
Abstract
Bacterial vaginosis has been associated with genital HIV-1 shedding; however, the effect of specific vaginal bacterial species has not been assessed. We tested cervicovaginal lavage from HIV-1-seropositive women for common Lactobacillus species: L. crispatus, L. jensenii, and seven BV-associated species: BVAB1, BVAB2, BVAB3, Leptotrichia, Sneathia, Megasphaera, and Atopobium spp. using quantitative PCR. We used linear and Poisson regression to evaluate associations between vaginal bacteria and genital HIV-1 RNA and DNA. Specimens from 54 U.S. (310 visits) and 50 Kenyan women (137 visits) were evaluated. Controlling for plasma viral load, U.S. and Kenyan women had similar rates of HIV-1 RNA (19% of visits vs. 24%; IRR=0.95; 95% CI 0.61, 1.49) and DNA shedding (79% vs. 76%; IRR=0.90; 0.78, 1.05). At visits during antiretroviral therapy (ART), the likelihood of detection of HIV-1 RNA shedding was greater with BVAB3 (IRR=3.16; 95% CI 1.36, 7.32), Leptotrichia, or Sneathia (IRR=2.13; 1.02, 4.72), and less with L. jensenii (IRR=0.39; 0.18, 0.84). At visits without ART, only L. crispatus was associated with a lower likelihood of HIV-1 RNA detection (IRR=0.6; 0.40, 0.91). Vaginal Lactobacillus species were associated with lower risk of genital HIV-1 shedding, while the presence of certain BV-associated species may increase that risk.
Introduction
U
Materials and Methods
Study cohort
Data were collected during prospective, observational studies of HIV-1-infected women in Seattle, WA (n=35), Rochester, NY (n=17), and Nairobi, Kenya (n=76) between 2002 and 2009. Study participants were 18–50 years of age at study entry, were not pregnant or menopausal, had an intact cervix, and had no active genital infections at study entry. There were no entry restrictions with regard to CD4 cell count, PVL, or antiretroviral therapy (ART). The University of Washington, University of Rochester, and University of Nairobi Institutional Review Boards approved the studies, and all participants provided written informed consent.
Participants enrolled in the United States had four study visits in the first year and three visits per year in subsequent years. Participants enrolled in Kenya were followed every 3 months for 1 year. Each study visit included a comprehensive face-to-face interview ascertaining demographic information, reproductive and medical history, sexual behavior, and medication use. Plasma was obtained for HIV RNA quantification. A pelvic examination was performed during which cervicovaginal lavage (CVL) was performed by lavaging the ectocervix and vaginal walls with 7 ml of a solution of phosphate-buffered saline and 10 mM lithium chloride (LiCl), which was then collected from the vaginal fornix, spun at 3000 rpm for 5 min to separate the epithelial cells, and stored at −80°C in 1-ml aliquots. A sample from the cervical os was collected by twirling a cytobrush 360 degrees in the os. This was then placed in a cryovial and stored at −80°C.
Plasma and CVL HIV-1 RNA were quantified by an independently validated real-time polymerase chain reaction (PCR) assay described previously 7 with a lower limit of quantification of 30 copies/ml. Cervical cytobrush specimens underwent silica extraction, 8 followed by real-time PCR amplification of the human beta-globin gene to quantify the number of cells collected. 9 A portion of HIV-1 gag was amplified to quantify the amount of viral DNA as previously described. 10 Samples were considered inadequate if fewer than 100,000 cells were present in the cytobrush sample.
Bacterial vaginosis (BV) was diagnosed from vaginal Gram stain using Nugent's criteria 11 ; a score of 0–3 was considered normal, 4–6 intermediate, and 7–10 positive for BV. Yeast was diagnosed by wet mount. Cervicitis was defined as more than 30 white blood cells seen per high-powered (100 ×) microscopy field on the Gram-stained slides. Trichomonas vaginalis was detected by culture using the InPouch system (Hardy Diagnostics, Santa Maria, CA) in the United States, and by wet mount in Kenya. Urine was collected for Neisseria gonorrhoeae and Chlamydia trachomatis detection by nucleic acid amplification (COBAS Amplicor PCR).
