
Abstract
Myeloid dendritic cells (mDCs) are essential for initiation of adaptive immune responses but are depleted in HIV infection. Evidence suggests that apoptosis mediates loss, and to further understand the pathways involved, expression of caspases mediating apoptosis via the extrinsic and intrinsic pathways was analyzed. Blood samples were obtained from 14 HIV-infected patients (nine HAART and five antiretroviral naive) and 10 healthy controls. The expression of intracellular active caspases 8 and 9, associated with extrinsic and intrinsic pathways of apoptosis, and the expression of cell membrane death receptors and their ligands were assessed by flow cytometry in mDC. Additionally, expression of active caspases 8 and 9 in purified mDCs cultured for 5 days with HIV-Bal was analyzed. Frequencies of mDCs in the blood of HIV-infected patients were decreased while expression of CCR7 was up-regulated. Up to 94.4% and 91.8% of mDCs from HIV-infected patients expressed active caspases 8 and 9, respectively, compared to 24.5% and 19.9% from healthy controls (both p<0.0001). However, monocytes and B and T cells from HIV-infected patients also showed increased levels of these caspases. Percentages of FASL expression in mDCs were also elevated in HIV-infected individuals while mDCs expressing cell membrane death receptors remained unchanged. No differences between HAART and naive patients were observed for any of the molecules measured. Caspases 8 and 9 were up-regulated in mDCs cocultured with HIV despite lack of productive infection in vitro. Extrinsic and intrinsic pathways of apoptosis are up-regulated in HIV infection but do not correlate with cell depletion.
Introduction
P
In HIV-1 infection there is progressive loss of both pDCs and mDCs 4 –13 and given the pivotal role played by DCs in the initiation of immune responses this may contribute to the immunosuppression that characterizes HIV infection. Loss of mDCs in particular may impair the generation of adaptive immune responses. Analysis of blood mDC from HIV-1-infected patients by polymerase chain reaction (PCR) for proviral DNA indicates that less than 1% are infected suggesting that infection may not be directly responsible for their reduced numbers. 7 Our previous studies showed poor survival of in vitro cultured mDCs isolated from HIV-infected individuals compared to cells from healthy controls 14 and we speculate that increased apoptosis may play a role in the loss of mDCs. In support of this hypothesis studies on in vitro-generated monocyte-derived DCs exposed to HIV found reduced levels of Bcl-2 and increased susceptibility to TRAIL and Fas ligand-induced apoptosis 15 while loss of mDCs in SIV-infected macaques was associated with caspase 8-dependent apoptosis. 16 In HIV-infected individuals there are reduced levels of Bcl-2 and increased expression of Annexin V, 7AAD, and caspase 3 in mDCs, further supporting apoptosis as a mechanism of mDC loss. 17,18 These findings prompted us to further analyze pathways that may be involved in mDC apoptosis in HIV infection.
There are two major apoptotic pathways, the extrinsic pathway that is initiated by receiving signals through death receptors on the cell surface, and the intrinsic pathway mediated by disruption of intracellular homeostasis, a process critically dependent on mitochondria. 19,20 Apoptosis is executed by a family of cysteine-dependent aspartate-directed proteases, caspases. Caspase 8 and caspase 9 have been recognized as the initiator caspases of extrinsic and intrinsic apoptosis pathways, respectively. To provide further evidence of the pathway mediating mDC apoptosis in HIV infection we have measured the expression of active caspases 8 and 9 in mDCs from HIV-infected patients and from healthy controls. In addition, mDCs from healthy controls were treated in vitro with infectious HIV and levels of caspases 8 and 9 were measured. However, we find that expression of caspases 8 and 9 are increased in HIV infection in a number of different mononuclear cell populations and is not correlated with loss of specific cell types.
Materials and Methods
Patient characteristics
Thirty milliliters of blood in heparin tubes was collected from 10 healthy volunteers and 14 HIV-1-infected individuals attending the clinic at Chelsea and Westminster Hospital, London, UK. Ethical approval was given by the local Ethical Committee and informed written consent was obtained from each volunteer. The majority of patients were male (92.9%, 13/14) with a median age of 50 [interquartile range (IQR): 46–51] years. Nine patients were receiving highly active antiretroviral therapy with undetectable plasma viral loads and a median CD4 count of 566±244 cells/μl. The remaining five patients were therapy-naive with a median plasma viremia of 3.38±1.31 log10 and a median CD4 count of 720±331 cells/μl. There were no significant differences between therapy-naive and therapy-treated patients in terms of their CD4 T cell count (p=0.32, Student's t-test).
Blood separation
Peripheral blood monocular cells (PBMCs) were isolated within 1 h of blood collection by centrifugation over a Ficoll-Histopaque gradient (PAA Laboratories, UK). Cells were harvested, washed with phosphate-buffered saline (PBS), and live cells identified by Trypan blue (Sigma Aldrich, UK) exclusion counted.
