
Abstract
The high HIV-1 prevalence, up to 4.6% in Guinea-Bissau, West Africa, makes it a relevant location for testing of therapeutic vaccines. With the aim of performing a clinical study in Guinea-Bissau, after first testing the vaccine for safety in Denmark, Europe, we here describe the design of a universal epitope peptide-based T cell vaccine with relevance for any geographic locations. The two major obstacles when designing such a vaccine are the high diversities of the HIV-1 genome and of the human major histocompatibility complex (MHC) class I. We selected 15 CD8-restricted epitopes predicted from conserved regions of HIV-1 that were subdominant (i.e., infrequently targeted) within natural infections. Moreover, the epitopes were predicted to be restricted to at least one of the five common HLA supertypes (HLA-A01, A02, A03, B07, and B44). Here, we validated the resulting peptide-specific, HLA-restricted T cell specificities using peptide–MHC class I tetramer labeling of CD8+ T cells from HIV-1-infected individuals. The selected vaccine epitopes are infrequently targeted in HIV-1-infected individuals from both locations. Moreover, we HLA-typed HIV-1-infected individuals and demonstrated that the selected vaccine epitopes, when targeted, are restricted to the five most common HLA supertypes at both locations. Thus, the HLA supertype-directed approach achieved HLA coverage of 95% and 100% of the examined cohorts in Guinea-Bissau and Denmark, respectively. In conclusion, the selected vaccine epitopes match the host populations and HIV-1 strains of these two distant geographic regions, justifying clinical testing in both locations.
Introduction
T
Therapeutic vaccination aiming at activating cellular immunity with or without ART is a promising method for disease prevention. 3 The purpose of such immunotherapy is to focus a cellular immune response against conserved viral determinants potentially contributing a broadly based immune response preventing viral escape. The idea of using a polyepitope vaccine was first described by Hanke and McMichael. 4
Appropriate MHC class I-restricted CD8+ T lymphocyte responses are important for HIV-1 and simian immunodeficiency virus (SIV) disease control. 5 Immunodominant epitopes, which are primarily targeted during the infection, have been the first to be identified and the first to be used for T cell-based vaccine design. 6 However, the immunodominance of specific CD8+ T cells may hinder development of potentially more effective responses to subdominant epitopes. 7 –10 Thus, the vaccine approach is to redirect CD8+ T cell immunity during the infection to infrequently targeted (immunologic subdominant) epitopes, which independent of clade and geographic origin are conserved among HIV-1 strains. 10 –14 In a test-of-concept therapeutic vaccine trial in Denmark we demonstrated that it is possible to redirect immunity to such subdominant epitopes during an untreated chronic HIV-1 infection. In that study, we focused cellular immune responses on seven subdominant human leukocyte antigen (HLA)-A*02-restricted epitopes and HIV-1 subtype B infection in asymptomatic HIV-1-infected HLA-A*02+ Danish individuals. 15
To broaden this principle, we here describe the rational design of a peptide-based vaccine that matches the host populations and HIV-1 strains in any geographic location, here exemplified by Denmark, Europe, and Guinea-Bissau, West Africa. Also, for ethical reasons, the vaccine should be tested for safety in our own country, Denmark, before Guinea-Bissau and therefore the vaccine needs to match different populations. There are two major challenges when designing such a vaccine: the differences and the high variation of the HIV-1 genome as well as the differences in distribution of the human major histocompatibility complex (MHC) class I alleles.
In Denmark and Europe, subtype B is the dominating subtype of HIV-1, 16 whereas in Guinea-Bissau the HIV-1 epidemic is dominated by subtype A3, the circulating recombinant form CRF02_AG, and subtype C. 17,18 However, infrequently targeted and immune subdominant epitopes are often conserved among HIV-1 strains independent of clade and geographic origin and may therefore be good targets of a broad therapeutic vaccine. 10 –13
To overcome the high diversity of the human MHC class I alleles we chose to select CD8+ T cell epitopes that are restricted within the five most common HLA-supertypes. HLA class I molecules can be clustered into groups, designated as supertypes, representing sets of molecules that share largely overlapping peptide binding specificity. 19,20 The selected vaccine epitopes were originally predicted by artificial neural network computers 14,21 and the peptide-HLA binding predicted using the pan-specific NetMHCpan method. 22 Here we describe the rational selection and antigenicity of the selected vaccine epitopes in HIV-1-infected and HLA-typed individuals in Guinea-Bissau. Using peptide–MHC class I tetramer labeling of peripheral blood mononuclear cells (PBMCs) from HIV-1-infected individuals we also validate the predicted HLA restriction, and the hypothesis that the vaccine would match the populations of different geographic regions.