A 150-μl aliquot of CVL underwent DNA extraction and PCR for human papillomavirus (HPV) as previously described 12 using MY09/MY11 primers, and then underwent subtyping by reverse line blot assay. The remaining CVL was spun at 3,000 rpm for 5 min to pellet any cellular debris and supernatant was tested for genital shedding of herpes simplex virus (HSV) type 1 and 2, and cytomegalovirus (CMV) using PCR. 13,14 The CVL cell pellet was resuspended in 350 μl of lysis buffer and DNA was extracted using the MoBio Bacteremia Kit (Mo Bio Laboratories Inc., Carlsbad, CA), as previously described. 15 Extracted DNA was tested in a quantitative PCR assay using primers targeting the human 18S rRNA gene to validate that successful DNA extraction occurred. An internal noncompetitive amplification control PCR using exogenous DNA from a jellyfish gene was used to test for the presence of PCR inhibitors. 15
DNA extracted from the CVL cell pellet was then subjected to taxon-directed TaqMan 16S rRNA gene quantitative PCR assays for the detection and quantification of L. crispatus, L. jensenii, Atopobium vaginae, Megasphaera spp., Leptotrichia/Sneathia spp., BVAB1, BVAB2, and BVAB3, which have been described elsewhere. 15,16 The assays were run using an ABI 7500 Thermocycler (Applied Biosystems, Foster City, CA) or Eppendorf Mastercycler ep Realplex thermal cycler (Eppendorf, Westbury, NY). Negative reactions were assigned a value at the lower limit of detection for that bacterium for quantitative analyses.
Linear regression using generalized estimating equations (GEE) with an independent correlation structure and robust standard errors were used to assess the effect of each bacterial species on detection or log10 quantity of genital HIV-1 RNA and DNA. This method accounts for correlation due to repeat measures among subjects. Given that detection of HIV-1 RNA and of DNA were common outcomes (>10%), we used Poisson regression with GEE (as described above) to assess the effect of bacterial species on detection of HIV-1 RNA and DNA. Test results for genital infections (HSV, CMV, HPV, gonorrhea, chlamydia, and cervicitis) were missing for 5–10% of specimens. These data appeared to be missing at random, therefore multiple imputation using chained equations (ICE) was performed. 17 Potential confounders were evaluated using the imputed dataset and multivariable models were constructed to control for plasma viral load (chosen a priori), as well as variables associated with the presence of HIV-1 RNA or DNA shedding in univariable analyses (p<0.10). We evaluated for effect modification by use of ART in separate models that included variables for the outcome, species of bacteria, log10 PVL, ART use. and an interaction term for the species and ART use. All statistical tests were assessed using a two-sided alpha of 0.05 and were conducted using Stata version 11.0 (StataCorp, Inc., College Station, TX).
Results
A total of 108 HIV-1-infected women were enrolled, 58 in the United States and 50 in Kenya, contributing specimens and clinical data from 566 visits. Visits without plasma or genital viral load results or information on ART use were excluded, leaving 104 women contributing 447 visits for this analysis, 310 (69%) from U.S. women and 137 (31%) from Kenyan women. The mean number of visits±standard deviation for U.S. women was 5.7±3.6, with a range from 1 to 14, while Kenyan women averaged 2.7±1 visits, with a range from 1 to 4. U.S. women were slightly older than Kenyan women (Table 1), more likely to use ART, and to have had a tubal ligation.
IQR, interquartile range; IUD, intrauterine device.
