
Abstract
Due to the different mechanisms HIV-1 has evolved to escape from a neutralizing antibody response it has been extremely challenging to develop an effective anti-HIV-1 vaccine. The V3 region of the gp120 HIV-1 envelope glycoprotein has been considered as one of the possible targets for an anti-HIV vaccine. It is well known that the V3 region of gp120 is at least partially masked in circulating strains and becomes exposed only after CD4 binding. However, when the virus is bound to surface CD4, steric hindrance prevents effective neutralization by V3-directed antibodies. Here we have used a 27-residue CD4-mimetic peptide in combination with immune sera elicited by an optimally constrained V3 peptide to enhance neutralization of a panel of clade B viruses. We observed strong synergism between the immune sera and the CD4-mimetic in the neutralization of tier 1 and a representative tier 2 clade B virus suggesting that the constrained V3 peptide immunogen correctly mimics the V3 conformation even in tier 2 clade B viruses. This synergy should improve the potential of CD4-mimetic compounds for preexposure prophylaxis and in the treatment of HIV-1-infected patients who usually manifest high titers of V3-directed antibodies. Moreover, constrained V3 immunogens elicit immune sera that may neutralize HIV in synergy with CD4 binding site antibodies that expose V3 and the coreceptor binding site.
C
The V3 loop of the HIV-1 envelope protein gp120 was identified very early as a potential target for a vaccine. V3-specific antibodies are present in sera of practically all HIV-1-infected individuals. 3,4 High levels of V3-directed antibodies are also found in healthy individuals following gp120-based vaccination. 3 V3-directed HIV-1-neutralizing monoclonal antibodies isolated from infected patients such as the 447-52D and HGN194 neutralize tier 1 viruses very efficiently. 5,6 Some neutralization-resistant tier 2 viruses are neutralized by these two antibodies at elevated concentrations of the antibodies (4–100 μg/ml). However, to date most V3-directed antibodies have manifested limited neutralization of “tier 2” isolates, thus diminishing the potential of V3-based immunogens as candidates for an anti-HIV-1 vaccine.
It has been shown that V3 becomes exposed after gp120 binds to CD4 while forming the binding site for the HIV-1 coreceptor, either CCR5 or CXCR4. However, sera obtained after immunization with gp120 constructs that elicited enhanced V3-directed antibody response poorly neutralized resistant strains. The efficacy of this antibody response was considerably broadened and improved after preincubation of the virus with sCD4. 6 Thus, several studies have demonstrated that sCD4 can act in synergy with V3-directed antibodies to broaden the neutralization profile of such antibodies to include a larger fraction of tier 2 isolates. 6,7 Importantly, from the perspective of the present study, peptides and small organic molecules can mimic sCD4 and expose the V3 loop for neutralization. 8,9
We recently designed a synthetic constrained V3 peptide immunogen that elicited an antibody response capable of neutralizing a spectrum of clade-B tier 1 strains differing in the sequences of the V3 β-strands. 10 This neutralizing antibody response was five times stronger than that elicited by a gp120 construct with deleted V1/V2 and 30-fold stronger in comparison with the corresponding linear V3 peptide. 10 Neutralization of tier 2 viruses was not demonstrated.
While the development of an anti-HIV-1 vaccine has been hampered by the mechanisms that the virus uses to escape the antibody response, there has been considerable success in identifying antiretroviral drugs for preexposure prophylaxis. 11 CD4-mimetic compounds have been considered as preexposure prophylaxis. 8,12 –14 However, the low potencies of small organic CD4-mimetics, problems associated with administering CD4-mimetic peptides, and the potential of enhancing infection of CD4−, CCR5+, or CXCR4+ cells has thus far considerably limited enthusiasm for this approach.
Currently, vaccine candidates provide only modest and temporary protection against HIV-1. 15 Better protection was observed using microbicides. 11,16 Recent studies in macaques suggest that vaccine and microbicides may complement each other to provide better protection than when used separately. 17 In analogy, synergism between CD4-mimetic compounds and V3-directed antibodies could contribute significantly to protection when CD4-mimetics and a V3-directed vaccine are used together as preexposure prophylaxis.
