
Abstract
Cell surface heparan sulfate proteoglycans (HSPGs) are involved in the binding and entry of human T-cell leukemia virus type 1 (HTLV-1) into host cells, while sulfated polysaccharides such as heparin inhibit HTLV-1 infection. Chondroitin sulfate proteoglycans (CSPGs) are classified as another major type of proteoglycans. Here, we examined the effect of four types of chondroitin sulfate (CS) on HTLV-1 infection. Accordingly, a human T cell line, MOLT-4, was inoculated with cell-free HTLV-1 in the presence or absence of soluble CS, and the synthesis of reverse-transcribed HTLV-1 DNA within cells 20 h after inoculation was detected using polymerase chain reaction (PCR). Among the four types of CS (A, C, D, and E), the E type (CSE), which was derived from the squid cartilage, exhibited anti-HTLV-1 activity. Furthermore, we observed that CSE directly interacted with recombinant HTLV-1 envelope (Env) proteins and inhibited the binding of HTLV-1 virions to MOLT-4 cells, indicating that the interaction between Env and CSE plays a significant role in its anti-HTLV-1 activity. In addition, CSE inhibited syncytium formation that was induced by HTLV-1-producing cells. When CSE was mixed with the synthetic fusion inhibitor peptide corresponding to the ectodomain of the Env transmembrane subunit (TM) gp21, the HTLV-1 infection was further inhibited when compared with the inhibitory effect of each compound alone. Thus, further elucidation of the in vitro antiviral mechanism of CSE shown in this study will lead to the development of CSE-like molecules for the entry inhibition of HTLV-1.
Introduction
P
Human T-cell leukemia virus type 1 (HTLV-1) is an oncogenic retrovirus that induces adult T cell leukemia/lymphoma (ATL) and HTLV-1-associated myelopathy or tropical spastic paraparesis (HAM/TSP). 4 HTLV-1 primarily infects CD4+ T cells in vivo. 5 HTLV-1 infection among humans occurs through the transmission of infected cells harboring integrated HTLV-1 provirus via breast feeding, the transmission of contaminated blood, and sexual contact. HTLV-1 infection is endemic in southwestern Japan, the Caribbean Basin, Central Africa, South America, Papua New Guinea, and the Solomon Islands with an estimated 10–20 million HTLV-1-infected individuals worldwide. 6
Viral envelope (Env) glycoproteins, surface subunit (SU) gp46 and transmembrane subunit (TM) gp21, are required for HTLV-1 infection of target cells. 7 HTLV-1 gp46 (SU) binds to the cell surface and directly interacts with the cell surface receptors, while gp21 (TM) plays a significant role in the fusion between viral and cellular membranes. Glucose transporter 1 (GLUT-1), 8,9 neuropilin-1 (NRP-1), 10 –12 and heparan sulfate proteoglycans (HSPGs) 13 –15 are molecules that directly participate in HTLV-1 entry, and their respective roles in HTLV-1 infection have been reviewed in detail. 16 –18
HSPGs function as attachment factors for HTLV-1 entry. More recently, we reported that HSPGs dramatically enhance HTLV binding and entry into target cells and identified that the specific forms of HSPGs (with syndecan-1 or syndecan-2 core proteins) enhance the susceptibility of cells to HTLV-1. 19 Furthermore, critical roles for HSPGs in transmission and infection of HTLV-1 mediated through dendritic cells (DC) have also been investigated: (1) blocking HSPGs on monocyte-derived DCs inhibited HTLV-1 binding 20 and (2) blocking HSPGs on the surface of CD4+ T cells inhibited the HTLV-1 infection of T cells through DC. 21
CSPGs represent another major proteoglycan type. CS chains are composed of repetitive units of disaccharides, consisting of glucuronic acid (GlcA) and N-acetylgalactosamine (GalNAc). GlcA can be O-sulfated at C2 and rarely at C3, whereas GalNAc can present sulfate groups at C4 and/or C6. Several types of CS chains can be distinguished based on the disaccharide units characterized by sulfation at specific positions. CS chains are divided into at least four subclasses (Fig. 1): CSA [GlcAβ1-3GalNac(4S)], CSC [GlcAβ1-3GalNAc(6S)], CSD [GlcA(2S)β1-3GalNAc(6S)], and CSE [GlcAβ1-3GalNAc(4S,6S)]. The sulfation patterns determine the functional interaction of CS with growth factors, cell surface receptors, and other components of the extracellular matrix. 22 CS chains that contain disulfated disaccharide units, such as CSD or CSE, are frequently isolated from marine organisms. Squid cartilage-derived CSE is a potent inhibitor of herpes simplex virus (HSV), 23 whereas CSA, CSC, and CSD exhibit little or no anti-HSV activity. 24,25 More recently, Kato et al. 26 reported that CSE inhibits dengue virus infection.