U.S. women reported use of ART at 176 (57%) visits, and Kenyan women at 11 (8%) visits. HIV-1 RNA was detected in cervicovaginal lavage at 77 (30%) visits when subjects were not taking ART and at 14 (7%) of visits while subjects were on ART (p=0.005). HIV-1 DNA was detected in cervical cytobrush samples at 197 (76%) visits when subjects were taking ART and 153 (82%) visits when they were not (p=0.29). Grouping across ART status, U.S. women had HIV-1 RNA detected at 58 (19%) visits and Kenyan women at 33 (24%) visits (IRR 0.95; 95% CI 0.61, 1.49), while HIV-1 DNA was detected at 79% of visits in U.S. women and 76% in Kenyan women (IRR 0.90; 0.78, 1.05) Of note, five women from the United States and five women from Kenya never shed HIV-1 RNA or DNA at any visit; 38% of visits from these women occurred while on ART. When detectable virus was present, the mean quantity of HIV-1 RNA and DNA detected was similar between U.S. and Kenyan women (Fig. 1).
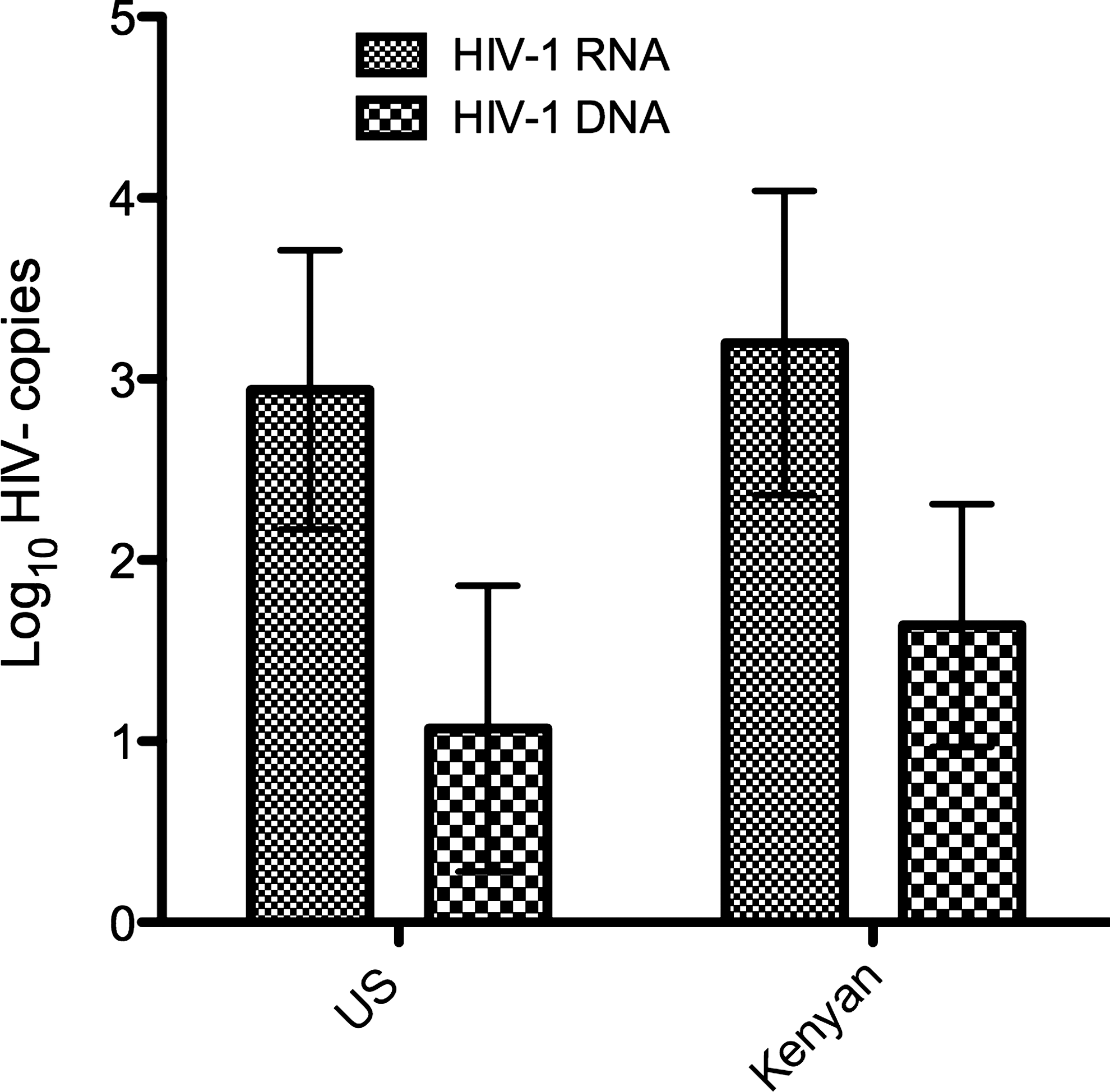
Comparison of quantity of HIV-1 RNA (copies/ml cervicovaginal lavage) and HIV-1 DNA (copies/106 cells) between U.S. and Kenyan women with detectable virus in genital tract samples. No significant differences were seen in quantity of genital tract RNA (p=0.16) or DNA (p=0.32) between U.S. and Kenyan women with virus detected.