CD4M33 is a 27-residue peptide based on a scorpion toxin scaffold that induces the same conformational change in gp120 as CD4. 18,19 Soluble CD4-mimetic compounds such as CD4M33 and the small organic molecule NBD-556 and improved variants of these compounds have been considered as potential anti-HIV-1 therapeutics or prophylactic agents. 8,13,19,20 Here we investigated the capacity of CD4M33 to act in synergy with polyclonal sera that contain a high level of V3-directed neutralizing antibodies raised against the optimally constrained C4-V3T303C-E322C peptide. 10 CD4M33 synergy was examined on a panel of HIV-1 clade B strains to examine whether the antibodies elicited by the constrained V3 peptide behave like 447-52D and other V3-directed antibodies elicited by HIV envelope immunogens that showed synergism with sCD4. 21 –23 Our study focused on testing the neutralization of clade B strains, as our immunogens were based on a clade B V3 sequence containing the GPGR V3 tip. Others have demonstrated that soluble CD4 can similarly expose the V3 for neutralization also in clade C strains. 6
The polyclonal rabbit sera used in this study were obtained from our previous study on constrained V3 peptides. 10 Our goal was to test whether the neutralizing responses elicited by the constrained V3 peptide immunogens are improved in the presence of a CD4 mimetic peptide, CD4M33, and whether, similar to what was found with antibodies elicited by gp120 immunogens, the antibody response elicited by the optimally constrained V3 peptide can neutralize tier 2 viruses in the presence of the CD4-mimic peptide. The answer to this question will have implications for the potential use of constrained V3 peptides in future vaccine development, and on the potential use of CD4-mimetic compounds in the treatment of HIV-1-infected patients and as preexposure prophylaxis.
Neutralization assays based on a single round of pseudovirus infection were carried out by Monogram Biosciences, Inc., South San Francisco, CA as previously described. 10,24 Titers were calculated as the reciprocal of the serum dilution conferring 50% inhibition (Table 1) in the presence or absence of CD4M33. CD4M33, i.e., TpaNLHFCQLRCKSLGLLGKCAGSBipCACV-NH2 (Tpa, thiopropionic acid; Bip, biphenylalanine), was synthesized as described previously. 18,25
Titers are given as 1/dilution, which resulted in 50% inhibition of the virus (IC50 values). Positive results, for which the IC50 was at least 3-fold higher than any inhibition of the aMLV negative control, are in bold. For the antisera alone (B961–B964; representing different rabbits) the IC50 was between a 10-fold dilution and a 182-fold dilution. In the presence of CD4M33 the dilution of the serum that resulted in the IC50 increased markedly up to a 1250-fold dilution. The ratio between the IC50 of sera alone and in combination with CD4M33 is shown in italic letters.
For each strain, neutralization was evaluated for the serum alone starting at a 1:10 serum dilution followed by consecutive 2-fold dilutions. Similarly for CD4M33, the assay started using 2 μg/ml peptide that, depending on the strain tested, was 3- to 10-fold higher than the IC50 of CD4M33. In a third neutralization experiment, the combination of the serum and CD4M33 at a constant ratio was tested, i.e., a 1:10 serum dilution with 2 μg/ml of CD4M33 was used as the starting concentration for the 2-fold serial dilution curve (Fig. 1). These three neutralization curves were used to assess the combination effect of the immune sera and CD4M33. The combination index (CI) was calculated using CalcuSyn software version 2.0 (Biosoft, Cambridge, UK) according to the model by Chou and Talalay 26 using the mutually exclusive assumption (similar modes of action). CI<1, CI=1, and CI>1 indicate synergism, an additive effect, and antagonism, respectively.
