
Abstract
The anti-HIV microbicide, tenofovir (TFV) gel, has been shown to decrease HIV-1 acquisition by 39% and reduce herpes simplex virus 2 (HSV-2) transmission by 51%. We evaluated the effect of a 1% TFV gel on genital HSV-2 infection in a mouse vaginal challenge model. In vitro plaque assays and luminex multiplex bead analysis were used, respectively, to measure postinfection vaginal viral shedding (day 1) and cytokine secretion (day 2). To further investigate the anti-HSV-2 properties, we evaluated the direct antiviral effect of TFV and the oral prodrug tenofovir disoproxil fumerate (TDF) in cell culture. Compared to placebo-treated mice, TFV-treated mice had significantly lower clinical scores, developed later genital lesions, and showed reduced vaginal viral shedding. Furthermore, the levels of IFN-γ, IL-2, TNF-α, and other cytokines were altered in the vaginal fluid following topical tenofovir treatment and subsequent HSV-2 challenge. Finally, we found that both TFV and TDF inhibited HSV-2 infection in vitro; TDF showed a 50-fold greater potency than TFV. In conclusion, we confirmed that the microbicide TFV had direct anti-HSV-2 effects in a murine vaginal challenge model. Therefore, this model would be suitable for evaluating present and future microbicide candidates. Furthermore, the present study warrants further investigation of TDF in microbicides.
Introduction
H
Effective prophylaxis is the most efficient way to halt both HIV-1 and HSV-2 infections. Microbicides are pharmacological agents that block or inactivate sexually transmitted pathogens; they are applied topically in either the vagina or rectum. They may be used as pre-exposure prophylaxis (PrEP) applied prior to sexual intercourse and/or post-exposure prophylaxis (PEP) applied after sexual intercourse. 7 An important property of microbicides may be to limit local inflammation, by reducing either the levels of secreted proinflammatory cytokines or the number of immunological target cells in the genital mucosa. 8 Tenofovir (TFV)-containing microbicide trials have shown divergent results. In the first successful anti-HIV microbicide trial (CAPRISA 004), a gel containing 1% TFV was proven effective against HIV infection, with an overall 39% reduction in the rate of HIV acquisition compared to a placebo gel, 9 whereas the use of TFV gel was discontinued due to lack of efficacy in the recent VOICE trial. 10 However, the CAPRISA 004 study provided the additional finding that TFV gel reduced the risk of HSV-2 acquisition by 51% 11 independently of HIV seroconversion status. This suggests that the TFV gel may prevent HIV-1 and HSV-2 transmission through separate, independent mechanisms. 11 A recent study showed that TFV had a direct anti-HSV-2 effect in cell cultures through inhibition of the HSV-2 DNA polymerase. 12 In addition to direct antiviral effect, TFV and the oral prodrug of TFV, tenofovir disoproxyl fumarate (TDF), may affect innate and adaptive immune responses. In vitro, TFV induces cytokine production in murine peritoneal macrophages and enhances interleukin (IL)-12 production after a lipopolysaccharide challenge in macaque peripheral blood mononuclear cells (PBMCs). 13 –15 We have previously shown that in human primary PBMCs TDF reduced IL-10 secretion, enhanced IL-12 secretion, and decreased proinflammatory cytokine secretion in response to a bacterial or viral infection or to the stimulation of innate, pathogen-sensing, toll-like receptors. 16
In the present study, we hypothesized that an anti-HSV-2 effect of vaginally applied TFV could be verified in a vaginal challenge mouse model. The study design was similar to that used in the CAPRISA 004 study with application of TFV once before and once after virus challenge. The endpoints included an evaluation of the clinical health of the mice, vaginal viral titers, and a characterization of the vaginal cytokine profile in HSV-2-infected mice treated with either TFV gel or placebo gel. Based on previous findings in vitro, we also hypothesized that TFV and the oral prodrug TDF could inhibit HSV-2 infection by a mechanism directly inhibiting virus replication. Therefore, we conducted an in vitro study to evaluate the direct antiviral effects and cytotoxicity of TFV and the oral prodrug TDF.