Yeast vaginitis was more prevalent in U.S. women than Kenyan women, detected at 64/310 visits (21%) versus 14/147 (10%; p=0.02). CMV shedding in CVL was more prevalent in Kenyan women, detected at 21/137 visits (15%) versus 13/282 visits (5%) in U.S. women (p=0.005). Sexually transmitted infections were uncommon; N. gonorrhoeae was detected at 1/424 visits (<1%), C. trachomatis at 4/425 visits (<1%), and (in U.S. women only) T. vaginalis in 18/309 visits (6%). Trichomoniasis testing was performed at only 75/137 (55%) visits for Kenyan women, and no cases were found. HSV shedding was detected in CVL at 11 visits (4%) in U.S. women compared with 12 visits (9%; p=0.07) in Kenyan women. In univariable analyses, plasma HIV-1 RNA concentration (log10) (IRR=2.06; 95% CI 1.73, 2.44) and yeast vaginitis (IRR=1.70; 1.16, 2.5) were positively associated with detection of HIV-1 RNA in CVL. BV, CMV shedding, HSV-2 shedding, cervicitis, and trichomoniasis (U.S. women only) were not significantly associated with HIV-1 RNA shedding (data not shown).
Over the course of the study, the proportion of visits at which women had BV was similar between U.S. and Kenyan women (30% vs. 24%, respectively, p=0.40). However, L. jensenii was more frequently detected among U.S. women (p=0.001), and there appeared to be a trend to greater detection of BVAB1 (p=0.06) and BVAB3 (p=0.07) in U.S. women (Fig. 2). The quantity of Megasphaera was higher in U.S. women compared to Kenyan women (p=0.02), although it was detected in a similar proportion of women. At visits in which women were not taking ART, those colonized with L. crispatus were less likely to have HIV-1 RNA genital shedding than women who were not colonized with L. crispatus (after controlling for PVL and yeast) (Table 2). At visits while women were taking ART, detection of L. jensenii was associated with a lower likelihood of detection of genital HIV-1 RNA, while BVAB3, Leptotrichia, and Sneathia were associated with an increased likelihood of detection. Inclusion of geographic site in the models did not change these associations (data not shown).