Materials and Methods
Patients
Guinea-Bissau
A consultative statement of approval (J.nr.:2006-7041-70) was obtained by the Developing-Country Committee of the Danish National Committee on Biomedical Research Ethics (J.Nr.2006-7041-70) and was subsequently approved by the Ministry of Health in Guinea-Bissau (Trial registration of 22-Jan-2007 and NCP No. 04/2008). Prior to the interview and blood sampling, the patients were informed about the study in Creole (verbally) and Portuguese (literally). Informed consent was obtained by signature or fingerprint, and the subject was allowed to have a bystander present. The patients were all asymptomatic HIV-1-positive adults (18–50 years old), with CD4+ T cell counts above 400 per μl blood, and HIV-2, HBV, HCV, and TB negative.
Denmark
The protocol was approved by the Capital Region's Ethical Committee on Biomedical Research (H-D-2008-063). Written informed consent was provided by all individuals. The patients were all asymptomatic HIV-1-positive adults, with CD4+ T cell counts above 400 per μl blood, and HIV-2, HBV, HCV, and TB negative.
United Kingdom
Ethics approval was given by the Oxford Research Ethics Committee (Thames Valley cohorts). Study subjects gave written informed consent for their participation. The patients were all healthy HIV-1-positive adults, with CD4+ T cell counts above 400 per ml blood, and HIV-2, HBV, HCV, and TB negative.
HLA genotyping
Guinea-Bissau
Genomic DNA was purified using the Qiagen Blood DNA mini kit (Qiagen, Hilden, Germany) according to the manufacturer's instructions. The DNA in elution buffer was stored at −80°C until analysis. HLA alleles A (exon 2, 3, and 4), B (exon 2, 3, and 4), and C (exon 2, 3, and 4) were sequenced with AlleleSEQR (Abbott Laboratories) according to the manufacturer's instructions. The sequencing products were precipitated with BigDye Xterminator solution and SAM solution (Applied Biosystems, Carlsbad, CA), vortexed for 30 min, and then centrifuged briefly. Electropherograms were generated by the ABI Prism 3100 Genetic Analyzer, and analyzed with Assign 3.5+ Software (Conexio Genomics, Australia). To achieve maximum resolution of the genotypes, high-resolution sequencing primers HARPS (HLA Allele Resolution Probe System, Abbott Laboratories, IL) were used.
Denmark
Genomic DNA was purified using the QIAamp DNA Mini Kit (Qiagen) and shipped to Genome Diagnostics BV (Netherlands) for HLA genotyping.
Intracellular cytokine staining
PBMCs were isolated from whole blood, either by Ficoll gradient centrifugation or using CPT tubes (BD Biosciences, San Jose, CA), according to the manufacturer's recommendations. PBMCs were rested overnight at 37°C, 5% CO2, in cell medium [RPMI-1640 (Sigma-Aldrich, St. Louis, MO), supplemented with 2 g/liter NaHCO3 (Gibco, Carlsbad, CA), 10% fetal bovine serum (Gibco), and 100 units/ml penicillin/streptomycin (Gibco)]. The following day 0.125–0.5×106 cells were stimulated with pools of peptides (Schafer-N, Lyngby, Denmark), with each peptide at a concentration of 5 μg/ml. This concentration of peptide during stimulation was used in order to detect low avidity responses. Peptides were grouped in pools according first to HLA restriction and second to protein category (structural/enzymatic Gag and Pol; structural Env; accessory/regulatory). Staphylococcus enterotoxin B (Sigma-Aldrich), 1 μg/ml, served as a positive control, and medium alone served as background control. Stimulations were done during 6 h, 37°C, 5% CO2, and in the presence of 12.5 μg/ml Brefeldin A (Sigma-Aldrich). Following stimulation, the cultures were treated with EDTA (2 mM) (Sigma-Aldrich) to separate any cell aggregations formed during stimulation. Cells were subsequently fixed and permeabilized using BD cytofix/cytoperm kit (BD Bioscience) according to the manufacturer's instructions. After permeabilization cells were stained with anti-CD3 PerCP (BD Biosciences), anti-CD8 PE (Dako), anti-interferon (IFN)-γ FITC (BD Biosciences), anti-interleukin (IL)-2 FITC (BD Biosciences), anti-tumor necrosis factor (TNF)-α FITC (BD Biosciences), and anti-macrophage inflammatory protein (MIP)1β FITC (R&D systems). Stained cells were fixed using 2% paraformaldehyde. Staining of CD3 and CD4 after permeabilization allowed detection of these markers even when down-regulated in response to stimulation.