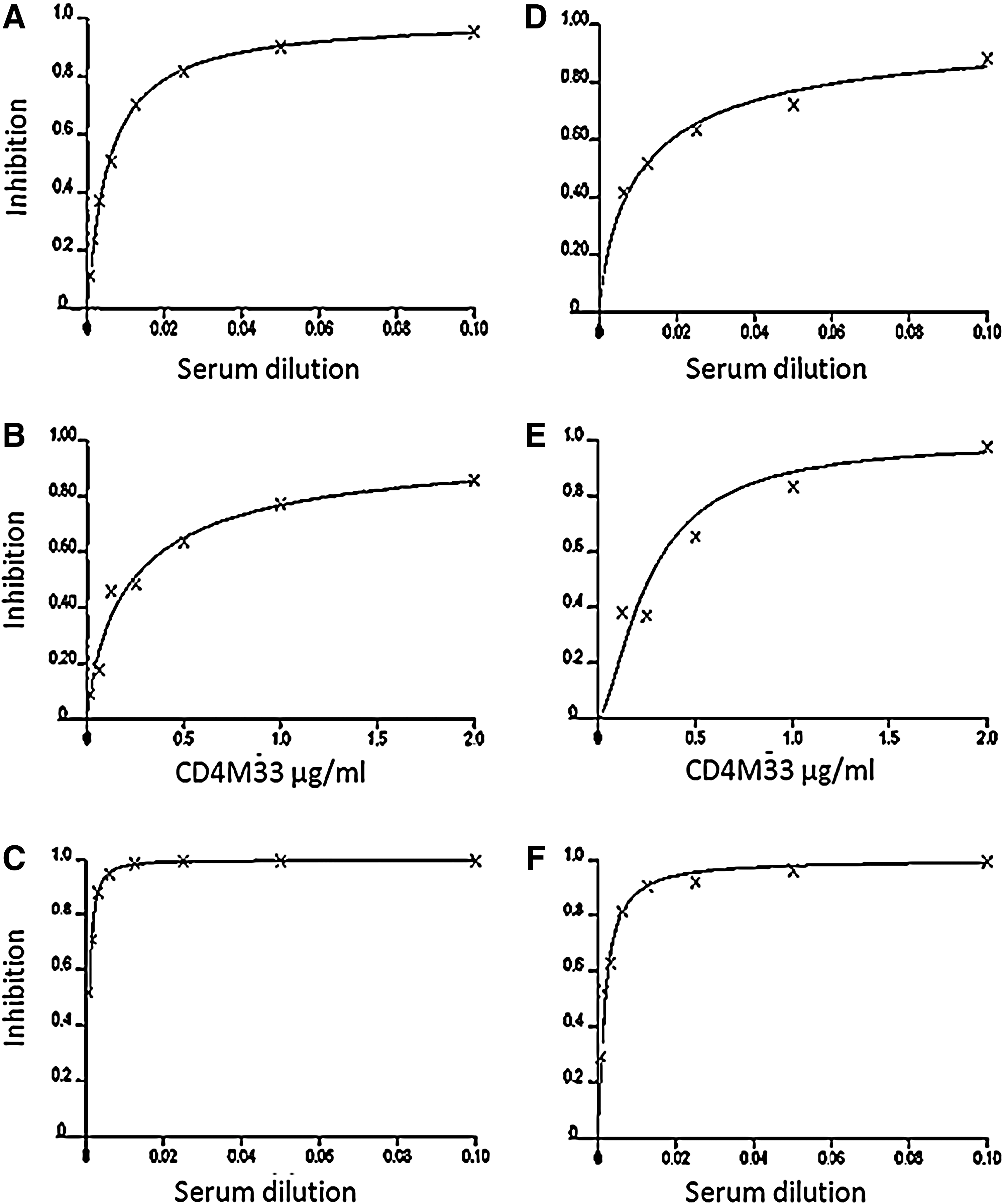
Neutralization curves for Bal
The CI for a given percent of inhibition is calculated by dividing the serum dilution (1:titer) in the combination by the serum dilution when the sera are used alone and by dividing the CD4M33 dose (in μg/ml) in the combination by the dose of CD4M33 when used alone. The CI is the sum of these two quotients. In cases where the serum alone is not neutralizing, the data presented are the ratio between the CD4M33 dose needed in combination with the sera and the dose when used alone so that values smaller than 1 indicate a synergistic effect. 1/DRI, where DRI is the dose reduction index, was used when the immune sera did not show more than 50% neutralization at 1:10 dilution. DRI determines the magnitude of the dose reduction of an agent when given in combination compared with the concentration of that agent required to achieve the same effect when given alone. A one sample t-test for a hypothetical mean value of 1 was used to evaluate whether the CI and 1/DRI values were statistically significant. GraphPad QuickCalc Internet tools (
The rabbit immune sera used in this study were raised against an optimized long constrained C4-V3 peptide, C4-V3303C-322C, containing a 19 residue V3 epitope.
