
Abstract
Whether or not HIV-1 continues to infect cells in individuals treated with effective antiretroviral therapy (ART) remains controversial. Here, we determined whether the redistribution of the HIV-1 proviral burden with respect to antigen specificity of CD4+ cells would provide evidence for ongoing infection cycles in vivo. HIV-1 preferentially infects antigen-stimulated CD4+ T cells. In the setting of prolonged effective ART, we postulated that if infection cycles were occurring, influenza-specific CD4+ T cells, activated by influenza vaccination, would preferentially accumulate proviral burden. Peripheral blood mononuclear cells (PBMCs) were collected from HIV-1-infected subjects who had been treated with effective ART for >5 years, before and after influenza vaccination. CD4+ T cells were sorted by antigen specificity and HIV-1 proviral burdens were determined. Levels of HIV-1 production upon in vitro antigenic stimulation were also measured. At baseline, influenza-specific CD4+ T cells carried higher HIV-1 proviral loads than HIV-1-p55-specific CD4+ T cells. Upon influenza vaccination we observed trends toward elevated levels of HIV-1 proviral DNA in influenza and HIV-1-p55-specific, but not tetanus toxoid or cytomegalovirus (CMV)-specific CD4+ T cells. Higher levels of HIV-1 virions were produced upon influenza stimulation in postvaccination as compared to baseline samples. While the trends toward increased proviral burdens in influenza-specific cells failed to reach statistical significance, our observation of disproportionately high levels of provirus in influenza-specific cells at baseline indicates that this may represent a real increase that is cumulative over multiple annual vaccinations. This has implications for the eradication of HIV-1 by adding to the evidence that the resting CD4+ T cell viral reservoir is continually replenished in ART-treated subjects.
Introduction
T
The existence of ongoing viral replication in ART-treated subjects does, however, remain an area of controversy with other studies showing no evidence for decay of residual plasma viremia (1–49 copies/ml) upon ART intensification. 15 –18 An argument has also been made against ongoing replication based on the observation of very low levels of influx of predominant plasma HIV-1 clones into the resting CD4+ T cell compartment. 19 –21 The resolution of this controversy is of practical importance in guiding treatment intensification, and has implications for the eradication of HIV-1 infection.
We tested the hypothesis that ongoing HIV-1 replication in the context of prolonged effective ART would result in the redistribution of proviral load among CD4+ T cells with respect to antigen specificities. In acute and chronic untreated HIV-1 infection, as well as in individuals treated with ART for a relatively short duration, it has been demonstrated that HIV-1-specific CD4+ T cells carry disproportionately high proviral burdens as compared to total memory CD4+ T cells and cytomegalovirus (CMV)-specific CD4+ T cells. 22 This is thought to be largely due to the antigen-driven activated state of HIV-1-specific cells in the context of ongoing replication, given that activated CD4+ T cells are much more susceptible to infection than their resting counterparts. 23 –27 Although the majority of these infected cells die, a subset becomes quiescent and contributes to the resting memory CD4+ T cell latent reservoir. We reasoned that the paucity of HIV-1 antigens in subjects treated with effective ART would result in a substantial reduction in this bias, and hence that ongoing low-level replication would be manifested as a more even distribution of proviral burden between HIV-1-specific CD4+ T cells and cells of other specificities. We further posited that if low-level HIV-1 replication was occurring in ARV-treated subjects, CD4+ T cells specific for antigens that were frequently encountered in this context would continue to accumulate proviral load. We were able to test this for influenza-specific CD4+ T cells in vivo by taking advantage of the seasonal vaccination received by our cohort of ARV-treated subjects. Notably, several groups have demonstrated that vaccination with recall antigens, including either influenza or tetanus toxoid, results in the induction of HIV-1 replication in the absence of ART as evidenced by increases in both plasma viremia and proviral loads. 28 –34
Materials and Methods
Subjects and samples
This study was approved by the University of Toronto Institutional Review Board. Subjects were recruited based on the criteria of (1) treated with ART (viral load <50 copies/mm3) for >5 years and (2) received annual influenza vaccinations for >5 years. Clinical data are presented in Table 1. Leukapheresis was performed on subjects prior to and 2 weeks after receiving a standard dose of Fluviral influenza vaccine, a trivalent inactivated vaccine (Shire Biologics).
ART, antiretroval therapy.