Materials and Methods
Mice, viruses, and TFV gel
The mice used in this study were 7-week-old, C57BL/6, females (Taconic M&B, Ry, Denmark). All animal experiments described were reviewed and approved by The Animal Experiments Inspectorate, Copenhagen, Denmark (approval number 2009/561/1641). The 1% TFV gel comprised (R)-9-[2-(phosphonomethoxy)propyl]adenine (R-PMPA) (NIH AIDS Research and Reference Reagent Program, Bethesda, MD) diluted in purified water and a hydroxyethylcellulose (HEC) gel solution (universal HEC placebo gel, NIH AIDS Research and Reference Reagent Program, Bethesda, MD), with a final TFV concentration of 10 mg/ml. For the control group, we used the universal HEC placebo gel diluted in purified water. To synchronize the mice susceptibly to HSV-2 infection, mice were pretreated with 200 μl of subcutaneously administered medroxyprogesterone diluted in phosphate-buffered saline (PBS) at a concentration of 10 mg/ml (Depo-Provera; Pfizer, Ballerup, Denmark) 5 days before the HSV-2 infection. The intravaginal infection was achieved with a lethal dose of strain 333 HSV-2 (6.67×104 pfu/mouse), delivered in 20 μl of Iscoves medium (Lonza, Basel, Switzerland). HSV-2 strains were amplified in vero cells and quantified by viral titration as previously described. 17
Mouse infection studies
Mice (n=56) were randomized by cages into two groups; one received 20 μl 1% TFV vaginal gel and the other received HEC placebo gel. The gel was applied 10–12 h before and 10–12 h after the HSV-2 infection. Mice were anesthetized with isoflurane [2-chloro-2-(difluoromethoxy)-1,1,1-trifluoro-ethane] for the gel applications and for the infection. To allow absorption of the gel or virus, mice remained anesthetized for 5–10 min after each application. In a separate study, we evaluated the vaginal viral titer and cytokine profile in vaginal lavages from 20 mice, 10 in each group; the protocol was repeated three times (total n=60) in exactly the same way. The vaginal lavages were performed on days 1 and 2 post-infection (pi) (the study flow chart is shown in Fig. 1). For the lavages, mice were anesthetized, and 30 μl Iscoves Medium was pipetted in and out of the vagina, 10–12 times, with three repetitions (total lavage volume, 90 μl). Lavages collected from each mouse were immediately placed on dry ice and stored at −80°C. Before visible signs of infection had developed (day 1), the group allocation (TFV vs. placebo) was blinded to the investigator by a third part to ensure an objective clinical evaluation. The follow-up included daily monitoring of weight and disease score, based on the following scale: 0: healthy; 1: genital erythema; 2: moderate genital infection; 3: purulent genital lesions and/or in generally poor condition; 4: hind limb paralysis (leading to euthanasia) (Table 1).
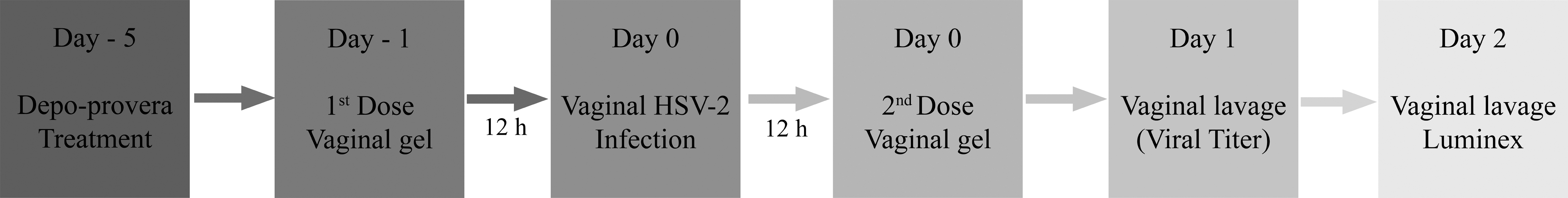
Flowchart of study design. On day −5, mice (n=56) were pretreated with subcutaneous medroxyprogesterone. On day −1, the mice received the first dose of either 1% tenofovir (TFV) vaginal gel or the corresponding placebo gel [vehicle hydroxyethylcellulose (HEC) gel]. After 10–12 h the mice were vaginally infected with herpes simplex virus 2 (HSV-2); 10-12 h later, they received the second dose of the gel. Mice were monitored daily for changes in weight and disease scores from days 0 to 6. Another group of mice (n=60) underwent vaginal lavages on days 1 and 2 to determine the viral titer (with in vitro plaque assays) and the cytokine profile (with luminex assays).
Viral plaque assays
A standard vero cell plaque assay was used to evaluate the direct antiviral activity of TDF (9-[(R)-2 [[bis[[(isopropoxycarbonyl)oxy]methoxy]phosphinyl]methoxy]propyl]adenine fumarate) and TFV (both from NIH AIDS Reagent and Reference Program, Bethesda, MD). Cells were grown in Dulbecco's modified essential medium (Lonza, Basel, Switzerland) supplemented with 5% heat-inactivated fetal bovine serum (FBS) and 1% penicillin-streptomycin (both from Invitrogen, Glostrup, Denmark). After 24 h of culture, TFV was added to the cells at concentrations of 10 μM (3 μg/ml) to 1000 μM (287 μg/ml) and TDF was added at concentrations of 10 μM (6 μg/ml) to 100 μM (64 μg/ml). The control cultures received PBS as a vehicle. The HSV-2 wild-type strain MS (HSV-2MS) was added 30 min after TFV pretreatment.