10
To map the epitope recognized by the sera we determined the ELISA titer at half-maximum of the immune sera for the constrained V3 epitope used as an immunogen named longV3 (LV3) as well as the titer for another two shorter V3 peptides constrained to the same β-hairpin conformation at positions 305 and 307 named mediumV3 (MV3) and short V3 (SV3). The sequences of the peptides used are as follows: LV3: Biotin-SGS-
The CI of CD4M33 with rabbit polyclonal sera containing high levels of V3-directed neutralizing antibodies was evaluated for several clade B strains. Generally CI<0.7 is considered good synergy and CI<0.3 is considered strong synergy. Strong synergy was observed between C4-V3T303C-E322C immune sera and CD4M33 for the tier 1B viruses Bal, BX08, SS1196, and 6535 with the CI ranging from 0.2 to 0.37 for 50% inhibition and from 0.1 to 0.56 for 75% inhibition (Table 2). For the tier 2 strain QH0692, where the V3T303C-E322C immune sera were not neutralizing when used alone, a lower concentration of CD4M33 was needed to achieve a given inhibition in the presence of the sera compared to CD4M33 when used alone. The ratio of CD4M33 concentration in combination with antisera to CD4M33 concentration alone (1/DRI) ranged from 0.24 to 0.44 for 50% inhibition and from 0.43 to 0.95 for 75% inhibition of QH0692, with an average of 0.68±0.24 for the 75% inhibition. The synergy for this tier 2 strain for 75% inhibition is less pronounced than for the tier 1B stains tested, as indicated by higher 1/DRI values for 75% inhibition and a p-value of 0.074. Still 2/4 sera have CI values<0.53. Given the fact that QH0692 is one of the more resistant tier 2 viruses, 27 we feel this is an important observation. Testing more tier 2 strains is necessary to generalize our observation for QH0692.
Shown are the combination index (CI) for 50% and 75% inhibition as calculated using CalcuSyn software version 2.0.
In cases in which sera were not neutralizing shown is the ratio of the CD4M33 dose required when in combination with the sera compared to the CD4M33 when used alone. Average, standard deviation, and p-values for one sample t-test are shown.
Sterilizing immunity was achieved by the combination of sera and CD4M33 in most cases (Fig. 1). This high level of neutralization is most evident in Bal with 94–99.9% inhibition obtained at 1:80 serum dilution in the presence of 0.25 μg/ml of CD4M33. For SS1196 ∼95% inhibition was achieved at a 1:20 dilution of the serum (1 μg/ml of CD4M33); for 6535 two immune sera reached >95% inhibition at 1:20, one at 1:40 and one reached 91% inhibition at 1:10.
For BX08 92–96% inhibition was achieved at the highest concentration used, 1:10 sera +2 μg/ml CD4M33. For QH0692 85–93% inhibition was observed at the highest concentration. It is hard to predict the in vivo correlation with the inhibition results we observed for QH0692. However, given that infection by HIV-1 is known to be a relatively inefficient process, and that additional mechanisms such as ADCC may contribute to protection against the virus, 28,29 it is very possible that even a modest level of synergy can significantly improve protection.
