a dose-dependent reduction of the virus load in the blood, delayed disease progression, and death from 50 to 70 days. SFFV associated with red blood cells in the blood was also reduced, suggesting that transport of the retrovirus inside the body can be inhibited. We also demonstrated that the virus load is kept down by repeated treatments with ODNs every 12 to 24 h. Finally we showed that viral infection can be completely prevented if the animals are treated early and with high concentrations of the ODN. Prevention required ODN therapy at the same time as virus infection. 50 We then tested HIV in a mouse model endowed with a human immune system. We used mice with a severe combined immune deficiency (SCID) and human peripheral blood lymphocytes, huPBL-SCID. These mice allow HIV infection and replication. When treated with HIV and ODN, five out of six mice did not become infected. 51 It appeared that a single early treatment was sufficient to successfully prevent infection. If at this time point the virus escaped, repeated treatments were ineffective in preventing viral replication. We also noticed a similar effect with the SFFV-treated mice. 52 Thus we conclude that an early and hard treatment is most effective and requires a low dose of only about 5 to 25 μM ODN. Based on these results we suggest that the most likely success achieved by this approach will be a topical application as a topical microbicide.
Vaginal Microbicide Studies
Vaginal studies in mice were performed with a recombinant lentivirus, which is produced from a vector system in a packaging cell line. 52 We determined that 5 μM ODN inactivated 95% of the virus (106 IU) in vitro within 4 h. Mice were pretreated with hormone Depo-Provera to synchronize the estrus cycle. After 1 week the mice were treated with an intravaginal inoculum of recombinant lentivirus FUGW (104 IU) with or without ODN (25 μM) in caboxymethyl cellulose (CMC) gel. The regimen of treatment with ODN was 30 min before or 30 min after virus inoculums, as a prophylactic or therapeutic design, respectively. The virus load was determined 4 h later in lavage fluid and the virus was quantified by quantitative PCR. A 30-fold to 100-fold viral reduction was determined. We were also able to demonstrate uptake of a GFP-expressing FUGW lentiviral vector in primary human vaginal VK2 and cervical Ect1 cells. Uptake was demonstrated by immunofluorescence. In these cells, the ODN affects the virus similarly. We compared ODNs modified by phosphorothioate modifications and 2′-O-methyl modifications at the ends and within the linker region. The methylated ODNs had a slightly superior inhibitory effect. 52 No toxicity was observed in the vagina and cervix in human cells, as well as in the mice. Furthermore, the results demonstrated that the ODN did not affect only cell-free virus particles. The ODN could also reduce the amounts of DNA provirus copies inside the human cells. Thus, the ODN exhibited efficacy in vaginal or cervical cells, which may be important for a microbicide, since virus particles that may escape the ODN treatment inside the vagina may be inactivated inside the cell later on.
The stability and efficacy of the ODNs may be further improved by other modifications. Recently, locked nucleic acid (LNA) modifications showed very high stability up to several months. 53 We also showed that the ODNs did not induce resistance when compared to other inhibitors, and are effective against multidrug-resistant virus isolates. Recently, clinical trials have indicated unexpected side effects of compounds in the vagina, which lead to an effect opposite than intended—to an increased instead of a reduced uptake of virus. Also the effect of semen has to be evaluated. This mouse model for vaginal transmission of retroviruses suffers from the fact that the recombinant virus cannot infect the vaginal cells in the mouse, because it is not infectious. Therefore at present we have no proof as to whether infection and systemic spread of HIV are prevented by the ODN. This could be answered only by monkey studies, which are currently underway.
Drug Resistance and Toxicity
We furthermore demonstrated that the ODN did not give rise to drug resistance at a rate similar to a known inhibitor of HIV in cell culture. 51 The presence of ODN in the culture medium during serial transfer inactivated the virus quickly—immediately after leaving the cell. We managed to keep the culture for 14 passages with no occurrence of resistance. ODN inhibited growth of HIV in vitro in a single passage up to 100- or 1000-fold. A more than 105-fold reduction can be achieved, and the virus remains drug-sensitive even after prolonged treatment. In contrast, treatment with Foscarnet induced multidrug resistance within a few passages. 51 We attribute the strong inhibitory effect by the ODN to destruction of viral infectivity during medium transfer by each passage. Cleavage of viral RNA is different from binding of an inhibitor. Thus the ODNs may exhibit properties that predestine them as microbicides. To our knowledge no other compound exists that specifically drives HIV particles into a suicide in a sequence-specific manner. Vaginal rings may allow the application of ODNs with controlled release over longer periods and may replace application of gels. The synthesis of ODNs has become cheaper with increased demands. ODNs have previously not indicated toxicity in humans in many antisense trials, which makes their application promising. 53
Since we are specifically inhibiting the cell-free virus particles with ODNs, application could be preexposure prophylaxis (PrEP) in the form of a microbicide against sexual transmission of HIV in the Third World. ODNs targeted to virions are the only compounds known to specifically destroy virus before infection. This may be of benefit for prevention of mother-to-child transmission and therapeutic treatment also. There are 6.6 million new infections per year, with about 400,000 newly infected children. The number of women who may want to protect themselves could likely be hundreds of millions, which may allow the compound to be made cheaply by mass production. Microbicides are the most urgent unmet medical need to fight the spread of HIV.
Footnotes
Acknowledgments
We are grateful to Drs. Konstantin Fackeldey and Marcus Weber for help in preparing the model of the crystal structure of the RT-RNase H and stimulating discussions. We are indebted to Prof. Dr. Peter Deuflhard for his support of this work.
Author Disclosure Statement
No competing financial interests exist.