Isolation and culture of myeloid DC from healthy individuals
Leukocyte cones were obtained from the National Blood Transfusion Unit (Colindale, London, UK). PBMCs were isolated by centrifugation over Ficoll-Histopaque and then separated into a low-density fraction enriched in DC by centrifuging over a 50% Percoll gradient for 30 min at 300×g. mDCs were then isolated using anti-CD1c immunomagnetic beads (BDCA-1; Miltenyi Biotec, Germany) according to the manufacturer's guidelines. Purified mDCs were cultured in 96-well round-bottom microtiter plates (105 cells/well) in 200 μl of RPMI 1640, containing 10% heat-inactivated endotoxin-tested fetal calf serum (FCS), 100 IU/ml penicillin, 0.1 mg/ml streptomycin, 2 mM
Infection of myeloid DCs with HIV-1
HIV-1BAL was grown in PM1 cells (both supplied by the Centralised Facility for AIDS Reagents at the National Institute for Biological Standards and Control, UK) and HIV-1 p24 gag protein in infected cultures measured by ELISA (Perkin Elmer, UK). Virus infection of mDCs was conducted as previously described by Steinman and colleagues. 21 In brief, HIV-1BAL was used to infect freshly isolated mDCs for 5 days at a dose of 2 ng of p24 gag/105 mDCs. Infection of DCs by HIV-1 was then assessed by intracellular staining using anti-p24 antibody (KC57-PE, Beckman Coulter) and 0.5% saponin (Sigma Aldrich, UK) in PBS (PAA Laboratories, UK).
Flow cytometry and caspase detection
PBMCs or cultured mDCs were harvested and washed in PBS (PAA Laboratories) containing 2 mM ethylenediaminetetraacetic acid (EDTA), 2% FCS, and 0.05% sodium azide (NaN3, all from Sigma Aldrich, UK). Cells were stained for 20 min at 4°C with the following antibodies, which were all purchased from BD Biosciences unless otherwise stated: Percp-cy5-labeled CD3, APC-cy7-labeled CD8, PE-cy7-labeled CD14, Horizon V500-labeled CD19, FITC-labeled FAS, PE-labeled TRAIL, Brilliant Violet-labeled CD1c (Biolegend), APC-labeled DR4 (Biolegend), PE-labeled FASL (Biolegend), FITC-labeled DR5 (Abcam, UK), and PE-labeled CCR7 (R&D Systems). For some experiments, cells were further stained to detect caspase 8 and 9 activity using the CaspaTag Caspase 8 In Situ Assay Kit and CaspaTag Caspase 9 In Situ Assay Kit, respectively (Millipore, UK), in accordance with the manufacturer's instructions. Cells were then washed and fixed with BD stabilizing fixative (BD Biosciences). Cells were acquired using a three-laser configuration LSR-II machine (BD Biosciences). The data were then analyzed using FlowJo 7.6.4 (Tree Star, Inc., USA). Between 4×10E5 and 7×10E5 live events were acquired that corresponded to a minimum of 1,000 events within the mDC gate. The gating strategy utilized to identify the different populations is shown in Supplementary Fig. S1 (Supplementary Data are available online at
Detection of caspase 3 and Annexin V
To correlate caspase 8 and 9 activity with a proapoptotic phenotype, active caspase 3 and Annexin V expression were measured in proof of concept experiments. In brief, PBMCs from six healthy individuals were left untreated or heated at 56°C for 5 min. Cells were then stained for caspase 8 and 9 as previously indicated followed by staining with APC-conjugated Annexin V (BD Biosciences). Cells were fixed with 2% paraformaldehyde in PBS for 10 min followed by permeabilization using 0.5% saponin in PBS and stained with PE-conjugated active-caspase 3 Ab (BD Biosciences). Cells were washed and fixed with BD stabilizing fixative, acquired, and analyzed using FlowJo 7.6.4.
Statistical analysis
Normality of the data was assessed using a Shapiro–Wilk test. Normally distributed data are summarized as mean±standard deviation while nonnormally distributed data are shown as median (interquartile range). Student's t-test analysis was performed to assess the differences between two groups. Wilcoxon rank-sum tests were used for nonnormally distributed data. Matched samples were analyzed using paired Student's t-test. Analyses were two-sided and p values below 0.05 were considered significant. All statistical analyses were performed using GraphPad Prism version 5.0 software (GraphPad Software, San Diego, CA).