A vero plaque assay similar to that described above was used to assess the vaginal viral titers from the lavages collected on day 1 pi. The vaginal lavages were diluted and added to cells after 24 h of culture. For both vero assays, 0.8% human immunoglobulin (Beriglobin; CSL Behring, Pennsylvania) was added after incubation for 1 h. The cells were then maintained for 48 h at 37°C in 5% CO2. Subsequently, cells were stained with 0.03% methylene blue in distilled water and viral plaques were counted.
Cytokine profiles from vaginal lavages
The lavages from day 2 were analyzed with a luminex multiplex bead assay (Invitrogen, Glostup, Denmark) for cytokines interferon (IFN)-γ, interleukin (IL)-1α, IL-2, IL-6, CXC chemokine ligand 10 (CXCL10) (IP-10), CC chemokine ligand 2 (CCL2) (MCP-1), CC chemokine ligand 9 (CXCL9) (MIG), CC chemokine ligand 3 (CCL3) (MIP-1α), tumor necrosis factor (TNF)-α, and fibroblast growth factor (FGF) as described by the manufacturer.
Cell toxicity studies
To evaluate TFV and TDF cell toxicity, vero cells were seeded in 96-well plates and maintained overnight before applying TFV and TDF. After a 24-h incubation, the cells were stained with the [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] (MTT) substrate. Briefly, cells were incubated for 3 h in media that contained 0.5 μg/ml MTT (Sigma-Aldrich, Brondby, Denmark). Subsequently, cells were lysed with a 1:1 vol. 96% EtOH and DMSO; cell survival was quantified by reading the absorbance at 540 nm.
Statistical analyzes
Data from the viral titer and luminex assays were log transformed. Comparisons of disease score, vaginal viral titers, and cytokine concentrations were performed with a Mann–Whitney test. Disease-free survival was analyzed with a log-rank test. In vitro data were analyzed with a four-parameter fit. All statistical analyses were performed with PRISM Graph Pad version 5.0.
Results
Topically applied TFV reduced HSV-2 infections in C57BL/6 mice and increased disease-free survival
To evaluate the clinical effect of TFV microbicide on vaginal HSV-2 infections in mice (n=56), either TFV or placebo gel was applied in the vagina at two times: 10–12 h before and 10–12 h after a vaginal inoculation of HSV-2 at a lethal dose. The flow chart of the study is presented in Fig. 1. Subsequently, mice were monitored daily for changes in weight and disease scores. The mean disease score (MDS) was calculated for each group (Table 1). We found that TFV-treated mice were healthier and fewer mice exhibited lesions than placebo-treated mice; the result was a 50% lower MDS on day 6 in the TFV group (p=0.0287) (Fig. 2A) [MDS TFV: 0.5 (95% CI: 0.1421–0.8579), MDS placebo: 1.0 (95% CI: 0.5193–1.409)]. Disease-free survival (DFS) was assessed as the time from infection to the appearance of the first symptom/lesion. The TFV group exhibited a significantly longer DFS compared to the placebo group (p=0.0045) (Fig. 2B). We also observed a lower mean weight loss in the TFV group compared to the placebo group (data not shown). Taken together, these data showed a protective or disease-modifying effect of the TFV microbicide gel against a high titer HSV-2 challenge in the vaginal tract.

TFV gel reduced vaginal HSV-2 titer in infected mice
To evaluate whether the reduced infection and mortality in the TFV group were due to a reduction in vaginal viral titer we collected vaginal lavage on day 1 pi (n=60) and assessed the vaginal viral burden in a vero cell plaque assay. We found that TFV gel significantly reduced the vaginal viral burden in HSV-2-infected mice (Fig. 3). On day 1, the mean viral titer was 8.4×104 pfu/ml (SEM±2.2×104 pfu/ml) in the TFV group, and it reached 1.6×105 pfu/ml (SEM±3.6×104 pfu/ml) in the placebo group (p=0.0246). In conclusion, TFV gel significantly reduced the viral load in the vagina.