Structures of the chondroitin sulfate (CS) and heparin (Hep) molecules used in this study. CS types A, C, D, and E are indicated by CSA, CSC, CSD, and CSE, respectively. Mr indicates an average molecular weight.
In this study, we examined the effects of the several types of soluble CS on HTLV-1 infection. In a previous study, we detected reverse-transcribed HTLV-1 DNA in target cells infected with a cell-free virus using an HTLV-1-specific polymerase chain reaction (PCR) assay, 27 although the infection of cells with cell-free HTLV-1 is quite inefficient when compared with other retroviruses. 28 HTLV-1-specific PCR bands are detectable 1 day after infection, and the formation of reverse-transcribed DNA is inhibited after treatment with HTLV-1-specific neutralizing antibodies. 27,29 Using this PCR assay system, we observed that CSE strongly inhibited the early stage of HTLV-1 infection.
Materials and Methods
Materials
CS, isolated from whale cartilage (CSA), shark cartilage (CSC and CSD), and squid cartilage (CSE), and heparin, isolated from pig intestine, were all purchased from the Seikagaku Corp. (Tokyo, Japan). Complexes of sulfated GAGs with dipalmitoylphosphatidylethanolamine (PE) (sulfated GAG-PE) were prepared as previously described at the Institute for Molecular Science of Medicine, Aichi Medical University. 30,31 The synthetic peptides used in this study were purchased from Hokkaido System Science (Sapporo, Japan). The purity of the peptides was more than 95%. The monoclonal antibody against human CD4 (NU-TH/I) was purchased from Nichirei, Tokyo, Japan.
Cells
MOLT-4 clone 8 cells 32 are a human T-cell target cell line for virus inoculation. U251-MG cells are derived from a human astrocytoma. 33 c77 is a subclone of 8C feline kidney cells 34 that was cocultivated with lethally irradiated ATL-2M cells. 35 C91/PL cells were established from cord lymphocytes cocultivated with the leukemic cells from an ATL patient. 36 c77 and C91/PL cells were used as HTLV-1-producing cells. MOLT-4 cells persistently infected with T-cell line-tropic human immunodeficiency virus type 1 (HIV-1) (IIIB strain), MOLT-4/IIIB cells, 37 were used as HIV-1-producing cells. MOLT-4, C91/PL, and MOLT-4/IIIB cells were maintained in RPMI 1640 medium supplemented with 10% fetal calf serum (FCS). c77 and U251-MG cells were maintained in Eagle's minimum essential medium supplemented with 10% FCS. All cells were maintained at 37°C in a humidified, 5% CO2 atmosphere.
Virus preparation
To prepare cell-free HTLV-1 or HIV-1 samples, c77 and MOLT-4/IIIB cells, respectively, were seeded, and the culture supernatants were harvested, centrifuged at a low-speed on the following day, and subsequently passed through 0.45-μm cellulose acetate syringe filters (Pall, Port Washington, NY). The virus samples were stored at −80°C until use.
Cell-free virus inoculation and detection of reverse-transcribed viral DNA by PCR
MOLT-4 cells were inoculated with cell-free HTLV-1 or HIV-1 in the presence or absence of the sulfated GAGs for 1 h at 37°C, and washed twice with phosphate-buffered saline (PBS), and subsequently fresh medium was added. After incubation for 20 h, the cells were washed and lysed with 10 mM Tris–HCl (pH 8.3) containing 1 mM ethylenediaminetetraacetic acid (EDTA) (Dojindo, Kumamoto, Japan), 0.45% Nonidet P-40 (Sigma, St. Louis, MO), 0.45% Tween 20 (Sigma), and 0.2 mg/ml proteinase K (Sigma). The cell lysates were incubated for 2 h at 52°C and heated for 10 min at 96°C to inactivate proteinase K. To detect the formation of reverse-transcribed HTLV-1 DNA, PCR was performed with HTLV-1-specific pX primers under the following conditions: 30 cycles at 93°C for 1 min, 67°C for 45 s, and 72°C for 1 min, and one cycle at 72°C for 5 min. PCR primers with the following nucleotide sequences were used: pX 7302-7326, 5'-CCCACTTC CCAGGGTTTGGACAGAG-3' and pX 7504-7481, 3'-CTGTA GAGCTGAGCCGATAACGCG-5'.
For the detection of reverse-transcribed HIV-1 DNA, PCR was performed with the LTR primers M667 and AA55 38 under the following conditions: 30 cycles at 93°C for 1 min, 60°C for 45 s, and 72°C for 1 min, and one cycle at 72°C for 5 min. PCR primers with the following nucleotide sequences were used: M667, 5'-GGCTAACTAGGGAACCCACTG-3' and AA55, 3'-CTGCTAGAGATTTTCCACACTGAC-5'.