Results
The percentages of mDCs in peripheral blood were decreased in HIV-infected patients
The percentage of mDCs was found to be significantly lower in HIV-1-infected patients compared to healthy controls [0.61% (0.31–0.95%) vs. 1.10% (0.74–1.41%) respectively, p<0.05, Fig. 1a]. mDCs in therapy-treated patients accounted for 0.69%±0.29% of total PBMCs while mDCs in naive patients represented 0.52%±0.32% of total PBMCs. It has previously been reported that blood DCs in HIV patients may be depleted from the periphery as they migrate to secondary lymphoid tissues. 18,22 Hence we analyzed the expression of CCR7, a lymphoid homing marker, on circulating mDCs in both healthy and HIV-1-infected patients. The percentages of mDCs expressing CCR7 on their surface in both HIV seropositives and healthy controls were minimal (1.12%±0.90% vs. 1.26%±0.75% for healthy controls and HIV-infected patients, respectively, p=0.69). However, the mean fluorescence intensity (MFI) of CCR7 was found to be statistically higher in the small number of cells that expressed CCR7 in mDCs from HIV-infected patients compared to healthy controls (911.1±414.2 vs. 334.8±159.9, respectively, p<0.001, Fig. 1b).
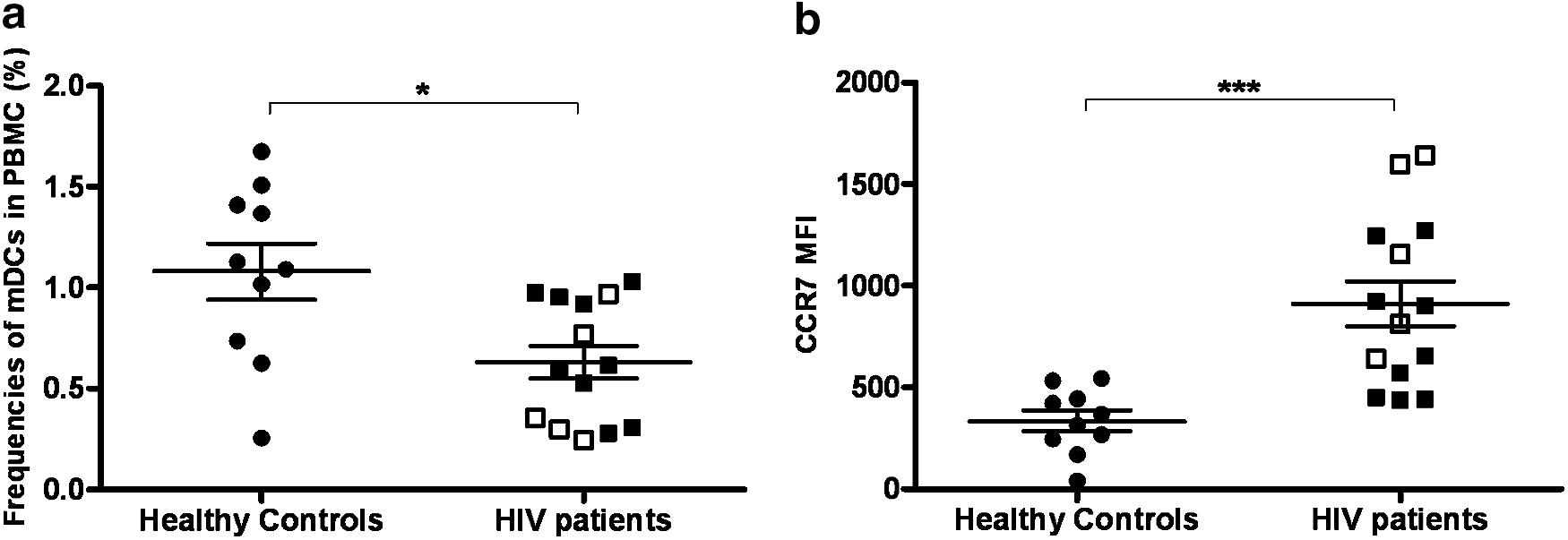
Frequency of myeloid dendritic cells (mDCs) in peripheral blood was decreased in HIV-infected patients while CCR7 expression on their surface was up-regulated. Frequencies of mDCs in peripheral blood mononuclear cells (PBMCs) were decreased in HIV-infected patients (n=14) compared to healthy controls (n=10)
Caspases as indicators of apoptosis
As proof of principal that caspase 8 and 9 activity is associated with apoptosis PBMCs were heated to 56°C for 5 min to induce apoptosis and then assayed for Annexin V and for caspase activity. The data from these experiments are summarized in Supplementary Fig. S2. Untreated PBMCs showed minimal expression of active caspase 8 and 9, while they were induced in heat-treated PBMCs (Supplementary Fig. S2A). The majority of caspase 8 or 9-negative PBMCs did not express active caspase 3 or Annexin V. Cells that expressed high levels of caspase 8 or 9 showed significant expression of active caspase 3 and moderate expression of Annexin V indicating their apoptotic status (Supplementary Fig. S2B). Cells that expressed low levels of caspase 8 or 9 did not express active caspase 3 or Annexin V, which suggests that activation of caspase 8/9 may precede activation of caspase 3 and expression of Annexin V (data not shown).