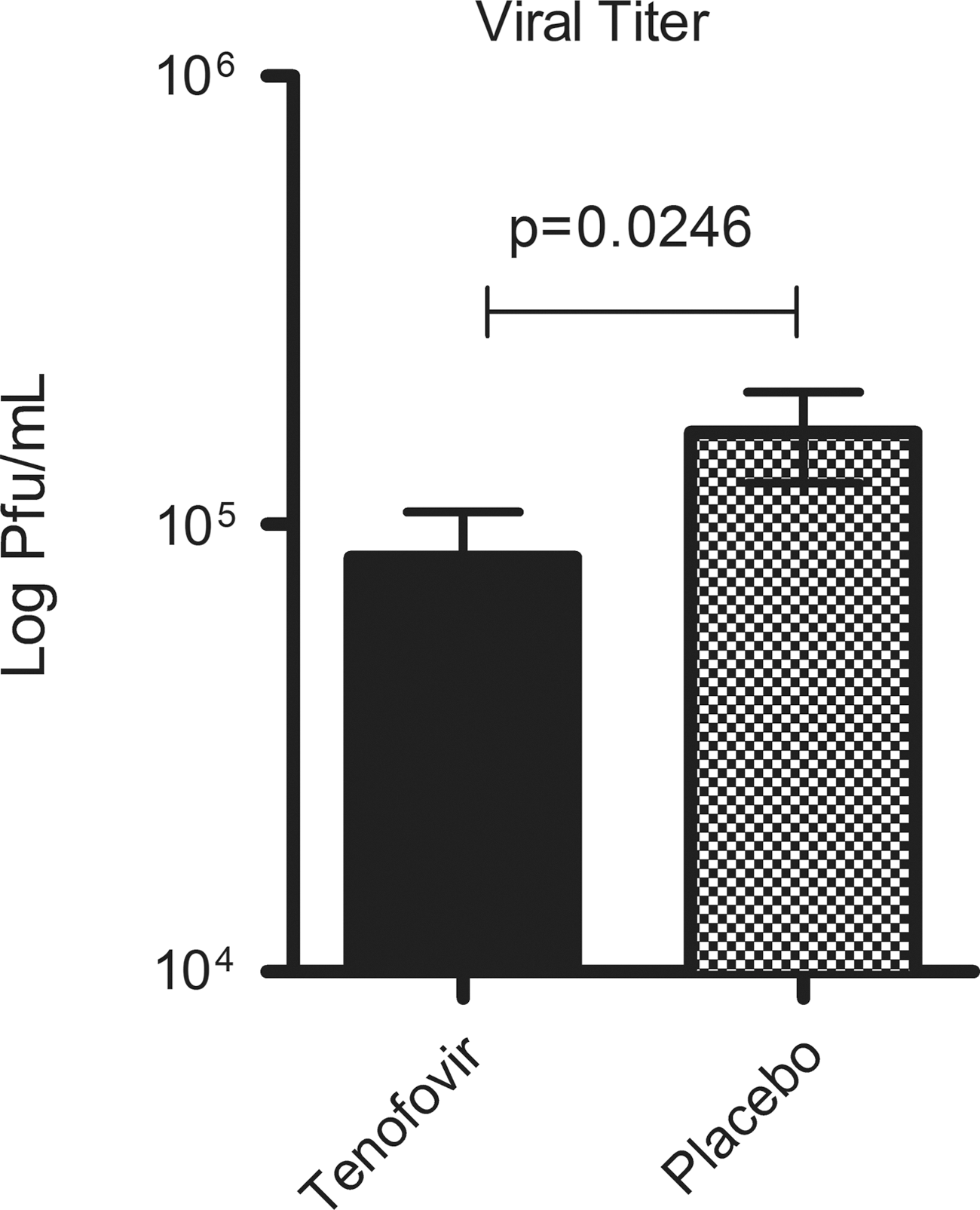
TFV gel significantly reduced the vaginal viral burden in HSV-2-infected mice. Viral titers were measured from vaginal lavages performed on day 1 pi (n=60). Data from in vitro plaque assays were pooled from three separate trials that used the same procedure and showed the same trend. Data were log-transformed, and represent the mean±SEM; comparisons were performed with the Mann–Whitney test.
TFV changed the cytokine profile in the murine vagina during HSV-2 infection
Because the cytokine milieu in the vagina can negatively or positively affect inflammation and antiviral responses, we evaluated the impact of TFV gel during HSV-2 infection. Vaginal lavages collected on day 2 were analyzed for levels of several inflammatory and immunoregulatory cytokines, including chemokines, IFN-γ and FGF, in a luminex multiplex bead assay. The TFV group showed significantly higher levels of IL-1α compared to the control group (p=0.0213). Similar trends were observed for IFN-γ and the chemokine CCL3, but these were not statistically significant (p=0.132 and p=0.386, respectively) (Fig. 4A). In contrast, the placebo group showed significantly higher levels of the sentinel proinflammatory mediator TNF-α compared to the TFV group (p=0.0265). Similarly, the placebo group showed significantly higher levels of IL-2 compared to the TFV group (p=0.0232). The same trend, though not significant, was found for IL-6 (p=0.553), CCL2 (p=0.259), and CXCL10 (p=0.355). The placebo group also showed a significantly higher level of the proangiogenic cytokine FGF compared to the TFV group (p=0.0167) (Fig. 4B). In conclusion, the presence of TFV selectively affected cytokine levels in the vagina during HSV-2 infection.