Assessing levels of virus production upon antigenic stimulation
Peripheral blood mononuclear cells (PBMCs) were cultured at 5×106 cells/ml in stimulation medium [RPMI+10% human AB serum+1 μg/ml anti-CD28+1 μg/ml anti-CD49d+1 μM zidovudine (AZT)+1 μM lamivudine (3TC)+1 μM nevirapine (NVP)] with either 1 μg/ml cytochrome c (Austral Biologicals), 1 μg/ml HIV-1-p55 (Austral Biologicals), 1 μg/ml tetanus toxoid (List Biological Laboratories), 1/50 dilution of Fluviral influenza vaccine (Shire Biologics), or 0.2 μg/ml staphylococcal enterotoxin B (SEB) (Sigma). Supernatants were collected following 7 days of stimulation and viral RNA was quantified using the AMPLICOR-HIV Monitor test (Roche).
Assessing levels of provirus in antigen-specific cells
CD8+ T cells were depleted from PBMCs using Dynal beads (Invitrogen). The remaining cells were plated at 1×107 cells/ml in stimulation medium with either: cytochrome c, HIV-1-p55, tetanus toxoid, or Fluviral influenza vaccine at the concentrations indicated above, or with a 1/20 dilution of CMV antigen (Virion). These were enriched for antigen-specific cells using a cocktail of the interferon (IFN)-γ, tumor necrosis factor (TNF)-α, and interleukin (IL)-2 cytokine secretion assay enrichment and detection kits (Miltenyi Biotec). We achieved the following purities of phycoerythrin (PE)-labeled cells following enrichment (mean, range): HIV-1-p55—75.8%, 51.2–91.1%, influenza—94.5%, 90.8–96.6%, CMV—71.6%, 43.3–92.3%, and tetanus toxoid—55.3%, 48.6–78.4%. Selected populations were lysed and treated with proteinase K (Roche Applied Science) for 1 h at 56°C followed by heat inactivation. Proviral DNA was quantified by real-time polymerase chain reaction (PCR), as previously described. 35
Statistical analysis
Statistical tests were performed with GraphPad Prism software version 4.0. The tests used for each analysis are indicated in the legend of Fig. 1.
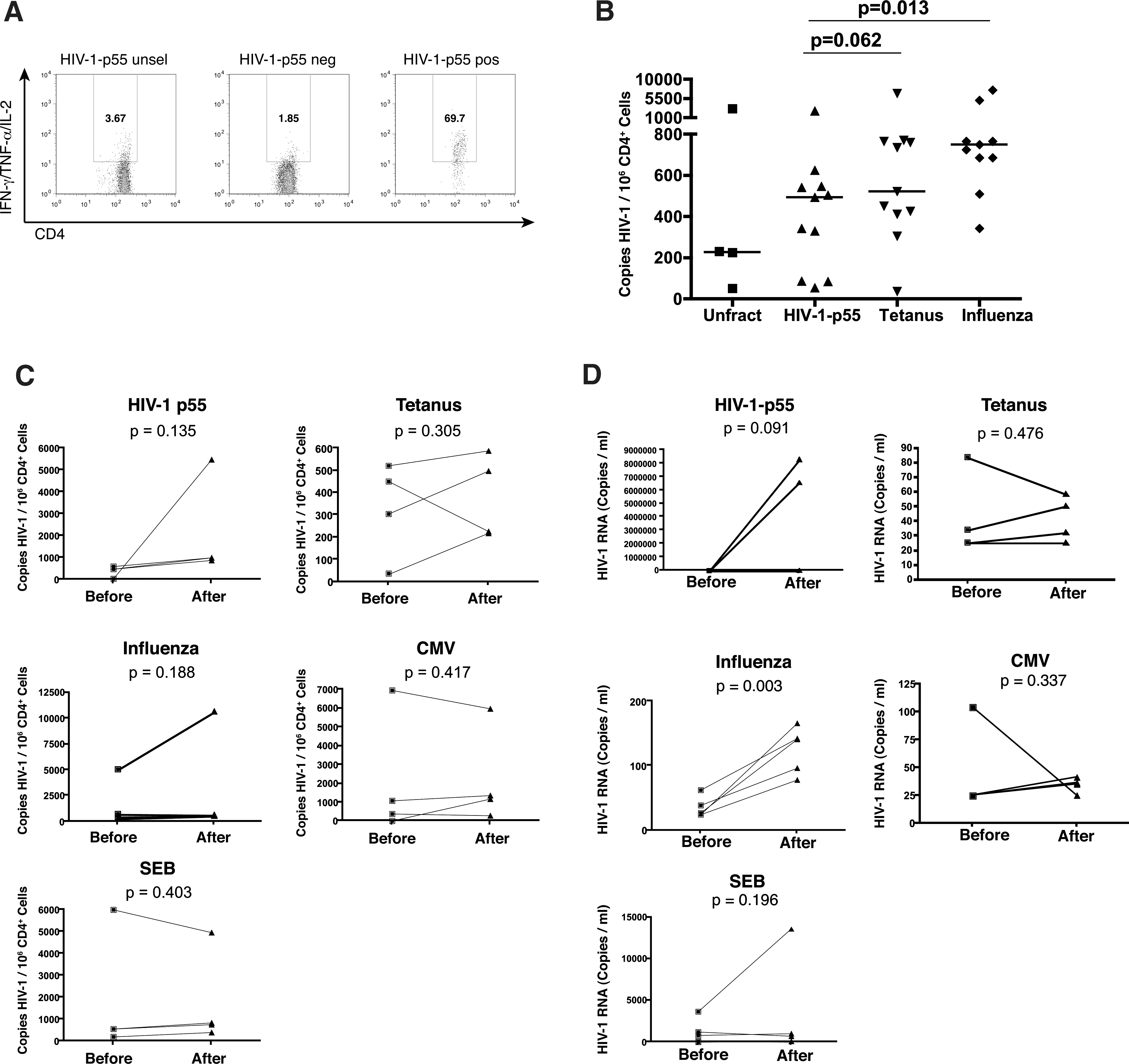
Quantitation of HIV-1 proviral loads and levels of virus production by antigen-specific CD4+ T cells at baseline and 2 weeks postinfluenza vaccination.