Cells were acquired on the flow cytometer CyFlow SL (Partec, Münster, Germany). Data were analyzed with FlowJo (TreeStar, Ashland, OR). Between 50,000 and 250,000 events in the lymphocyte gate were collected per sample. The level of cytokine expression was defined within CD3+CD8+ lymphocytes. The samples were considered positive if the percentages of cytokine-stained cells were more than two times background stimulation (the highest out of four samples per assay) and with a distinct population of cytokine-positive cells.
Peptide/MHC class I tetramer staining
The relevant PE-labeled tetramers were produced as previously described. 23 PBMCs were thawed and rested 1 h at 37°C, 5% CO2, before staining with PE-conjugated tetramer for 20 min, then washed and stained with the following extracellular antibodies: anti-CD3 PerCP (BD Biosciences), anti-CD8 APC-H7 (BD Biosciences), CD4-Qdot605 (BD Biosciences), and Live/Dead marker Violet (Invitrogen) for another 20 min. Cells were washed, fixed using Stabilizing Fixative (BD Biosciences), and samples were acquired on a BD LSR II flow cytometer. Data were analyzed using FlowJo. Cells were gated on singlets, lymphocytes, live cells, CD3, and then evaluated for CD8+ cells binding the peptide/MHC tetramer. In each patient a staining using a tetramer loaded with peptide to which this patient did not respond was included as negative control. A distinct population within the CD8+ population, not seen in the CD8− population or in the negative control, was considered as a positive response.
Peptide-MHC binding
The peptide-MHC-I affinity measurements were done using a homogeneous AlphaScreen-based assay. 24 In brief, recombinant, biotinylated MHC-I heavy chains were diluted to a concentration of 2 nM in a mixture of 30 nM β2M and a dose titration of the peptide of interest, and allowed to fold for 48 h at 18°C. The pMHC-I complexes were detected using streptavidin donor beads and a conformation dependent anti-HLA-I antibody, W6/32, conjugated to acceptor beads. The beads were added to a final concentration of 5 μg/ml and incubated overnight at 18°C. Detection was done on an EnVision multilabel reader (Perkin Elmer, Boston, MA).
Results
Epitope selection
In addition to the already well-characterized seven HLA-A2-rescticted HIV-1 epitopes, 12,14,15 we selected eight additional epitopes within a larger set of epitopes described by Perez et al. 21 The criteria for selection were (1) two epitopes from each of the proposed supertypes A01, A03, B07, and B44, (2) infrequently targeted within natural infection (<25% of tested HIV-1-infected individuals), and (3) conserved with within different HIV-1 subtypes. The selected additional non-A2 epitopes were from Gag, Pol, or Nef proteins. The 15 CD8+ T cell epitopes are listed in Table 1.
Immunogenicity of the vaccine epitopes
The 15 selected vaccine epitopes (Table 1) have been shown to be infrequently targeted in HIV-1-infected individuals living in Denmark and Sweden. 12,21 Here, we examine the preexisting immunity to these selected vaccine epitopes in a cohort of 31 untreated asymptomatic HIV-1-infected individuals in Guinea-Bissau (Table 2), using intracellular cytokine staining and flow cytometry. We stained for IFN-γ, TNF-α, IL-2, and MIP1β (Fig. 1). Due to a limited amount of blood, all the cytokines were measured together using the same fluorochrome in order to detect a response rather than determine individual cytokine responses. For the same reasons, in the in vitro stimulations the vaccine peptides were grouped in pools according first to predicted HLA-A and -B allele supertype restriction and second to target virus protein category (Table 2). Also, we aimed to determine if the in vitro responses to epitopes match the predicted HLA restriction.