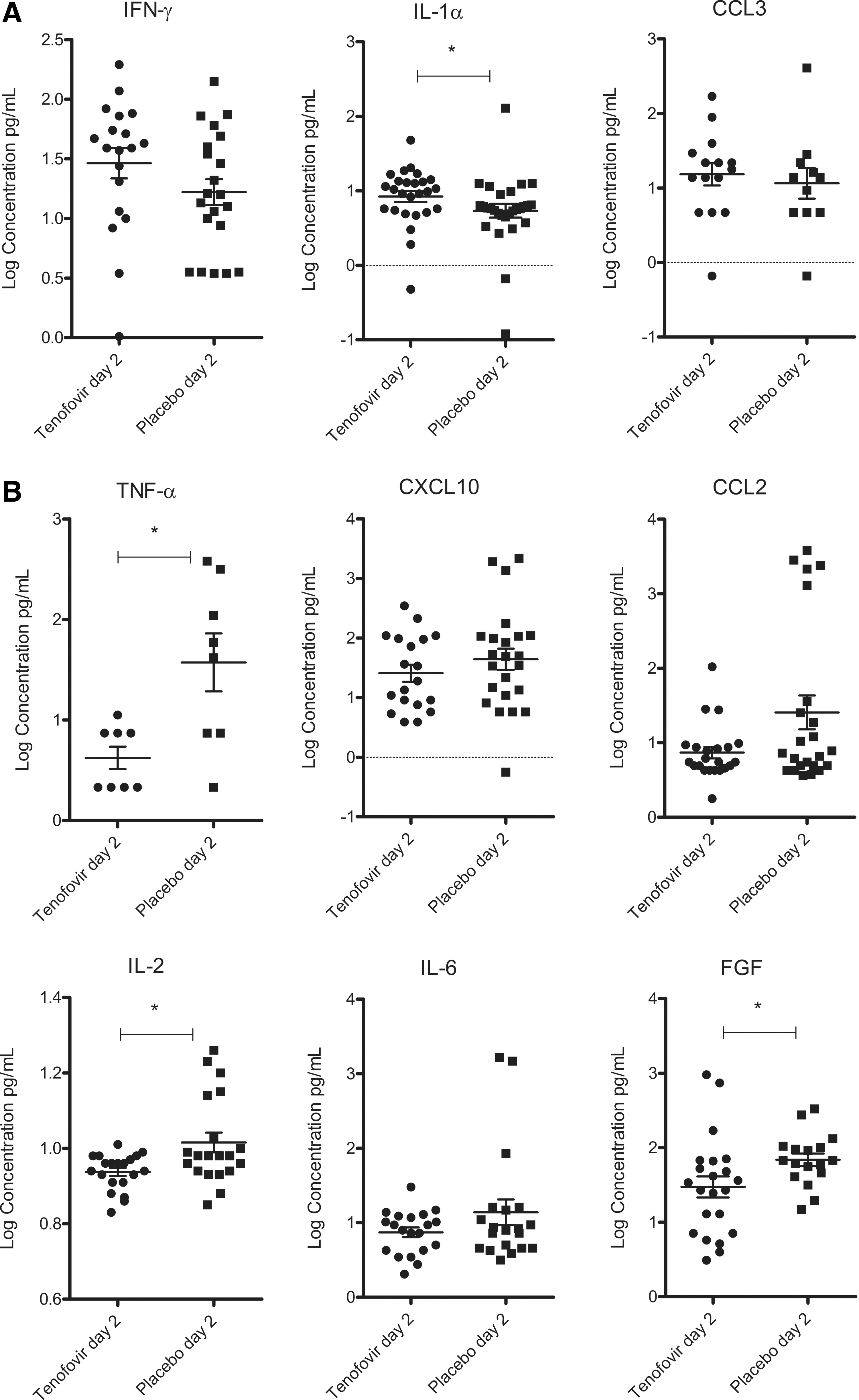
Cytokine levels were measured in vaginal lavages collected on day 2 pi. Assessments were performed with the luminex multiplex bead assay (n=60).