A graphic illustration of the synergy in the form of classical isobologram plots using the mutually exclusive assumption is presented in Fig. 2. In all cases the combination point falls significantly below the diagonal, indicating synergism. When the CI values for 50% and 75% inhibition were subjected to one-sample t-test analysis all values, with the exception of 75% inhibition of QH0692, are statistically significant (Table 2).
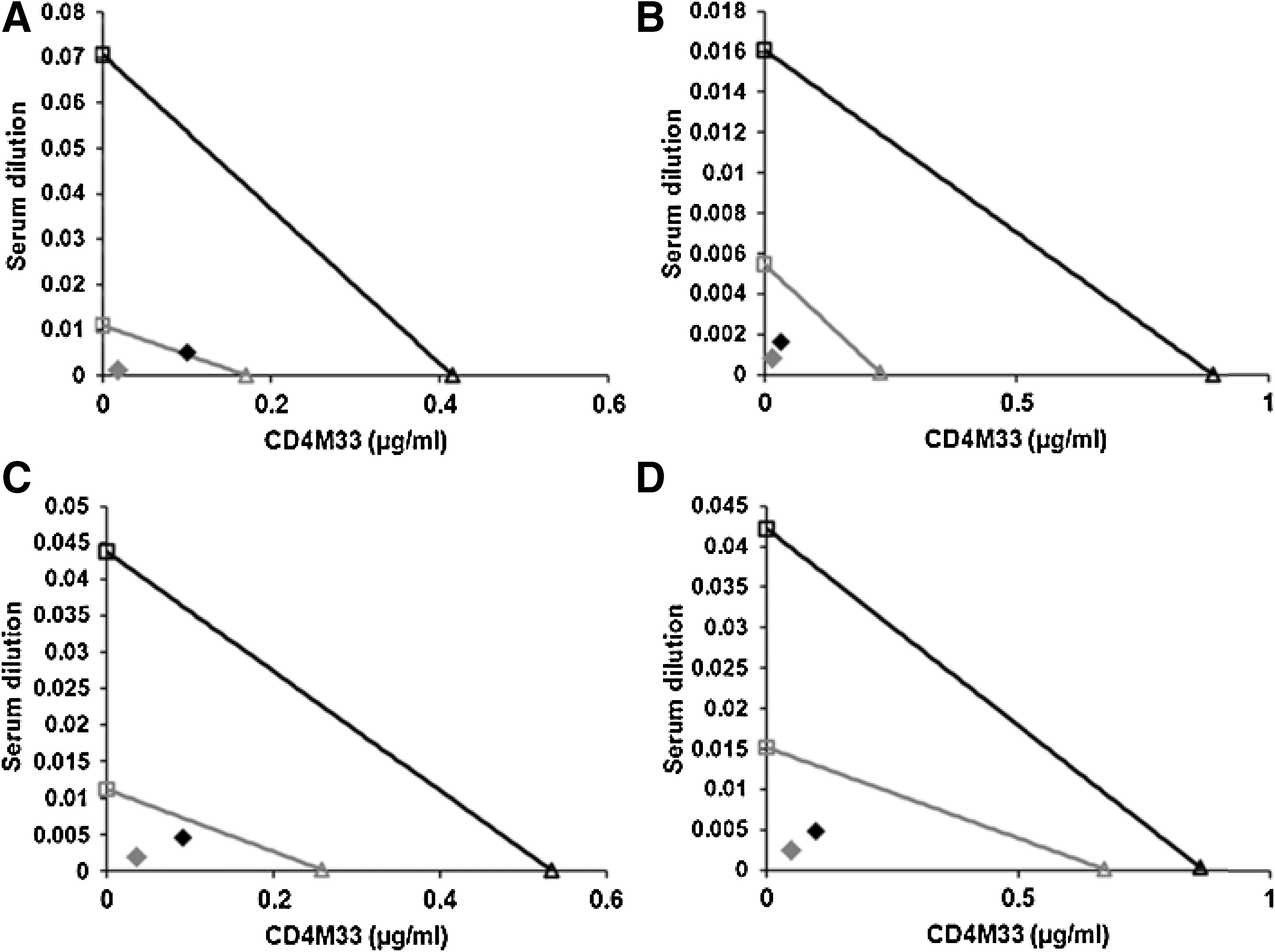
Classical isobologram plot representation of the synergy between the C4-V3T303C-E322C immune serum and CD4M33 in neutralizing HIV-1 strains. The data shown are for the B961 immune serum, for the neutralization of
Overall, the data indicate that QH0692 can be recognized by the C4-V3T303C-E322C immune sera. QH0692 was chosen in this study as a proof of concept since it is one of the more resistant tier 2 viruses and it has been used by others as a reference strain for neutralization assays. 27 Moreover, in earlier studies addition of sCD4 barely resulted in better than 50% neutralization of QH0692 when the V3-directed antibodies 445-52D and 2219 and modified-gp120 immune sera were tested. 6 This is in spite of the observation that these two antibodies and the immune sera showed considerable improvement in neutralization potency of some other tier 2 and tier 3 viruses after incubation with sCD4. Thus, even after the addition of sCD4, QH0692 proved to be difficult to neutralize. Nevertheless, testing the synergy between CD4M33 and C4-V3T303C-E322C immune sera for additional tier 2 viruses would be required to generalize our observation for the vast majority of clade B viruses.
Our findings imply that similarly to sCD4 the synthetic peptide CD4M33 can expose the V3 loop for neutralization by immune sera elicited by a constrained V3 peptide. Most importantly, this occurs even with a representative tier 2 clade B virus, QH0692. This suggests that optimally constrained peptide C4-V3T303C-E322C mimics correctly the native conformation of the V3 region even for tier 2 clade B viruses. The stronger neutralizing antibody response elicited by the optimally constrained V3 peptide, in comparison with a monomeric gp120 immunogen, 10 makes this peptide an attractive candidate for the elicitation of potent V3-directed neutralizing antibodies. Additional advantages of a synthetic V3 immunogen over gp120 immunogens are its stability (unlike the gp120 protein, which may suffer from aggregation, denaturation, and proteolysis), ease of production, and immunogenicity.