The human β-globin gene primers KM29 and KM38 39 were used as an internal control. The nucleotide sequences for these PCR primers are indicated as follows: KM29, 5'-GGTTGG CCAATCTACTCCCAGG-3' and KM38, 5'-TGGTCTCCTTA AACCTGTCTTG-3'.
The PCR products were visualized using electrophoresis on 2% agarose gels containing 0.5 μg/ml ethidium bromide. The intensity of specific PCR signals was measured using a densitometer (ATTO Co., Tokyo, Japan). The inhibitory effect of the test samples was determined through a comparison analysis of the PCR band intensities obtained after infection with virus in the presence of sulfated GAGs and the bands were obtained after infection with diluted virus. The intensities of the PCR bands for the 1/2 diluted virus samples were used to estimate the concentrations at 50% inhibition (IC50) of sulfated GAGs.
HTLV-1 binding or postbinding assay
MOLT-4 cells incubated at 4°C (precooled MOLT-4 cells) were inoculated with cell-free HTLV-1 at 4°C for 1 h in the presence or absence of CSE, washed with cold PBS to remove unbound virus, and cultured at 37°C for 20 h to examine the effects of CSE on virus binding. To examine the effects of CSE on postbinding, precooled MOLT-4 cells were inoculated with HTLV-1 at 4°C for 1 h, washed with cold PBS, and cultured at 37°C for 20 h in the presence or absence of CSE. Subsequently, the formation of HTLV-1 DNA was detected using PCR as described above.
In addition to the PCR assay, c77 cell culture supernatants (1 ml) were centrifuged at 20,000×g for 90 min at 4°C to generate concentrated HTLV-1 viral particles for the detection of HTLV-1 particles bound to the cell surface. We confirmed that the majority of HTLV-1 virions in the culture supernatants could be precipitated under these conditions. The supernatants were removed and the precipitates were resuspended with 200 μl of PBS. Subsequently, the precooled MOLT-4 cells were incubated with HTLV-1 virions in the presence or absence of CSE at 4°C for 1 h, washed, and lysed with sodium dodecyl sulfate (SDS) sample buffer containing 625 mM Tris–HCl (pH 6.8), 2% SDS, 10% glycerol, 50 mM dithiothreitol (DTT), and 0.25% bromophenol blue. The lysates were boiled for 5 min, subjected to SDS-polyacrylamide gel electrophoresis (PAGE), and transferred to Immobilon PVDF transfer membranes (Millipore, Bedford, MA). The membranes were then blocked with PBS containing 0.05% Tween 20 (PBST) with 5% nonfat dry milk (blocking buffer) for 2 h at 37°C and incubated with anti-HTLV-1 Gag (p24Gag) mouse monoclonal antibody (MAb) (clone 6G9) (Santa Cruz Biotechnology Inc., Santa Cruz, CA) overnight at 4°C. The bound antibodies were detected with horseradish peroxidase (HRP)-conjugated rabbit antimouse immunoglobulins (DAKO, Glostrup, Denmark) using the enhanced chemiluminescence (ECL) system (GE Healthcare, Buckinghamshire, UK). As an internal control, the membrane was reprobed with an anti-β-tubulin MAb (Sigma).
Binding of HTLV-1 to sulfated GAG-PE
The wells of 96-well microtiter plates (Corning, Lowell, MA) were coated with sulfated GAG-PE (50 μg/ml in PBS) overnight at 4°C, washed twice with PBS, and blocked with 5% bovine serum albumin (BSA) in PBS for 2 h at room temperature. Subsequently, HTLV-1 particles prepared as described above were added to the wells, incubated with GAG-PE overnight at 4°C, and washed three times with PBS. To detect the HTLV-1 particles bound to sulfated GAG-PE, SDS sample buffer was added to the wells to lyse the HTLV-1 followed by western blotting to detect p24Gag as described above.
Binding of sulfated GAGs to a recombinant HTLV-1 Env protein
The wells of 96-well microtiter plates were coated with 10 μg/ml of Escherichia coli-derived recombinant HTLV-1 Env protein [C-terminal half of gp46 (aa 165–312) and most of gp21 (aa 313–440)] (>95% pure) (Jena Bioscience, Jena, Germany) overnight at 4°C, washed twice with PBS, and blocked with 5% BSA in PBS for 2 h at room temperature. Subsequently, soluble sulfated GAGs (10 μg/ml) diluted with 5% BSA in PBST was added and incubated for 2 h at room temperature. Avidin binds to heparin, 40 CSD, and CSE (our unpublished results), therefore HRP-labeled avidin (DAKO) diluted 1:2,000 with 5% BSA in PBST was added to each well to detect bound sulfated GAGs. After incubation for 2 h at room temperature, a 3,3′,5,5′-tetramethylbenzidine substrate solution (Bio-Rad, Hercules, CA) was added for 5 min at room temperature to develop the color, and 1 N H2SO4 was added to terminate the reaction. The optical densities (OD) at 450 nm (OD450) were measured.