TFV and TDF inhibited HSV-2 infection in vitro and showed variable cell toxicity
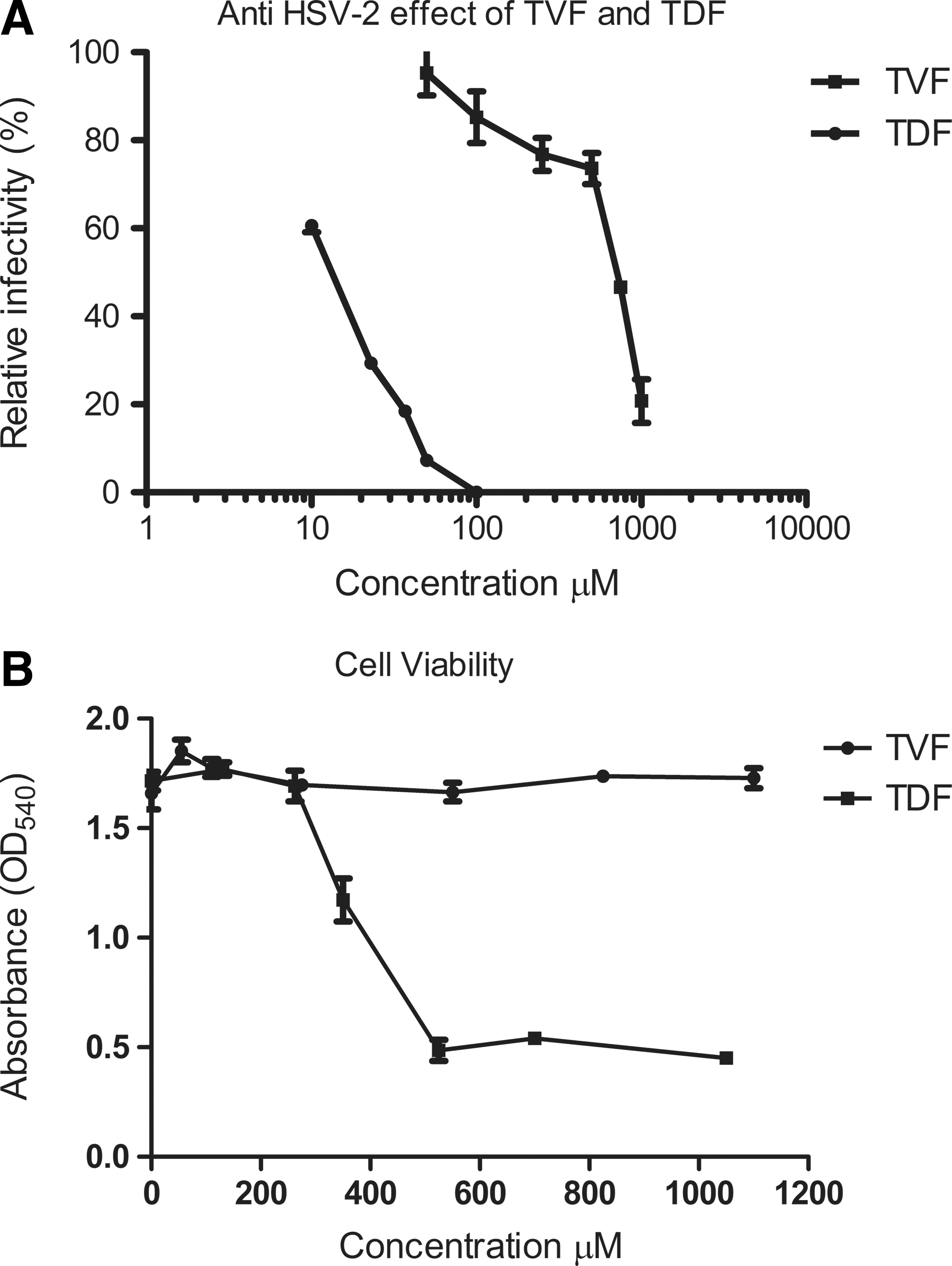
The observation that TFV-treated mice showed reduced viral loads and improved clinical scores prompted us to investigate the direct antiviral effect of TFV and TDF with vero plaque assays. TFV was found to inhibit infections of HSV-2MS in vero cell cultures at concentrations of 100 to 1000 μM (29 to 287 μg/ml) with an EC50 of 636 μM (182 μg/ml) (concentration that causes 50% of maximal effect) (Fig. 5A). On the other hand, TDF caused a reduction in HSV-2MS infection at much lower concentrations of 10 to 100 μM (6 μg/ml to 64 μg/ml), with an EC50 of 13.27 μM (8.4 μg/ml). Cell toxicity studies showed no effect of TFV on cell viability, but TDF reduced cell viability at concentrations >262 μM (Fig. 5B). In conclusion, TFV and TDF had direct anti-HSV-2 effects at concentrations that were not toxic to cells.
Discussion
In the present study, we demonstrated that the microbicide TFV, applied in the vagina prior to and after a HSV-2 challenge, decreased morbidity and mortality in C57BL/6 mice and reduced the vaginal viral titer. Furthermore, TFV treatment altered the cytokine profile in the murine vagina compared to placebo. Finally, we found that TFV and TDF had direct anti-HSV-2 effects with different potencies in the vero plaque assay.