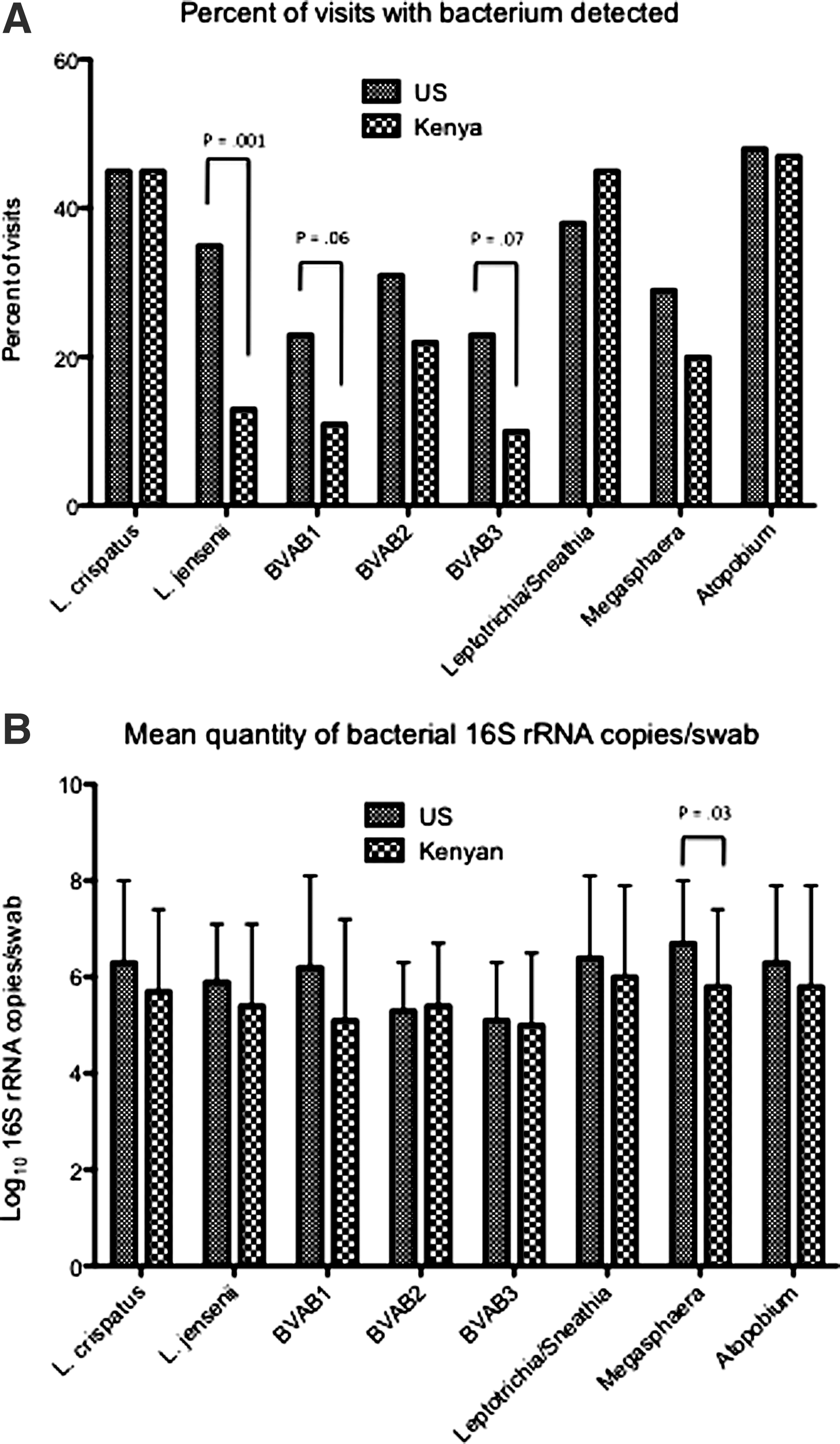
Comparison of vaginal bacterial colonization between U.S. and Kenyan women. Rates
Models adjusted for plasma viral load and the presence of yeast. Referent group=visits where HIV-1 RNA was not detected.
Number and proportion of visits where species was detected.
We evaluated effect modification by antiretroviral therapy (ART) use in separate partially adjusted models that included variables for the outcome, the species, log plasma viral load (PVL), ARV use, and an interaction term for the species and ART use. We have indicated statistical significance of the interaction term at the following levels: * p≤0.05; ** p≤0.01; *** p≤0.001.