Syncytium formation assay
U251-MG cells (2×104 cells per well) were seeded into the wells of 96-well plates and cultured overnight. The following day, HTLV-1-producing C91/PL cells (2×103 cells per well) were incubated with either PBS or sulfated GAGs for 10 min at 37°C and subsequently overlaid onto U251-MG cells. The cocultured cells were incubated at 37°C for 16 h, fixed with methanol, and stained with Giemsa's solution. The syncytia were then counted and the number of syncytia formed in control cultures was approximately 300 per well for each experiment. The inhibitory activity of sulfated GAGs against syncytium formation was calculated using the following formula: inhibition (%)=[(the number of syncytia formed without sulfated GAGs) – (the number of syncytia formed with sulfated GAGs)]/(the number of syncytia formed without sulfated GAGs)×100.
Results
Inhibitory effect of CSE on early stage of HTLV-1 infection before or during reverse transcription
The human T cell line MOLT-4 was inoculated with cell-free HTLV-1, and cell lysates were prepared 20 h later to detect the formation of reverse-transcribed HTLV-1 DNA using PCR. The expected pX DNA fragment (203 bp) was detected (Fig. 2A, lane 1). This cell-free HTLV-1 infection was specifically neutralized after pretreatment of HTLV-1 with the human serum positive for HTLV-1 (lane 2) but not with the human serum negative for HTLV-1 (lane 3). We next examined the effects of four types of CS (CSA, CSC, CSD, and CSE) on the formation of HTLV-1 DNA, namely, on the early stage of cell-free HTLV-1 infection. The formation of HTLV-1 DNA was detected to estimate the inhibitory effect of CS (Table 1). Both CSA and CSC did not have much of an inhibitory effect on the formation of HTLV-1 DNA, whereas CSE strongly inhibited viral DNA formation (Table 1 and Fig. 2B). A weaker inhibitory effect of CSD was also detected (Table 1). Consistent with a previous report, 13 the inhibitory effect of heparin was also detected in our assay system (Table 1). Thus, our results suggested that CSE inhibited HTLV-1 infection at an early stage before or during reverse transcription.

Inhibitory effect of CSE on the early stage of human T-cell leukemia virus type 1 (HTLV-1) infection.
MOLT-4 cells were inoculated with cell-free HTLV-1 or HIV-1 in the presence of one of sulfated GAGs, incubated for 20 h and examined for the synthesis of reverse-transcribed viral DNA within cells by PCR assays as described in Materials and Methods. The intensities of specific PCR bands were measured and 50% inhibitory concentrations (IC50) of each sulfated GAG were determined. Experiments were repeated three times and data are represented as means±SD.
To investigate the specificity of the anti-HTLV-1 activity of CSE, we examined the effect of soluble CS on cell-free HIV-1 (X4 virus) infection. After inoculation of HIV-1 to MOLT-4 cells, the reverse-transcribed HIV-1 DNA was detected using PCR with a primer pair specific for the LTR region (140 bp) (Fig. 2C). We confirmed that the pretreatment of MOLT-4 cells with the anti-CD4 antibody before inoculation specifically inhibited the formation of HIV-1 DNA (Fig. 2C, lane 2). As shown in Fig. 2D, in contrast to HTLV-1, CSE had little inhibitory effect on HIV-1 infection; however, heparin effectively inhibited HIV-1 infection (Table 1), and this result is consistent with that of a previous report. 2,37 The strong anti-HTLV-1 activity of CSE did not reflect its cell toxicity, as the slight effect of CSE (400 μg/ml) on the growth and viability of MOLT-4 cells was not observed until 2 days after treatment (data not shown).
Inhibition of the binding of HTLV-1 to MOLT-4 cells by CSE
Retroviruses bind to the cell surface at 4°C (binding step), but the postbinding steps such as membrane fusion or internalization are temperature dependent. 41 –45 Our results suggested that CSE inhibited HTLV-1 infection at an early stage before or during reverse transcription, therefore we examined whether CSE inhibited binding or postbinding (Fig. 3). For the examination of the binding step, precooled MOLT-4 cells were incubated with cell-free HTLV-1 at 4°C for 1 h and washed. Subsequently the temperature was shifted to 37°C to allow viral entry into the cells to examine the postbinding. As shown in Fig. 3A (lower panel), the addition of CSE at 4°C (binding step: lanes 2 and 3), but not at 37°C (postbinding step: lanes 4 and 5), clearly inhibited the synthesis of HTLV-1 DNA, indicating that CSE inhibited the binding step of HTLV-1 infection.