Previous mice studies had not determined the exact role of topically applied TFV against HSV-2 infection; moreover, no studies included a vaginal lavage to determine viral titer to validate clinical findings. 12,18 One study showed a protective effect of TFV, but they used athymic nude mice in a lumbosacral scarification model. 12 Athymic nude mice are devoid of an effective immune response, and a vaginal challenge model was not used. Therefore, that model may not be optimal for predicting either direct antiviral effects or potential secondary immunomodulatory effects in the vagina. Another study was conducted in BALB/c mice; they found that a vaginal TFV gel did not protect against a vaginal HSV-2 infection when the gel was applied for 7 consecutive days before the HSV-2 infection, but not after infection. 18 This negative result was not explained in the BALB/c mouse study; however, we might speculate that the timing of TFV application and possible administration of TFV after infection are essential for a positive clinical effect. Furthermore, strain specificity and the number of mice studied may have played a role in the results. It might be that TFV does not inhibit HSV-2 in vivo, but merely inhibits HSV-2 in the vero cell plaque assays. However, the recovered amount of TFV in the vaginal lavages does not exceed the minimum concentration for inhibition of HSV-2 in vitro (Fig. 5A). Therefore, we conclude that TFV inhibits HSV-2 in vivo (Fig. 3). To our knowledge, this study was the first to show that TFV gel had a protective or disease-modifying effect following a high titer, vaginal HSV-2 challenge in mice and to examine the effects of TFV gel on an HSV-2 infection directly at the port of entry.
A previous study demonstrated that when TFV in solution was applied to humanized BLT mice 4 h before and 4 h after infection, it inhibited an HIV-1 infection by 88% compared to placebo. 19 Taken together, these findings elaborated on previous studies in mice 20 confirming that the vaginal mouse model is a useful platform for testing microbicides in simple, preclinical efficacy evaluations.
In this study, the lethal viral dose of 6.7×104 pfu/mouse ensured a maximal infection rate that would facilitate reliable clinical evaluations for group comparisons. We focused on the early effects of TFV on HSV-2 disease (day 6), because that period entirely reflected the outcome of the PrEP/PEP treatment and viral exposure. Later effects can be confounded by lateral viral transmission among cagemates.
Previous studies suggested that TFV enhanced the production of TNF-α, IL-10, IL-1β, CCL5, and CCL3 in murine peritoneal macrophage cultures. 13,14 Moreover, TFV was reported to increase IL-12 production from macaque PBMCs in response to lipopolysaccharide stimulation. 15 We previously found in human primary PBMCs that TDF reduced IL-10 secretion, enhanced IL-12 secretion, and further reduced the levels of proinflammatory cytokines IL-8 and CCL3 in response to pathogen infection and stimulation of innate pathogen-sensing receptors. 16 In the present study, we found that topical TFV treatment altered the vaginal cytokine-profile at 48 h after HSV-2 infection compared to controls (Fig. 4). IFN-γ levels were elevated with TFV treatment compared to placebo. Elevated IFN-γ levels early during infection may be beneficial, because IFN-γ is important for increasing the effectiveness of T cell-dependent HSV-clearance. 21 In contrast, we found that the placebo group had significantly higher levels of the proinflammatory cytokine TNF-α compared to the TFV group. Reduced TNF-α levels during early vaginal infection could also be beneficial to the host, because anti-TNF-α antibody treatment after an HSV-2 vaginal infection has been found to prolong survival in mice, possibly due to reduced TNF-α-mediated leukocyte infiltration in the CNS. 22 It remains to be determined whether the observed reduction in vaginal TNF-α levels after TFV treatment was a direct effect of the treatment or a result of the reduced viral replication. In either case, the reduced TNF-α level may have contributed to prolonged survival in the TFV group.
An important property of anti-HIV microbicides may be the ability to reduce local inflammation, 8 which limits the availability of target cells. The ability of TFV gel to reduce the levels of proinflammatory mediators, such as TNF-α, CCL2, and CXCL10, may therefore be generally beneficial during very early infections. Overall, the TFV group did not exhibit high levels of inflammatory cytokines; this was consistent with previous findings in BALB/c mice, which showed reduced or unaltered levels of inflammatory cytokines in the vaginal tract after TFV treatment. 18 Conversely, we found that the concentration of the proangiogenic growth factor FGF was significantly higher in the placebo group than in the TFV group (p=0.0167); this may have been due to greater viral shedding in this group and, hence, more severe HSV-induced genital lesions. In addition, IL-2 levels were significantly lower in the TFV group (p=0.0232), which could be an important observation regarding the effect on HIV acquisition. IL-2 is a vital T cell proliferative cytokine; hence it may be important for the susceptibility to HIV infection. A limitation of our study was that although the levels of signaling molecules are known to change throughout the course of infection, we observed only a narrow time-interval of 48 h pi. Thus, our overall results were difficult to interpret and compare to previous findings measured at different time points. Also, the observed differences in measured cytokines may simply reflect the differences in HSV-2 infection in the two groups. Collectively, future studies should delineate these possible effects of TFV in vivo and quantify their contribution to enhanced viral clearance and reduced HSV-2 and HIV-1 diseases.