Plasma HIV-1 RNA concentration (IRR=1.09; 1.04, 1.14), ART use (IRR=1.33; 1.16, 1.54), and yeast (IRR=1.14; 1.00, 1.31) were also associated with increased detection of HIV-1 DNA in cervical cytobrush samples, while cervicitis was unexpectedly associated with a decreased likelihood of detection (IRR=0.87; 0.77, 0.99). BV, CMV shedding, HSV-2 shedding, and trichomoniasis (U.S. women only) were not associated with detection of HIV-1 DNA (data not shown). ART use did not appear to modify the association between HIV-1 DNA genital shedding and vaginal bacteria, thus we did not stratify this analysis. In a model that adjusted for ART use and the presence of yeast, no individual bacterial species was associated with the detection of HIV-1 DNA (Table 3). There was no association between the detection of specific bacterial species and quantity of HIV-1 DNA shedding (data not shown).
Models adjusted for plasma viral load and the presence of yeast. Referent group=visits where HIV-1 DNA was not detected.
Number and proportion of visits where species was detected.
Discussion
The human vagina is colonized with millions of bacteria 18 ; at times the microbial community is dominated by a few Lactobacillus species, and at times, such as in BV, it is characterized by significant diversity. 19 We undertook this investigation to assess whether the presence of individual bacterial species was associated with genital HIV-1 RNA and DNA shedding. Lactobacilli have antiinflammatory properties 20 and have been linked to decreased HIV-1 genital shedding. 21 Conversely, women with abnormal vaginal microbiota have higher levels of inflammatory cytokines 22,23 and higher rates of genital HIV-1 shedding. 21,24 We found that Lactobacillus species were associated with less detection of HIV-1 RNA in the genital tract, but found an association between genital HIV-1 RNA and BV-associated species only in women taking ART.
BV has been associated with increased genital proinflammatory cytokines, 25 and HIV-1 shedding, 21 though this association is not consistent across studies. We suspect that this variation is due to the fact that BV is a clinical syndrome that can differ in terms of microbial composition between women. 19 Different species of bacteria have been shown to have different effects: Mycoplasma hominis has been associated with higher levels, and Lactobacillus spp. with lower levels of genital HIV-1 RNA. 21 Gardnerella vaginalis has been associated with higher concentrations of vaginal interleukin (IL)-1β 26,27 and Lactobacillus spp. with lower concentrations of IL-8. In our analysis, use of ART modified the association between genital HIV-1 RNA shedding and the presence of vaginal bacteria.
Among women not using ART, we confirmed the suppressive effect of Lactobacillus species, showing that detection of L. crispatus is associated with a 35% lower risk of HIV-1 RNA shedding. However, in these women, there was no effect of any BV-associated bacterium on the risk of genital HIV-1 RNA shedding, even after controlling for plasma concentrations of HIV-1 RNA. This may be due to either ongoing replication of virus in the bloodstream, which then passes to the genital secretions via transudation, or due to production of virus from infected cells in the genital tract tissue, which is not impacted by the presence of vaginal bacteria. When viral replication is suppressed by ART, we found that a different Lactobacillus species (L. jensenii) was associated with decreased detection of genital HIV-1 RNA, and that two of the seven BV-associated species we tested for increased the likelihood of detecting HIV-1 RNA in genital secretions. This may be due to a transient stimulation of viral expression via activation of NF-κB 28 or via other inflammatory pathways. HIV-1 DNA detection in the genital tract was not affected by vaginal bacterial colonization, as we found no association between the presence of any single bacterial species and detection or quantity of HIV-1 DNA.
Our work confirms previous findings that PVL and ART significantly affect the presence and quantity of genital HIV-1 RNA and DNA shedding. 29 Systemic treatment of HIV-1 is associated with a decreased risk of transmission to sexual partners, 30 likely because of the associated decrease in prevalence and quantity of HIV-1 shedding in genital secretions. In fact, a recent study showed that the quantity of HIV-1 RNA in genital secretions directly correlates with transmission events. 3 While antiretroviral treatment reduces HIV-1 transmission, not all patients will qualify for ART, and others shed virus in genital secretions despite successful therapy. 1 Thus, understanding the range of factors that contribute to genital shedding of HIV-1 is important.