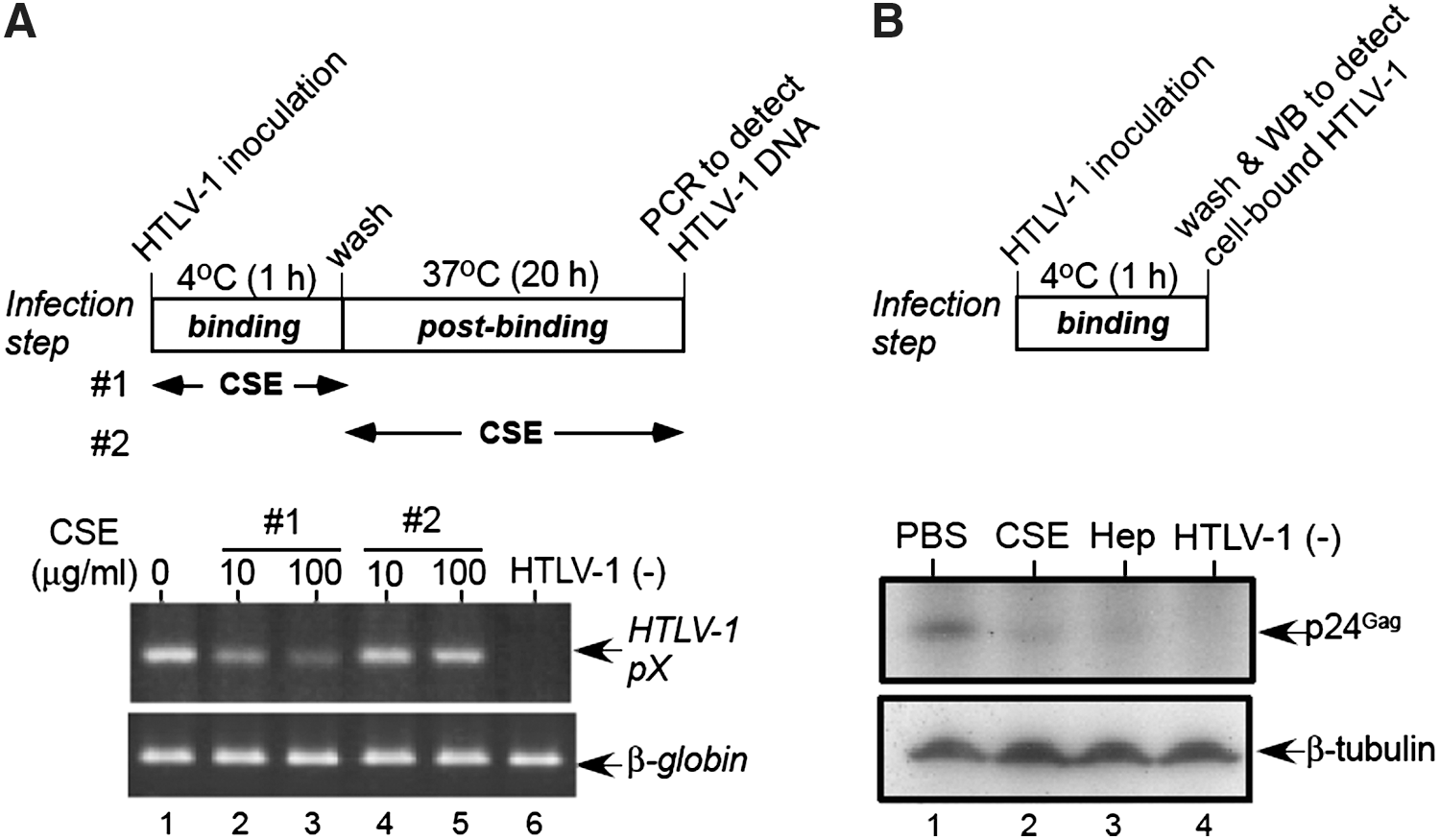
Inhibition of binding of HTLV-1 to MOLT-4 cells by CSE
To confirm this result, western blotting was performed to detect the HTLV-1 viral particles bound to the cell surface (Fig. 3B). Heparin has been shown to block the binding of HTLV-1 to CD4+ T-cells, 13 therefore, we used heparin as a specific binding inhibitor (lane 3). Similarly, CSE also inhibited the binding of HTLV-1 to MOLT-4 cells (lane 2), indicating that CSE targets the binding step of HTLV-1 infection for antiviral activity.