mDCs in HIV-infected patients exhibited high levels of active caspase 8 and 9
Extremely high proportions of mDCs [94.4% (80.5–96.2%)] within PBMCs of HIV-infected patients expressed active caspase 8, which was significantly higher than that in healthy controls [24.5% (17.1–49.7%), p<0.0001, Fig. 2a]. Similarly, 91.8% (83.4–94.7%) and 19.9% (13.6–26.3%) of mDCs expressed the active form of caspase 9 in HIV-1-infected individuals and healthy controls, respectively (p<0.0001, Fig. 2b). To determine whether caspase 8 and 9 were exclusively elevated in mDCs, we analyzed their expression by other cell types including B cells, CD4 and CD8 T cells, and monocytes. The proportions of B cells, CD4 and CD8 T cells, and monocytes expressing active caspase 8 and 9 were also found to be significantly higher in HIV-infected patients compared to healthy controls (Fig. 2a and b). Among the HIV-infected patient cohort, the frequencies of mDCs expressing caspase 8 and caspase 9 were comparable between patients who received antiretroviral treatment and those who did not, though no statistical analysis was performed to corroborate this observation due to the low number of therapy-naive patients.
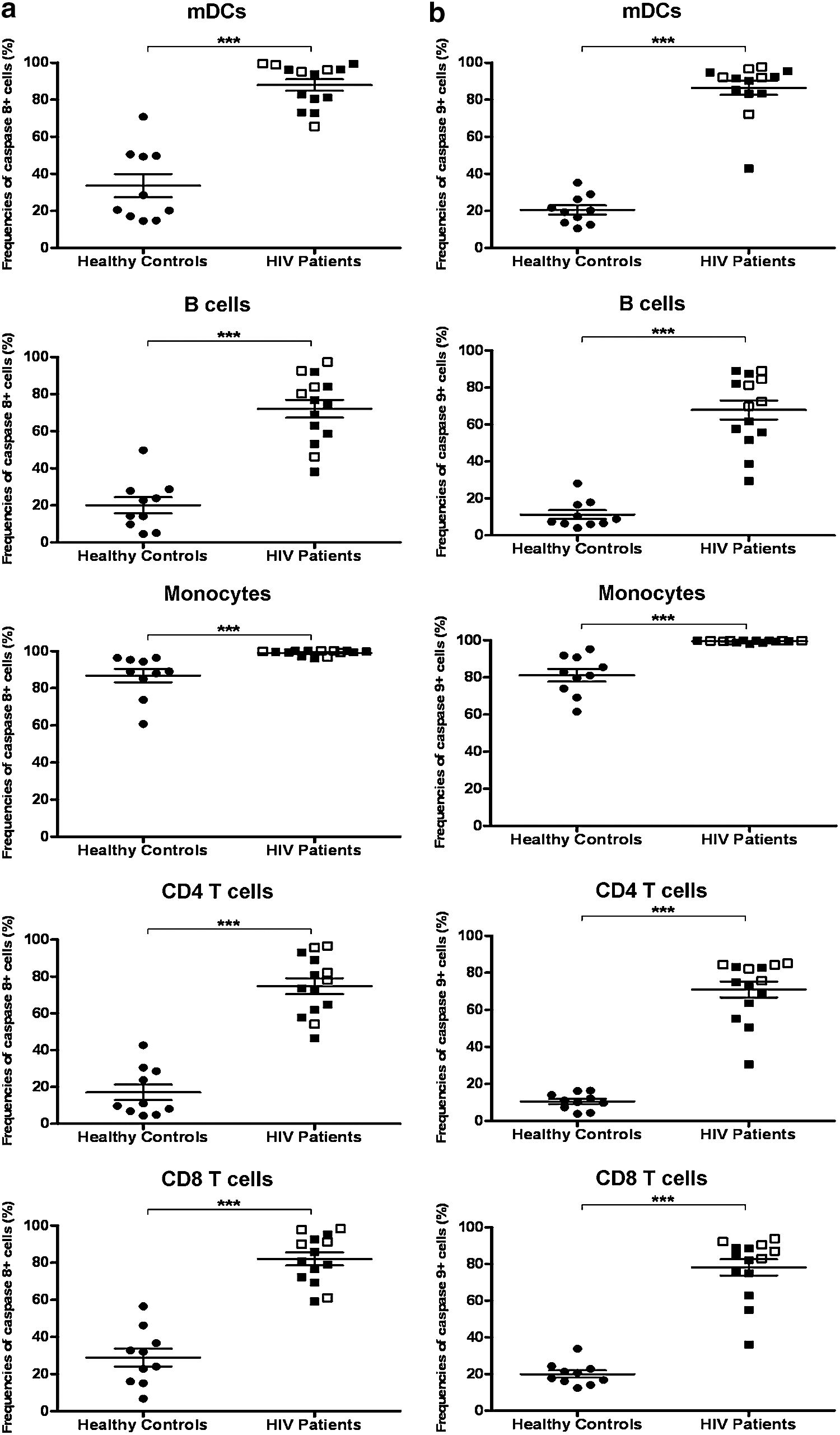
Cells from HIV-infected patients displayed a proapoptosis profile. Frequencies of cells that expressed active caspase 8
mDCs from HIV-infected patients expressed normal levels of death-inducing receptors
We observed increased levels of active caspase 8 expression by mDC in HIV-1-infected individuals, indicating that the extrinsic pathways of apoptosis were activated. Therefore, we sought to determine whether death-inducing receptors were up-regulated in these cells. The proportions of mDC, monocytes, and B cells from HIV-infected patients were found to express similar levels of FAS, DR4, and DR5 in comparison to healthy controls (Fig. 3a, b, and c). However, the frequencies of CD4 and CD8 T cells expressing FAS from HIV-infected