Many hypothesize that genital tract inflammation has a significant role in promoting HIV-1 shedding. 2,31 Genital infections associated with significant inflammation, including gonorrhea, chlamydia, yeast, and herpes, have been associated with increased rates of genital HIV-1 RNA 32,33 and DNA 34 shedding while treatment of these infections has been associated with a decrease in the detection of HIV-1 in genital secretions. 35,36 In this study, the only infection significantly associated with increased HIV-1 RNA shedding was yeast vaginitis. Typically, overgrowth of yeast in the vagina is associated with significant inflammation, which likely mediated HIV-1 RNA shedding either by increasing transudation of plasma virus or by activation of NF-κB to increase transcription of integrated HIV-1 proviruses. 28 Surprisingly, we found decreased detection of HIV-1 DNA in women with cervicitis. This could be due to increased apoptosis and destruction of infected cells, or to increased innate antiviral immunity in the area of inflammation. Alternatively, as cervicitis is characterized by a neutrophilic infiltrate, the proportion of T cells in cervicitis samples might be lower, decreasing the likelihood of detecting an HIV-1-infected cell. The paucity of women diagnosed with gonorrhea, chlamydia, or trichomoniasis, three other infections associated with clinical inflammation, limited our ability to evaluate their association with genital HIV-1 shedding.
Recent reports have described a significant difference in genital tract immune activation between women from the United States and Africa. 37 Others have shown differences in the composition of the normal vaginal microbiota between healthy white and African-American women, specifically related to decreased colonization with Lactobacillus species. 38 In our population, despite similar rates of BV, women enrolled in the United States were more likely to be colonized with L. jensenii, BVAB1, or BVAB3 than Kenyan women, which may suggest that BV in African women is characterized by different bacterial species than in American women. However, we saw no significant difference in rates of genital tract HIV-1 shedding between U.S. and Kenyan women in this study, after controlling for plasma viral load, suggesting the vaginal factors that mediate HIV-1 shedding are similar.
This study has potential limitations. Our Kenyan participants were enrolled at a single urban clinic and may be a self-selected population. The U.S. women were enrolled in two locations, but were older and more likely to be on ART. Although our participants contributed a large number of visits, the number of individual women is small. Also, given the large number of associations assessed in this analysis, our results should be seen as exploratory and hypothesis-generating. Both yeast vaginitis and cervicitis were defined only by laboratory testing, without regard to clinical symptoms, which may have led to overreporting of these conditions. HIV-1 RNA shedding and genital infections were relatively uncommon in this cohort, which may have limited our ability to detect an association with specific bacteria or infections. Detection of cervicovaginal HIV-1 RNA in Kenyan women, who were unlikely to be using antiretroviral medications, was approximately half as common as rates reported in other African cohorts not receiving treatment. 39,40 Our population was relatively healthy, with high CD4 counts and low rates of genital tract infection, which may account for the low rate of genital viral shedding, but may suggest that our results are not generalizable to a wider population. Additionally, we may have missed potential associations with species not tested for; however, the nine species we chose have been highly associated with bacterial vaginosis, which is the clinical syndrome we were hoping to examine.
Our study confirms that systemic factors such as plasma HIV-1 concentrations and use of ART are important determinants of HIV-1 genital shedding. We also showed that certain vaginal bacterial species may be more likely to induce genital shedding of HIV-1 RNA or DNA, especially when viral replication is suppressed by ART. Additionally, we showed that when Lactobacillus species are present, HIV-1 RNA genital shedding was uncommon, whether or not a woman was taking ART. This suggests that promoting a healthy, Lactobacillus-dominant vaginal microbiota may be an important strategy for decreasing the risk of viral shedding and potential transmission.
Footnotes
Acknowledgments
This work was supported by the National Institute of Allergy and Infectious Diseases at the National Institutes of Health (1R21AI080439-01A1; P01 HD40540). Dr. Mitchell is supported by a K08 award from NAIAD (1K08AI087969-01). Presented as a poster at Infectious Diseases Society of Obstetricians & Gynecologists annual meeting, August 10–13, 2011, Chicago, IL.
Author Disclosure Statement
Dr. Mitchell receives grant support from Merck, Sharpe and Dohme.