Interaction of CSE with HTLV-1 Env
We also determined whether sulfated GAGs interact with HTLV-1 viral particles. We used sulfated GAGs coupled to PE because PE is a hydrophobic molecule, sulfated GAG-PE can be effectively immobilized onto the surface of plastic plates through this carrier lipid (PE) 30,31 to analyze the direct interaction between the virus and sulfated GAGs. The wells of microtiter plates were coated with sulfated GAG-PEs, and HTLV-1 virions were added. The bound HTLV-1 was subjected to western blotting to detect the p24Gag protein. As shown in Fig. 4A, the binding of HTLV-1 to CSE-PE (lane 5) and heparin-PE (Hep-PE) (lane 6) was clearly detected. In addition, the weak binding of HTLV-1 to CSD-PE (lane 4) was also detected. Thus, the anti-HTLV-1 activities of sulfated GAGs are well correlated with their binding affinity for HTLV-1.
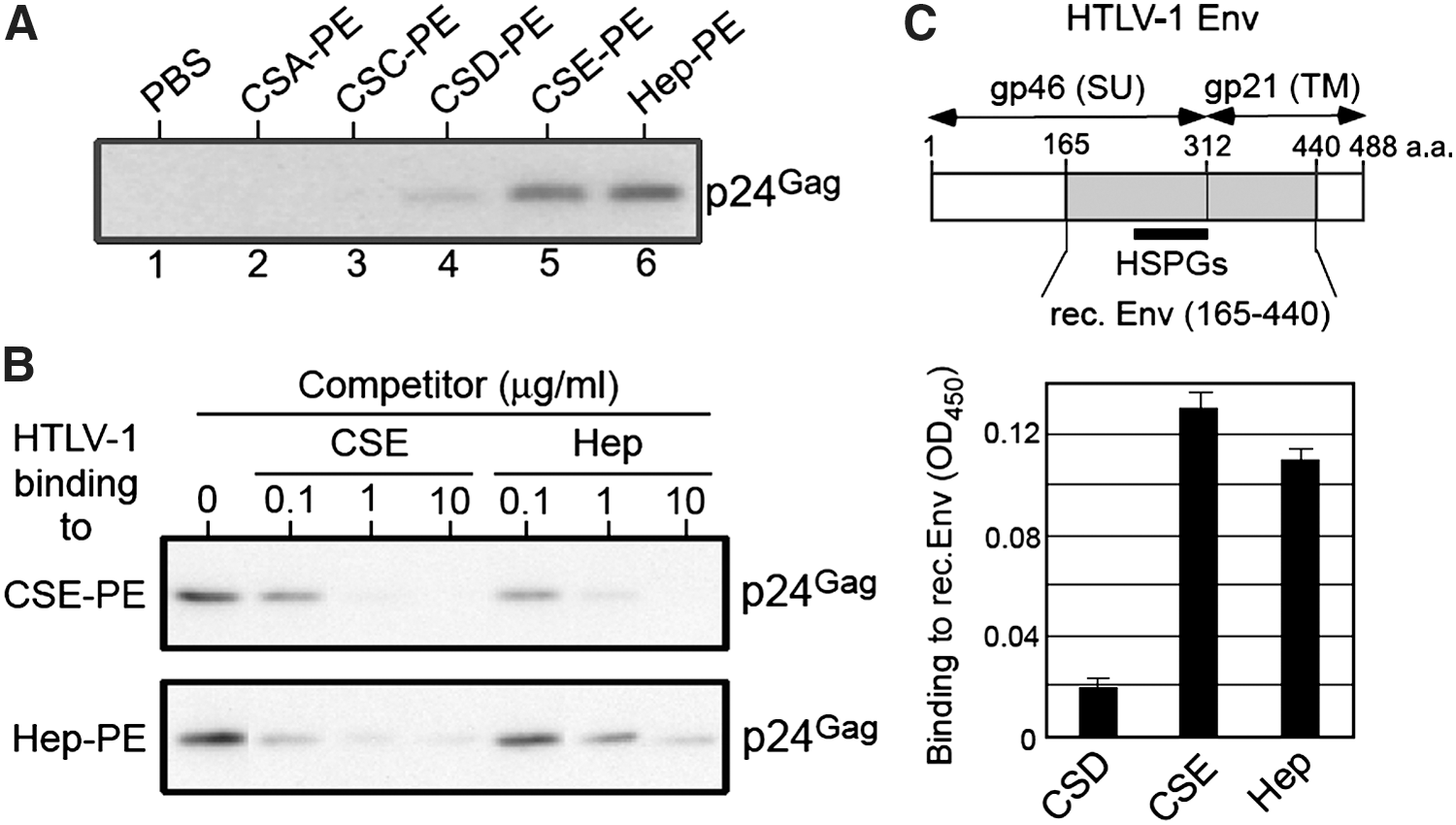
Binding of CSE to HTLV-1 Env protein.