patients were statistically higher than that in healthy controls (HIV-infected patients vs. healthy controls: 58.3%±12.8% vs. 40.0%±5.4%, p<0.0001 and 76.3% (54.3–80.6%) vs. 50% (39.9–61%), p<0.05, for CD4 T cells and CD8 T cells, respectively, Fig. 3a). In terms of TNF family death ligands, mDC, CD4, and CD8 T cells, monocytes, and B cells in HIV-infected patients all expressed significantly higher percentages of FASL but not TRAIL (Fig. 4a and b). There were no apparent differences in the expression of these death receptors and death ligands between highly active antiretroviral therapy (HAART) patients and naive patients (Fig. 4a and b), though no statistical analysis was performed to determine the significance between the above two groups due to the low numbers of therapy-naive patients.
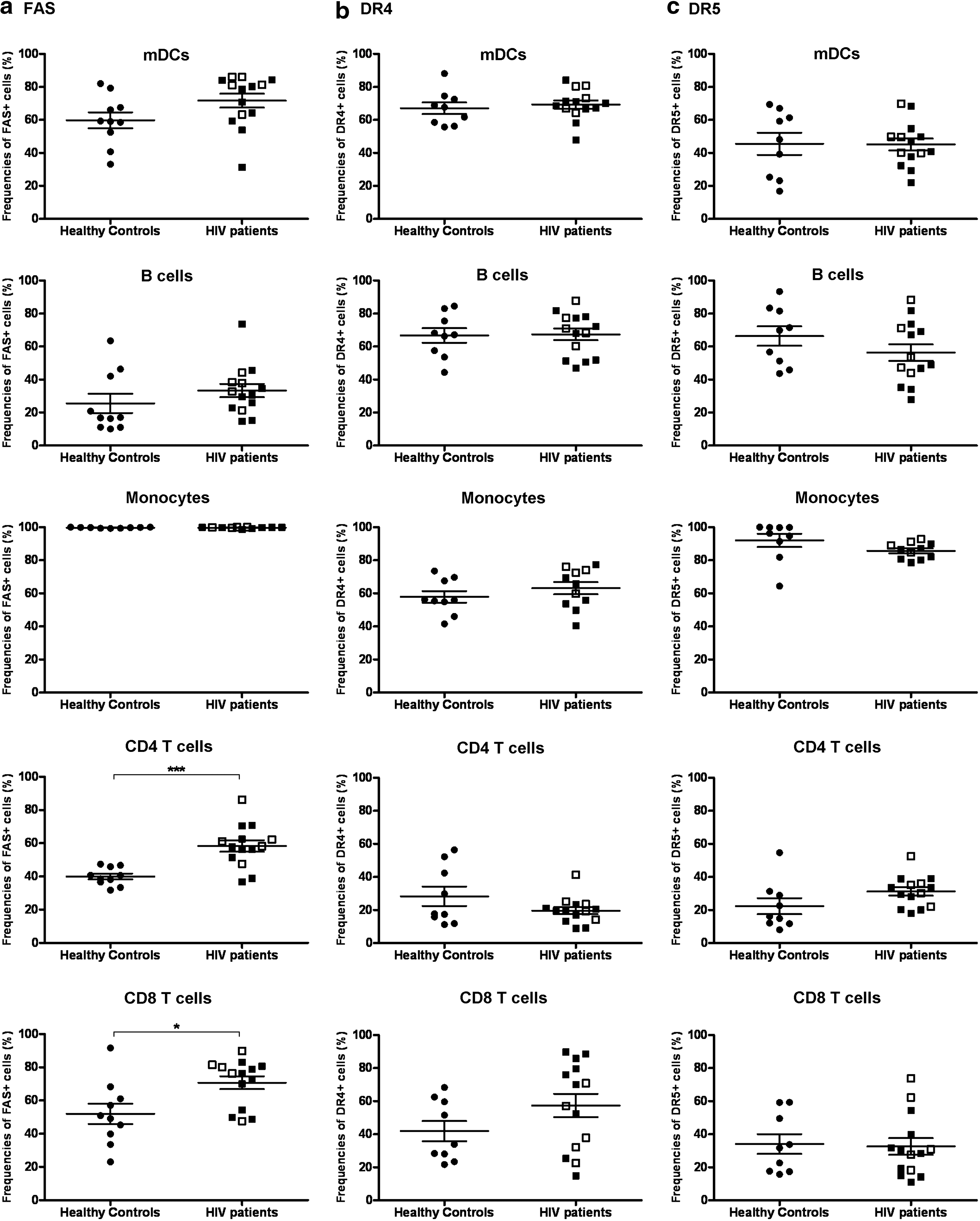
mDCs from HIV-infected patients expressed normal levels of death receptors on their surfaces. The expression of FAS
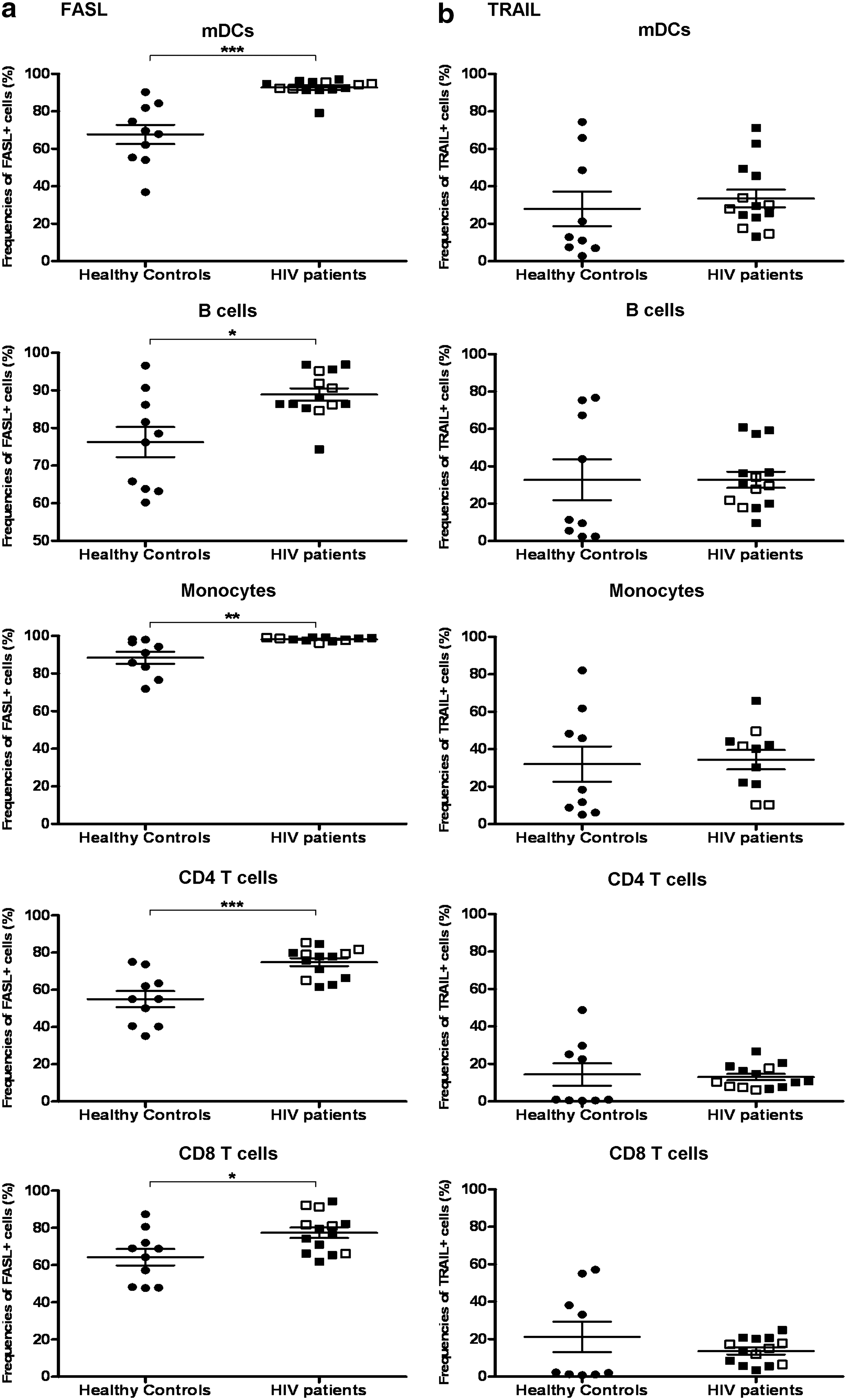
The frequencies of cells that express death receptor ligands on their surface were increased in HIV-infected patients. Percentages of FASL+ cells
Caspase 8 and caspase 9 expression were up-regulated in mDCs cocultured with HIV despite the absence of productive infection in vitro
Since we observed increased caspase activity and elevated death ligand expression in mDCs from HIV-1-infected individuals, we addressed the possibility that it may be a direct effect of HIV-1 infection of these cells. Thus, we purified mDCs from healthy individuals and treated them with infectious HIV-1 bal for a period of 5 days as previously described by Granelli-Piperno et al. 21 Negligible percentages of mDCs were found to be positive for HIV-1 p24 as indicated by intracellular staining (Fig. 5a). The proportions of mDCs that were found to be positive for active caspase 8 and 9 were comparable between HIV-1-treated and untreated mDCs [0.72% (0.22–1.19%) vs. 0.61% (0.40–1.13%), p=0.96 