Results
PBMCs were obtained from 11 HIV-1-infected subjects who had (1) been on effective ART for >5 years and (2) received annual influenza vaccinations for >5 years. For four of these subjects, samples were collected immediately before and 2 weeks postinfluenza vaccination. Levels of baseline HIV-1 proviral DNA were quantified in sorted HIV-1-p55, tetanus toxoid, and influenza-specific CD4+ T cells. We observed that HIV-1-p55-specific cells carried the lowest proviral burden with means of HIV-1-p55—564.5 copies/106 CD4+ T cells, tetanus toxoid—1,081.8 copies/106 CD4+ T cells, and influenza—1,685.2 copies/106 CD4+ T cells. The differences between proviral loads in HIV-1-p55 versus influenza specific cells were significant (p=0.013), while the differences between HIV-1-p55 and tetanus toxoid were not (p=0.062) (Fig. 1A and B). Thus, in subjects treated with effective ART for prolonged periods of time, influenza-specific CD4+ T cells carry a higher proviral burden than HIV-1-p55-specific CD4+ T cells. Two weeks postinfluenza vaccination we observed trends toward increased proviral loads in HIV-1-p55-specific (mean of 419.3 copies/106 CD4+ T cells prevaccination versus 2,108.1 postvaccination, p=0.135, increases observed in four/four subjects, Fig. 1C) and influenza-specific CD4+ T cells (mean of 1,675.8 copies/106 CD4+ T cells prevaccination versus 3,125.5 postvaccination, increases observed in three/four subjects, p=0.135, Fig. 1C). Neither tetanus toxoid-specific, CMV-specific, nor SEB-responsive CD4+ T cells displayed a consistent trend (tetanus toxoid—mean of 328.1 copies/106 CD4+ T cells prevaccination versus 383.5 postvaccination, p=0.305, CMV—mean of 2,113.2 copies/106 CD4+ T cells prevaccination versus 2,215.8 postvaccination, p=0.417, SEB—mean of 1,839.8 copies/106 CD4+ T cells prevaccination versus 1,775.5 postvaccination, p=0.403, Fig. 1C). Therefore, although we detected trends toward increased HIV-1 proviral burden in influenza and HIV-1-p55-specific CD4+ T cells, these were not significant in our small sample size.
An increase in proviral burden in antigen-specific CD4+ T cells upon vaccination would also manifest as increased viral production upon in vitro antigenic stimulation. This has previously been reported for vaccination with tetanus toxoid. 31 To test this, CD8-depleted PBMCs were stimulated for 7 days in the presence of 3TC, AZT, and nevirapine and levels of HIV-1 RNA in supernatants were quantified. We observed consistently higher levels of virus production upon influenza stimulation in postvaccination versus prevaccination samples (means of 125 versus 35 copies/ml, p=0.003, Fig. 1D). We did not observe a consistent or significant increase in levels of virus production following influenza vaccination upon stimulation with tetanus toxoid, CMV, or SEB (Fig. 1D).
Stimulation with HIV-1-p55 produced a striking result, where postinfluenza vaccination samples from two subjects produced very high levels of HIV-1 RNA in the range of millions of copies/ml. Two other subjects exhibited modest increases in virus production upon HIV-1-p55 stimulation postinfluenza vaccination and one exhibited a decrease. Thus, influenza vaccination enhanced virus production from both HIV-1 and influenza-specific cells, after restimulation with their respective antigens. This effect was antigen specific as we did not observe increased levels of virus production postvaccination upon in vitro stimulation with tetanus toxoid or CMV.