Both CSE and heparin are highly sulfated polysaccharides, therefore we determined whether soluble CSE or heparin cross-inhibits the binding of HTLV-1 virions to CSE-PE or Hep-PE. As shown in Fig. 4B, the binding of HTLV-1 to CSE-PE or Hep-PE was blocked in the presence of CSE or heparin in a dose-dependent manner, suggesting that a common region in HTLV-1 Env might be involved in its interaction with CSE or heparin.
HTLV-1 Env binds to HSPGs present on the surface of cells. 13 –15 Moreover, heparin is structurally similar to HS. Therefore, we examined whether heparin binds to recombinant HTLV-1 Env protein produced in E. coli, which lacks the N-terminal region (aa 1–164) but contains the major portion of the gp46 (SU) that is involved in the interaction of HTLV-1 Env with HSPGs (aa 215–312). 18,46 The results shown in Fig. 4C revealed that both heparin and CSE bound to the recombinant Env protein, suggesting that the HSPGs-binding domain in HTLV-1 Env is also involved in its interaction with CSE.
Effect of sulfated GAGs on syncytium formation induced by HTLV-1-producing cells
We performed syncytium formation assays to examine the effects of sulfated GAGs on the interactions of HTLV-1 Env with cell receptors that lead to fusion. HTLV-1-positive human T cells, C91/PL, and HTLV-1-negative human glioma-derived cells, U251-MG, were cocultured. Both CSE and heparin markedly inhibited syncytium formation in a dose-dependent manner (Fig. 5). Fifty percent inhibition (IC50s) was observed with CSE and heparin at 200 μg/ml and 17 μg/ml, respectively.
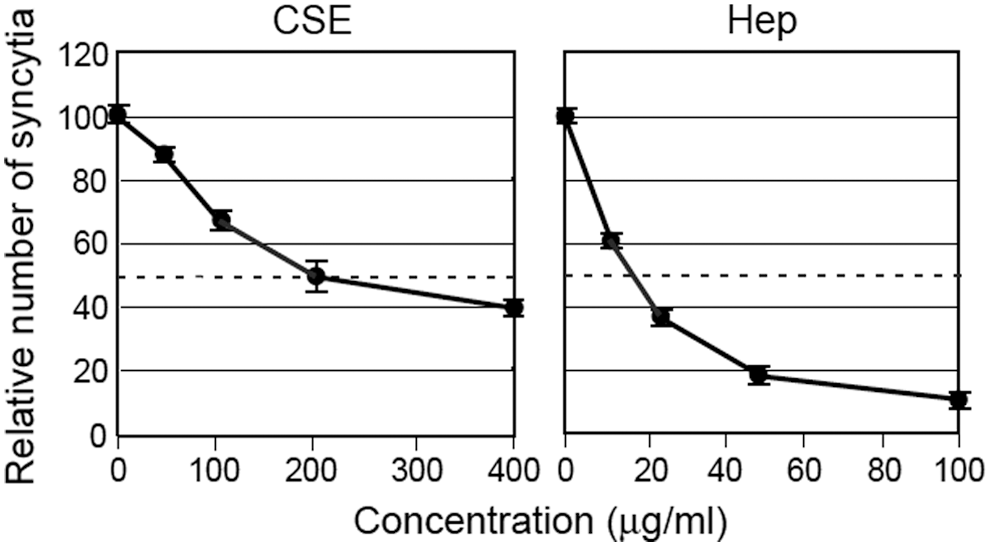
Effect of sulfated GAGs on the formation of HTLV-1-induced syncytia. U251-MG cells were cocultured for 16 h with C91/PL cells in the presence of varying CSE or heparin concentrations. The number of syncytia was counted through microscopic examination. The inhibitory effect of each sulfated GAG (%) was calculated in comparison with the average number of syncytia (∼300) in triplicate control wells (PBS). The data are represented as the mean±SD.
Inhibition of cell-free HTLV-1 infection by CSE in combination with a fusion inhibitor peptide derived from the HTLV-1 Env gp21 (TM)
We have previously reported that a synthetic peptide (pep.) derived from the ectodomain of the gp21 Env transmembrane (TM) protein corresponding to amino acids 400–429 (gp21 pep. 400–429) inhibits the infection of cell-free HTLV-1 at a postbinding (fusion) step. 29 Our results indicated that CSE inhibits HTLV-1 infection at the binding step (Fig. 3): therefore, we examined the effects of CSE in combination with the fusion inhibitor gp21 peptide. The estimated IC50 of the formation of reverse-transcribed HTLV-1 DNA through the gp21 pep. was achieved at 20 μg/ml. 29 We examined the combinatory effect of CSE and the gp21 pep. on cell-free HTLV-1 infection. As shown in Fig. 6, this combination further inhibited HTLV-1 infection, compared with the individual blocking effects of each compound.