Intracellular cytokine staining for interferon (IFN)-γ, tumor necrosis factor (TNF)-α, macrophage inflammatory protein (MIP)-1β, and interleukin (IL)-2. Lymphocytes were identified through a side scatter (SSC) versus forward scatter (FSC) plot followed by identification of CD3+CD8+. Hereafter cytokine expression was identified. Representative plots of an SSC versus cytokine-FITC are shown for unstimulated negative control
nd, not determined; UC, unclassified.
We were able confirm the antigenicity and subdominant characteristic of the vaccine epitopes during HIV-1 infection. Thus the A01/B07 pool (peptide Nef73 and Pol934), the A02 pool (peptide Vif101, Vpu66, and Vif23), the B07 pool (peptide Gag148 and Pol311), and the B44 pool (peptide Pol592 and Nef107) induced cytokine response in CD8+ T cells from one to four individuals. The Gag/Pol A02 pool (peptide Gag150, Gag433, and Pol606), the Vif/Vpu A02 pool (peptide Env67), and the A03 pool (peptide Pol313 and Pol973) did not induce a measurable cytokine response in these individuals. In most cases the HLA types of the responding patients matched the supertype prediction of the peptide reagent (Table 2). The only exception was patient 1100 where a cytokine response was seen against the Vif/Vpu A02 pool (peptides Vif101, Vpu66, and Vif23), although there is no A02 allele expressed by this patient.
Vaccine coverage obtained by the HLA supertype approach in Guinea-Bissau and Denmark
The literature of HLA types in Guinea-Bissau is limited. 25 –27 We therefore examined HLA types in a cohort of 39 untreated asymptomatic HIV-1-infected individuals in the capital Bissau eligible for therapeutic HIV-1 vaccination (Table 3). The HLA supertype designation is provided in Table 2. The selected vaccine epitopes are predicted to be restricted within the five most common supertypes in Guinea-Bissau. The population coverage of the HLA supertypes A01, A02, A03, B07, and B44 was 33%, 31%, 69%, 51%, and 26%, respectively. Thus, the HLA supertype approach gains HLA coverage of 95% of this Guinea-Bissau cohort, i.e., 37 out of the 39 individuals had at least one HLA allele within the supertypes included in the vaccine, without selecting individuals based on their HLA type. Moreover, 23 out of these 39 individuals had at least one allele where HLA restriction/binding was confirmed by peptide–MHC class I tetramer labeling of CD8+ T cells from HIV-1-infected individuals and/or peptide-MHC binding affinity (see below). For comparison, we also determined the supertypes present in a similar cohort of 20 untreated asymptomatic HIV-1-infected individuals in Denmark (Table 4). Here the vaccine epitopes are restricted within supertypes present in 20 out of 20 individuals (Table 4), again without selecting individuals based on their HLA type. In Denmark the population coverage of the HLA supertypes A01, A02, A03, B07, and B44 was 50%, 45%, 45%, 75%, and 35%, respectively. Thus, by using these epitopes recognized within these five most common HLA supertypes, this vaccine should in theory be able to induce CD8+ T cell responses equally frequently in these two different populations.
na, not available.
nd, not determined.
Alternative HLA genotypes are given for individuals in whom allele ambiguities could not be resolved.
UC, unclassified.
HLA restriction of vaccine epitopes
The HLA-A*02:01 restriction of the seven A02 epitopes has been described previously. 14 Here, we verify the predicted restriction of the non-A02 epitopes 21 using peptide–MHC class I tetramer labeling of CD8+ T cells from HIV-1-infected individuals in the Danish cohort and Thames Valley cohorts (United Kingdom). Individuals were selected based on their pervious detected cytokine response toward the desired epitope (data not shown). The peptides Pol934, Pol313, Gag148, Pol311, Pol592, and Nef107 were all as expected restricted to at least one allele within the predicted supertype. In the case of Pol313 we could show HLA restriction to three different alleles, A0301, A1101, and A6801, all within the A03 supertype. Moreover, in our analysis of Pol973 we were able to show restriction to HLA-A*30:01 within the A01A03 supertype, i.e., an allele with the characteristics overlapping both the A01 and A03 supertypes. One example per HLA epitope restriction and one negative control are shown in Fig. 2.