and 0.61% (0.23–1.31%) vs. 0.59% (0.31–0.93%), p=0.73 for caspase 8 and caspase 9, respectively, Fig. 5b]. However, the MFIs of both caspase 8 and caspase 9 in the cells that expressed these caspases were up-regulated in HIV-1-treated mDCs compared to the control group (5430.1±569.5 vs. 4318.8±430.3, p<0.05 and 5303.6±636.6 vs. 3820.4±339.1, p<0.05 for caspase 8 and caspase 9 in HIV-treated vs. untreated mDC, respectively, Fig. 5c). We attributed this difference to HIV-1 virions as treatment of mDCs with supernatant from the uninfected PM1 cell line that was used to grow HIV had no effect on caspase 8 or 9 expression (data not shown). There were no statistically significant differences between the expression of FAS, DR5, FASL, and TRAIL on the surfaces of mDCs when cocultured with HIV and control medium (data not shown).
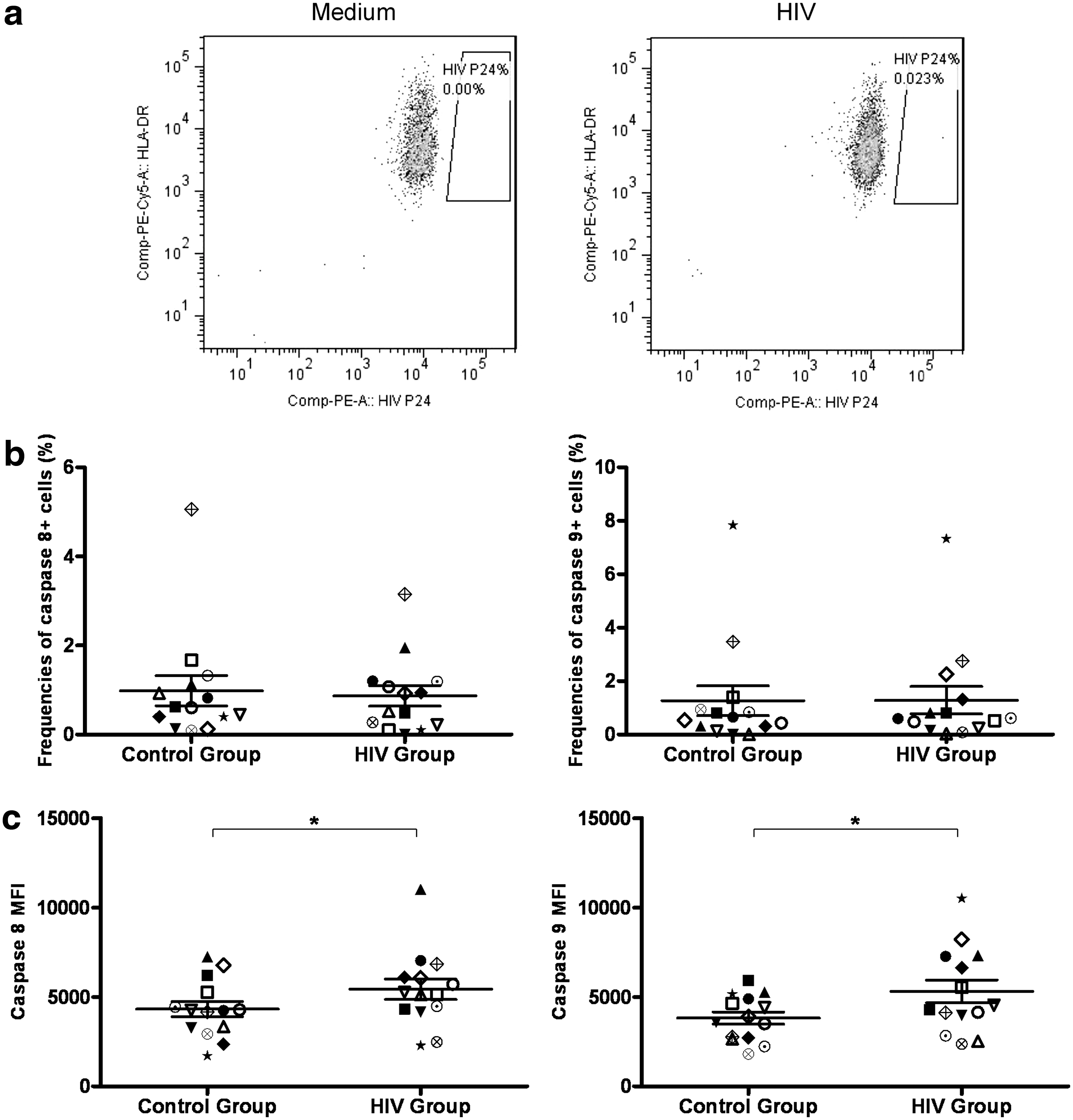
mDCs were more likely to undergo apoptosis in the presence of HIV in vitro despite unsuccessful infection. mDCs from healthy controls were treated with HIV-1BAL for 5 days and p24 levels were measured by intracellular staining
Discussion
The progressive loss of mDCs in HIV-1 infection has been reported by a number a groups 4,8,11 –13 and increased apoptosis is reported as a potential mechanism of loss. 14 –17 To provide further information on the detailed mechanism of induction of apoptosis in mDCs we analyzed expression of caspases 8 and 9, which are associated with early stages of the extrinsic and intrinsic pathways of apoptosis in mDCs from HIV-infected patients. Increased levels of both caspase 8 and 9 were observed in patients' mDCs compared to controls but interestingly there were no striking differences between antiretroviral-naive and HAART patients. This is in agreement with a previous study showing that HAART failed to restore Bcl-2 expression level in mDCs in HIV patients. 