Inhibition of cell-free HTLV-1 infection by combination of CSE with the fusion inhibitor peptide derived from the HTLV-1 Env gp21 (TM). MOLT-4 cells were inoculated with cell-free HTLV-1 in the presence of CSE (lane 2), the synthetic peptide derived from the ectodomain of the gp21 (TM) (aa 400–429) (gp21 pep.) (lane 3), or both (lane 4). The formation of reverse-transcribed HTLV-1 DNA in MOLT-4 cells was detected using PCR. β-Globin DNA was amplified as a control. The relative intensities of the pX DNA bands were determined using densitometry (lower panel). The band intensities of pX DNA were divided by those corresponding to the β-globin DNA bands. Three independent experiments generated similar results.
Discussion
In this study, we examined the effects of several types of CS on HTLV-1 infection. We observed that both CSE and CSD inhibited HTLV-1 infection, although the antiviral effect of CSD is weak. Both CSD and CSE are highly sulfated (two sulfates/disaccharide), while CSA and CSC are both monosulfated (Fig. 1). Therefore, not only the degree of sulfation but also the position of sulfates (at positions C4 and C6) in the predominant disaccharide units of CSE might be critical for its antiviral activity against HTLV-1 infection.
HSPGs function as attachment factors for HTLV-1 entry. 13 –15 In the present study, we observed that CSE inhibits the binding step of HTLV-1 infection. We also showed that CSE directly binds to the recombinant Env protein, which includes the HSPGs binding domain (residues 215–312) 18,46 in the C-terminal portion of gp46 (SU). Because soluble CSE inhibited the binding of HTLV-1 to Hep-PE, which is a structural homologue of HS (Fig. 4B), we proposed that CSE might inhibit HTLV-1 through direct binding to gp46 (SU) Env, particularly at the HSPGs binding domain to prevent the interaction of HTLV-1 Env with host cell surface HSPGs, resulting in the blocking of virus binding to target cells.
Cell surface HSPGs also interact with HIV-1 Env. 47 –49 However, CSE only weakly inhibited HIV-1 infection when compared with heparin when MOLT-4 cells were used as a target (Table 1 and Fig. 2D). The low inhibitory effect of CSE on HIV-1 infection remains unknown. However, we observed that heparin and CSE similarly bind to HIV-1 (data not shown), suggesting that the region of HIV-1 that interacts with CSE does not serve a role in HIV-1 attachment to MOLT-4 cells.
CSE also inhibited syncytium formation that was induced by HTLV-1-producing cells, but this inhibitory effect was relatively inefficient when compared with the antiviral activity against cell-free HTLV-1 infection. This result might reflect the difference between the cell-free virus and virus-producing cells as the number of Env molecules expressed in HTLV-1-infected cells is apparently much more abundant than that in viral particles. Alternatively, it has been reported that the assembly of a multimolecular complex, called the virological synapse, plays an important role in the cell-to-cell spread of HTLV-1. 50 Because heparin (∼10 kDa) inhibited syncytium formation more efficiently than CSE (∼60 kDa), the size of the sulfated polysaccharide is an important factor in the inhibition of syncytium formation through infiltration into narrow intercellular spaces (i.e., virological synapse).
In addition, we observed that a mixture of CSE and the synthetic fusion inhibitor peptide (gp21 peptide 400–429) further inhibited HTLV-1 infection compared to the effects of each compound alone (Fig. 6). Thus, due to the multistep nature of virus entry, the combination of inhibitory agents targeting different entry steps might be an advantageous approach to inhibit the entry of HTLV-1 into host cells.
The treatment of HTLV-1-associated diseases remains difficult, and therapeutic strategies targeting HTLV-1 replication are unavailable. In this study, we demonstrated that CS type E inhibits HTLV-1 infection. Further elucidation of the structural basis of the antiviral effect of CSE will contribute to the development of potent entry inhibitors, such as CSE, to prevent HTLV-1 infection.
Footnotes
Acknowledgments
The authors would like to thank Ms. Nakamura for her excellent technical assistance. This work was supported in part through Grants-in-Aid from the Japanese Society for the Promotion of Science and grants from the Japan Health Sciences Foundation and the 21st Century COE Program from the Ministry of Education, Culture, Sports, Science and Technology of Japan.
Author Disclosure Statement
No competing financial interests exist.