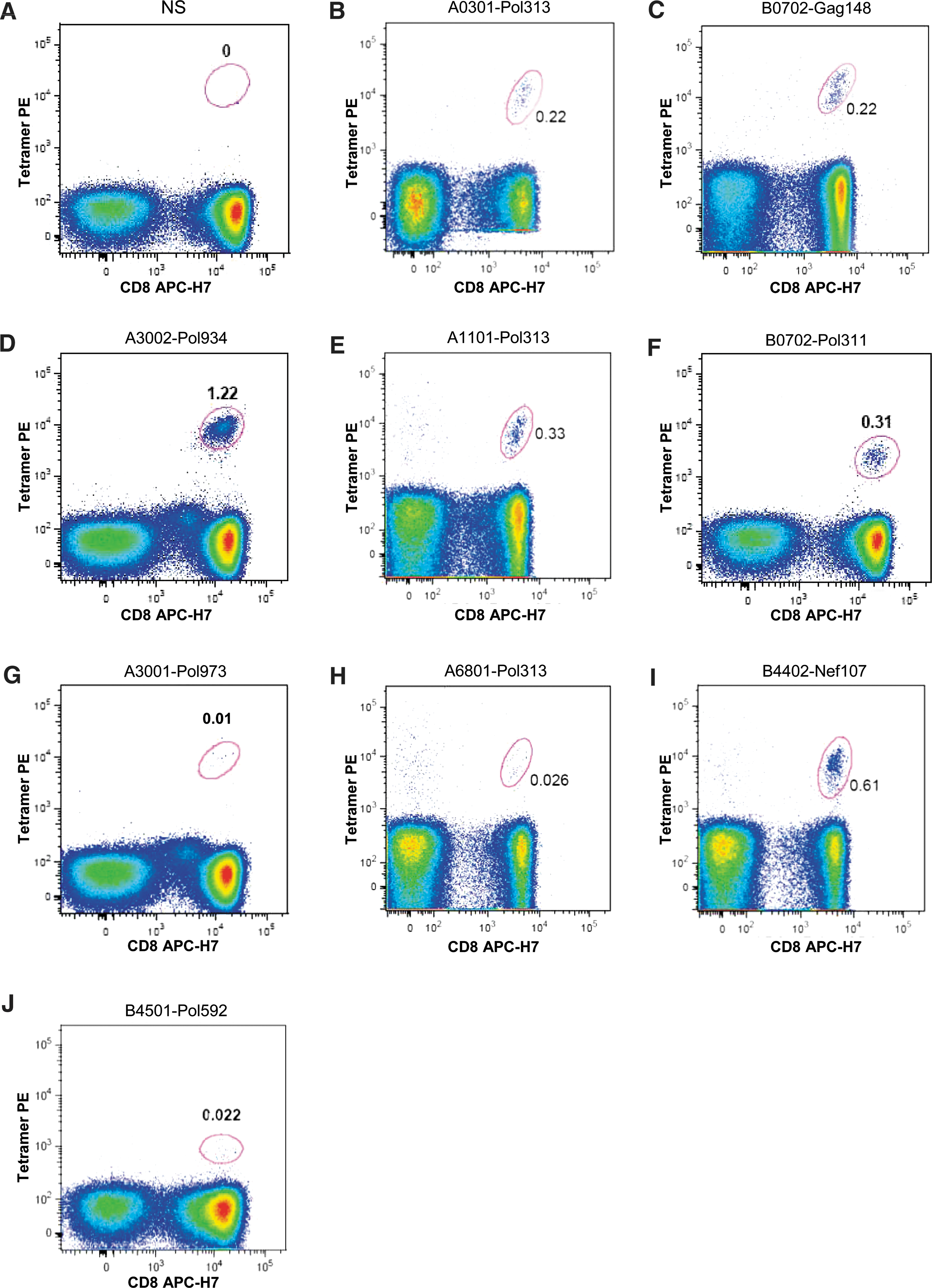
Epitope restriction using peptide–MHC class I tetramer labeling of CD8+ T cells from HIV-1-infected individuals. Singlets were identified using a forward scatter area (FSC-A) versus height (FCS-H) plot. Subsequently, lymphocytes were identified through a side scatter area (SSC-A) versus FSC-A plot, followed by exclusion of dead cells and identification of CD3+ events. Finally tetramer+CD8+ events were identified as shown. Negative staining
The HLA restrictions shown by peptide–MHC class I tetramer labeling were also confirmed by measured peptide–MHC binding affinity. Moreover, binding to more than one HLA allele is shown for six out of the eight epitopes peptides, with Pol934 binding to as many as seven different alleles, indicating promiscuous allele restrictions. Good and intermediate binding (IC50<500 nM) is shown in Table 5. Noteworthy, Pol934 shows binding to alleles within both the A01 and A03 supertypes.