17 As HAART patients had undetectable virus loads, it suggests that enhanced activity of caspases 8 and 9 was not directly stimulated by virus. Furthermore, in view of reports that HAART can improve mDC numbers, increased expression of caspases 8 and 9 does not appear to be correlated with cell loss. This is supported by the observation in this report that even for cells that are not depleted, namely B cells and CD8 T cells, there is increased expression of caspases 8 and 9 in HIV-infected patients. We also examined levels of death receptor expression on HIV patient mDCs but this was found to be no different from expression levels in controls. These findings suggest that although raised levels of caspases 8 and 9 may predispose mDCs to apoptosis a further trigger is required for full activation of the pathway.
We were surprised to find that all mononuclear cells examined from HIV-infected patients showed increased levels of caspases 8 and 9 compared with healthy controls. Since not all mononuclear cells are depleted during HIV-1 infection, expression of caspase 8 and 9 does not necessarily seem to correlate with cell depletion or later stages of apoptosis. Our observations suggests that HIV infection, perhaps through dysregulation of cytokines, causes an increase in caspases 8 and 9 associated with early stages of extrinsic and intrinsic pathways of apoptosis in a number of different blood leukocytes, which may cause predisposition to apoptosis. HIV itself may also induce higher expression of caspases 8 and 9 since the virus was found to increase their expression in isolated mDCs; however, this observation should be interpreted bearing in mind that a very low percentage of cells had detectable caspase activity. In fact, the levels of caspase 8 and 9 activity in cultured mDCs were negligible in comparison to those observed in mDCs within freshly isolated PBMCs even in the absence of HIV-1 infection. It is possible that in vitro culture of mDCs favors the survival of cells with low caspase 8 and 9 activity. Nonetheless, the