Discussion
We observed that in subjects treated with ART for prolonged periods of time (>5 years), influenza-specific CD4+ T cells carried a substantially higher proviral burden than their HIV-1-p55-specific counterparts. This is in contrast to the disproportionately high levels of proviral DNA in HIV-1-specific CD4+ T cells reported in untreated HIV-1 infection and in subjects treated with ART for relatively short durations. 22 This supports the idea that the recurrent stimulation of the influenza-specific CD4+ T cells population, through annual vaccination and influenza infection, renders them susceptible to infection by low levels of ongoing HIV-1 replication, resulting in the accumulation of proviral DNA. We tested this in vivo by determining whether we could observe an increase in HIV-1 proviral load in influenza-specific CD4+T cells in ARV-treated subjects following a single influenza vaccination. Although we did observe a trend toward an increased level of provirus in influenza-specific cells, which was not observed in tetanus toxoid or CMV-specific cells, this did not reach statistical significance. This may reflect the inadequate statistical power of our sample size to detect a relatively small in vivo increase in proviral levels over a single round of vaccination. This is supported by our observation of significantly higher levels of proviral DNA in influenza-specific cells in baseline than cells of other specificities, consistent with a cumulative effect over multiple years of annual vaccination.
We observed significantly higher levels of HIV-1 production upon in vitro influenza stimulation in postvaccination versus prevaccination samples. Since these stimulations were performed in the presence of 3TC, AZT, and nevirapine it is unlikely that de novo infection of vaccine-primed influenza-specific cells occurred in vitro. We propose that this likely reflects in vivo infection of vaccine-primed influenza-specific CD4+ T cells, and thus a larger pool of infected influenza-specific CD4+ T cells at initiation of the in vitro culture. This is supported by the trends toward higher levels of provirus in influenza-specific CD4+ T cells observed following vaccination. As the production of HIV-1 over 7 days comprises an amplification step, this may have improved our ability to detect a relatively small in vivo increase in infected influenza-specific CD4+ T cells following a single vaccination. We cannot rule out, however, an indirect effect whereby higher levels of cytokine production postvaccination may drive the production of HIV-1 from non-influenza-specific cells present in the culture.
It is also of interest to consider the possibility of a secondary effect of influenza vaccination, whereby the induction of HIV-1 replication by influenza vaccination activates HIV-1-specific CD4+ T cells and recruits them to sites of replication, rendering these more susceptible to infection. We observed a trend toward increased levels of HIV-1 proviral DNA in HIV-1-p55-specific CD4+ T cells upon influenza vaccination. This does not appear to agree, however, with our data showing relatively low levels of provirus in HIV-1-p55-specific CD4+ T cells at baseline. It is conceivable that HIV-1-p55-specific CD4+ T cells become infected upon influenza vaccination, due to such a secondary effect, but are less prone to returning to a resting memory state. In this regard, it is interesting to note that two subjects exhibited extraordinarily high levels of HIV-1 production upon HIV-1-p55 stimulation of postvaccination samples. We suggest that cells producing such high levels of HIV-1 are unlikely to survive and return to a resting state. The reason for this high level of HIV-1 production from HIV-1-p55-specific cells is presently unclear and will be the subject of future studies.
Our data add to the evidence that low-levels of HIV-1 replication occur in ARV-treated subjects following antigenic stimulation, comprising a barrier to eradication. Our findings should not deter current recommendations for routine vaccination of HIV-1-infected individuals for prevention of influenza or pneumococcal pneumonia. 36 We and others have previously demonstrated that although vaccinations can transiently enhance HIV-1 viral replication due to vaccine-induced immune activation, the immune protection conferred by vaccine-induced antibodies will prevent the more prolonged immune the activation and enhanced viral replication that can result from the full blown infections that the vaccines were meant to prevent. 30,31,33,37 In this article we show that although vaccination can prevent the deleterious effects of acute bacterial and viral infections, HIV-1 can still exploit newly primed antigen-specific memory cells resulting from vaccination for its own use, by establishing new reservoirs. However, the overall effect of vaccination may still be to limit HIV-1 viral reservoirs in the long run, at the expense of infecting a small population of vaccine-specific cells.
Footnotes
Acknowledgments
Funding for this work was obtained from the Canadian Institutes of Health Research. The following reagents were obtained from the NIH AIDS Research and Reference Reagent Program, Division of AIDS, NIAID, NIH: nevirapine and 3TC. R.B.J. gratefully acknowledges salary support from the Ontario HIV Treatment Network (OHTN). Biosafety level 3 laboratory space was provided by the Canadian Foundation for HIV Research (CANFAR) in partnership with the Ontario Innovation Trust and the Canadian Foundation for Innovation.
Author Disclosure Statement
No competing financial interests exist.