In the case of the A01-Nef73 peptide we were unable to display binding to selected alleles within the predicted A01 supertype (data not shown). This encouraged us to predict HLA binding of this 9-mer (QVLPRPMTY) using the HLA restrictor server. 28 This predicted that the 8-mer VLPRPMTY within the 9-mer would bind to HLA-B3501 (B07 supertype), as in fact previously described. 29 The B07 supertype is indeed present in four out of seven of the patients responding to the 9-mer Nef73 in a previous study, 21 whereas only one out of the seven displayed the A01 supertype allele. Moreover, three out of the four responding patients with the B07 supertype actually express the HLA-B*35:01 allele. 21 It is therefore likely that in these cases it is the 8-mer (VY8) within Nef73 that is responsible for the response but through the HLA-B*35:01 allele (B07 supertype). However, in our cohorts of HIV-1+ individuals we did not find any individuals with the HLA-B*35:01 allele responding against Nef-73, and we were therefore unable to confirm this hypothesis using the peptide–MHC class I tetramer labeling technique.
Altogether, at least one restriction for seven of the eight non-A02 epitopes could be confirmed (Fig. 2) and a B07 supertype restriction was suggested for Nef73 (VY8). Thus, we were able to demonstrate two epitope restrictions not previously reported in the Los Alamos HIV immunology databases, 30 namely Pol973 (KK9) and Nef107 (QY9) to A3001 and B4402, respectively.
Discussion
An effective broad therapeutic vaccine inducing relevant CD8+ T cell responses against HIV-1 with the potential to eliminate already infected cells would be a valuable addition to the antiretroviral therapy available today. We here describe the design of a universal epitope peptide-based T cell vaccine matching HLA types and HIV-1 strains in different geographic regions.
With the aim to perform a clinical study in Guinea-Bissau, after first testing the vaccine safety in Denmark, we here confirm the relevance for both locations. To be able to test the same vaccine for safety and immunogenicity in HIV-1-positive individuals in different regions is not only relevant because of ethical considerations, but is also scientifically challenging. In an attempt to overcome diversity we selected minimal subdominant CD8+ T cell epitopes, conserved within different clades of HIV-1, and predicted to be restricted within five common HLA supertypes. 14,21 There are several other approaches to deal with HIV-1 diversity. For example, mosaic proteins optimized via a computational method to maximize the coverage of potential T cell epitopes 31 and chimeric proteins based on consensus sequences of conserved regions. 32
We have earlier confirmed the presence and conservation of the selected subdominant epitopes in full-length sequences of HIV-1 strains from Denmark and West Africa, including Guinea-Bissau. 11,13 Two epitopes are clearly more frequent in one of the local geographic regions: Vpu66 in Denmark and Nef107 in West Africa. However, we know from previous studies that these epitopes are conserved within the majority of HIV-1 subtypes 14,21 and were included in the vaccine. Importantly, the median number of these 15 epitopes simultaneously present per full length HIV-1 genome is similar in West African and Danish isolates, 10 and 11.5, respectively. 11
It is noteworthy that epitope conservation does not always correspond to epitope presentation. The amino acid sequence in the region flanking an epitope is important for intracellular processing and presentation. 33 However, with this high frequency of epitopes per genome the vaccine should theoretically be able to simultaneously target multiple sites and thereby make viral escape difficult or with the consequence of lower viral fitness. We do not know the impact on fitness from these individual epitopes. However, several of the epitopes described here were included in a study by Matthews et al. 34 where the HLA-associated escape was evaluated in a cohort of 710 individuals infected with C-clade HIV-1. The B07-Pol311 (SM9) epitope restricted to HLA-B*07:02 and the 8-mer VLPRPMTY restricted to HLA-B*35:01 were associated with viral escape. The latter were described as reverting, (i.e., the wild type is favored when the HLA selection pressure is removed), suggesting that the mutation has a negative impact on infectivity and/or replicative capacity. Moreover, the epitopes A01-Pol934 (KY9), A03-Pol973 (KK9), and A03-Pol313 (AK9) restricted by HLA-A*30:02, HLA-A*30:01, and HLA-A*30:01, respectively, are also associated with HIV-1 escape in the same cohort (unpublished data).