fact that treatment of mDCs with HIV-1 virions in vitro resulted in higher caspase 8 and 9 activity supports the observation that HIV infection in vivo may prime several different cell types for apoptosis. However, additional signals that only DC and CD4 T cells are susceptible to are required for full activation of the apoptotic pathway leading to their depletion. A recent study showed that the Bcl-2 levels in mDCs from HIV patients were unchanged while caspase 3 levels were decreased after HAART, indicating a role of HIV in stimulating these additional signals for the apoptotic pathway. 17
Other mechanisms may contribute to the loss of mDCs in HIV infection. Increased accumulation of mDCs in lymph nodes in chronic infection 11 and increased migratory properties of mDCs in AIDS 18 may contribute to the depletion of blood mDCs. A recent study also reported that chemokine levels were elevated, although expression of chemokine receptors on mDCs was normal in HIV-infected patients with a stable response to HAART. 23 In the present study, CCR7 expression on mDCs was up-regulated in HIV-infected patients. Thus, mDC migration could be one of the causes of mDC depletion in circulation. However, the percentages of mDCs expressing CCR7 were extremely low, which suggests that migration of mDCs to lymphoid nodes might not be the main reason for their depletion. Evidence for destruction of mDCs is provided by data from rhesus macaques with AIDS showing mDC depletion in lymph nodes. 18,24
In conclusion, our study found an increase in caspases 8 and 9 in cells that are maintained and depleted in HIV infection. This may indicate that there is a broad priming of different cells types for apoptosis in HIV infection but that further stimuli are required, perhaps provided by the virus, to induce full apoptosis.
Footnotes
Acknowledgments
This research was supported by EUROPRISE (contract number 037611).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.