Finally, our in vitro studies demonstrated a direct anti-HSV-2 effect of TFV and the oral prodrug TDF. Our results were consistent with those from a recent study that also identified a direct anti-HSV-2 effect of TFV in vitro. 12 Our study also provides evidence for an antiviral effect of TDF against HSV-2 stain MS in Vero cells. Furthermore, our data are in line with a very recent report showing antiviral effect of TDF and TFV in human cervical cells. TDF has been proposed as an orally administered PrEP, but studies have shown that the concentration of TFV in cervicovaginal fluid reached only 68 ng/ml at the end of a 24-h oral dosing interval. 23,24 This is far below the demonstrated inhibitory concentration for HSV-2 strains in human cell cultures (EC50 of 123–193 μg/ml). 12,24 In comparison, the local concentration of TFV in cervicovaginal fluid upon topical administration at the end of a 24-h dosing interval is 100 μg/ml. 25 In the present study, we found an EC50 of 636 μM (182 μg/ml) for TFV and a substantially lower EC50 of 13.27 μM (8.4 μg/ml) for TDF.
Taken together, these data suggested that only topically administered TFV would show at least some effect against an HSV-2 infection. TDF is metabolized in plasma to TFV, which is phosphorylated intracellularly to the active metabolite tenofovir diphosphate (TDP). 26,27 Thus, the potential anti-HSV-2 effect of orally administered TDF would be unlikely to reach the genital tract. Consistent with this, no reduction in HSV-2 shedding was observed in patients with HIV that were using HAART that contained TDF. 28 Alternatively, we propose that the pharmacokinetic challenge may be more effectively overcome and the anti-HSV-2 effect of TFV might be optimized by adding TDF to the 1% TFV gel at a very low concentration (e.g., 50:1 of TFV:TDF).
However, formulation and administration of TDF microbicides may be important since TDF may be hydrolyzed in aqueous gel. 29 Therefore, alternative delivery methods may be better, such as TDF delivery via intravaginal rings. 23
Two observations indicated that the anti-HSV-2 effect of TDF in cell culture was probably not mediated by TFV after conversion from TDF. First, we used heat-inactivated fetal bovine serum; therefore, very limited amounts of active plasma esterases should have been present in the vero cell cultures. 30 Second, the effective antiviral dose of TDF was below that of TFV. Therefore, we concluded that both TFV and TDF possessed direct anti-HSV-2 effects. It remains to be determined whether the direct antiviral effects were through separate, independent mechanisms. Of note, we observed a toxic effect of TDF at concentrations >262 μM (Fig. 5B), but no toxic effects were observed with TFV, even at the highest concentration (1,100 μM). Further evaluation of these possible effects (including adverse events and toxic effects) requires additional studies, for example, clinical testing of TDF as PrEP/PEP in mice and human vaginal ex vivo explant tissues.
In conclusion, our study provided evidence that TFV inhibited high-titer HSV-2 infections in the female genital tract. Furthermore, our in vitro studies confirmed that the observed in vivo effect of TFV was, at least partly, due to a direct anti-HSV-2 effect. Moreover, we provided new evidence that TDF had a direct, very potent, antiviral effect. This result may provide a basis for the development of a new dual-acting microbicide against HIV and HSV-2. As the VOICE trial did not reproduce the efficacy seen in the CAPRISA 004 trail 9,10 the need for new antiviral drugs in microbicides is highly relevant. In light of the VOICE data our in vitro finding, that TDF is highly antiviral against HSV-2, could be of great importance. Further studies must delineate whether the TFV microbicide can also exert immunomodulatory properties in vivo, and whether the inclusion of TDF in future microbicide gels or by alternative delivery methods, such as intravaginal rings, may positively affect the clinical outcome.
Footnotes
Acknowledgments
This work was supported by grants from the Scandinavian Society of Antimicrobial Chemotherapy (SSAC), the Søren Segel & Johanne Wiibroe Segel's Research Fund, and Department of Clinical Medicine, Aarhus University, Denmark. Line Vibholm is supported by a scholarship and Line S. Reinert by a fellowship from the Faculty of Health Science, Aarhus University, Denmark.
Part of the data in this article was presented at the 28th Annual Meeting of the Scandinavian Society of Antimicrobial Chemotherapy (SSAC), Reykjavik, August 18 to 21, 2011 and the Keystone Symposia, Frontiers in HIV Pathogenesis, Therapy and Eradication, March 26 to 31, 2012, Whistler, British Columbia, Canada.
Author Disclosure Statement
No competing financial interests exist.