The chosen epitopes were shown to be immunological subdominant when tested within cohorts of HIV-1-infected viremic individuals living in Denmark 12 and Sweden. 21 Here, we also confirm this characteristic for the majority of the epitopes in a cohort of asymptomatic and untreated HIV-1-infected individuals living in Guinea-Bissau. However, a response could not be detected against all vaccine epitopes in this cohort. This is explained by the subdominant characteristic of the epitopes, the limited number of patients tested, and their HLA-alleles. Although the natural immunity toward the vaccine epitopes was evaluated during untreated and viremic HIV-1 infection it is not unlikely that during suppressed viral load (i.e., during ART) it would be more conceivable to induce additional and functional T cell responses. During chronic untreated HIV-1 infection there is a loss of immune function 35,36 that may be partially reversible by ART 37,38 and thereby the response to immunization improved.
As our therapeutic HIV-1 vaccine candidate is designed to match specific HLA genotypes we determined the relevant HLA alleles of clinical cohorts fulfilling our criteria for the intended immune therapy: asymptomatic HIV-1-infected adults living in Guinea-Bissau and Denmark. HLA supertype approach gains HLA coverage of 95% and 100% of these cohorts in Guinea-Bissau and Denmark, respectively. The vaccine should theoretically be able to induce CD8+ T cell responses in these two distant populations. Nevertheless, this does not mean that there is a 100% match between all the vaccine epitopes and HLA types present the cohorts, but that the vaccine should theoretically be able to induce at least one vaccine-specific response. The HLA frequencies seen in HIV-1+ individuals in Guinea-Bissau were similar to what is described by Spinola et al. 25,26 in individuals of unknown HIV status. This indicates that the data obtained from our limited cohort of HIV-1+ individuals reflect the population of Guinea-Bissau. However, our results differ from what Yindom et al. 27 recently described, likely explained by the different cohorts studied. Yindom et al. 27 studied individuals in the rural village of Caio, a relatively isolated community of the Manjako tribe in the northern part of Guinea-Bissau. In our cohort only six out of 39 individuals claimed this ethnicity.
The HLA types of the patients in Guinea-Bissau and previous studies 12,21 suggest that the epitopes are correctly predicted within the proposed supertypes. Although HLA prediction tools are useful for in silico predictions there are limitations in such data, which do not always correspond to experimental data. Therefore we verified the predicted restriction of the non-A02 epitopes using peptide–MHC class I tetramer labeling of CD8+ T cells from HIV-1-infected individuals. In most cases we could confirm restriction to at least one allele within the predicted supertype. It is important to note that these confirmed restrictions do not exclude numerous other HLA restrictions within the supertype, or even within other supertypes. The peptide–MHC binding data actually suggest that Pol934 would be restricted within the A01 as well as the A03 supertype. Extensive HLA promiscuity has been described. 39 Moreover, the 9-mers may be processed and presented as shorter epitopes with other restrictions. This is exemplified by the Nef73 epitope (QY9) where it appears to be an 8-mer epitope (VY8) that binds to HLA-B3501 within the B07 supertype. In the case of the Nef73 peptide we were unable to confirm any restriction using our peptide–MHC class I tetramers.
We have described the rational selection of a set of broadly covering HIV-1 vaccine epitopes to be included in therapeutic vaccine trials in Denmark and Guinea-Bissau. The epitopes were derived from conserved regions of HIV-1, were subdominant within natural infection, and were predicted to be restricted to five common HLA supertypes. We confirm that the selected epitopes match the HLA types of the host populations in the two distant geographic regions, Northern Europe and West Africa. Confirmed HLA restrictions of vaccine epitopes are associated with viral escape. The vaccine has been shown to be safe and well tolerated in preclinical toxicity testing. 40 This provides optimism for successful proof of concept testing during clinical trials in both Denmark and Guinea-Bissau using these epitopes as a peptide-based therapeutic vaccine.
Footnotes
Acknowledgments
We would like to thank the following: Drs. Gitte Kronborg and Jan Gerstoft (University Hospitals of Hvidovre and Copenhagen) and all the volunteers for participating in this study; Peter Aaby, for support and guidance through the Bandim Health Project; Birgit Knudsen, Solvej Kolbjørn Jensen, Irene Schrøder Jensen, and Bente Østergaard, for technical assistance. This work was supported by grants from the Danish International Development Agency (DANIDA) and The Danish AIDS Foundation.
Author Disclosure Statement
No competing financial interests exist.