As preexposure prophylactic agents, CD4 miniproteins have several advantages over sCD4. Their small size allows them easier access to the CD4 binding site (CD4bs), they are stable in denaturing conditions, and they are relatively resistant to proteolytic degradation as they have three disulfide bonds and nonnatural amino acids. Moreover, CD4 miniproteins are antigenically different from CD4. Together this makes them more attractive as therapeutics and potential preexposure prophylaxis agent. 14,18 Yoshimura et al. reported a synergistic effect between NBD-556, a CD4 mimetic, and the V3-directed antibody KD-247. 9 CI values were determined for only two laboratory adapted viruses. Overall, the CI values of 0.174 and 0.043 for 50% and 75% inhibition, respectively, determined for NBD-556 are somewhat better than those determined for CD4M33 in the present work. Small molecule CD4 mimetics are advantageous over CD4M33 in that they can be given orally and produced in large quantities at low cost. However, CD4M33 is three orders of magnitude more potent in HIV-1 neutralization compared with NBD-556, which neutralizes HIV-1 in the 50 μM range concentration and which, like other small molecule CD4 mimetics, has a small difference between the therapeutic and the toxic concentration. 8,9,30 Therefore, further improvement of the small organic CD4-mimetics is required to make them suitable for practical applications as preexposure prophylaxis. CD4-mimetic compounds have the potential to be used as an additional option to treat patients infected with HIV-1. Such individuals invariably develop very high titers against the V3 and synergism between the vaccine or infection-elicited anti-V3 antibodies should probably lower the required concentration of the small molecule CD4 mimic compound by approximately a factor of 10. This would greatly reduce the toxicity of the mimetics at concentrations at which they would be efficacious. Moreover, the presence of V3-directed antibodies may considerably reduce the risk of higher infection rates of CD4−/CCR5+ or CXCR4+ cells that may be caused by the use of CD4-mimetic compounds. 31 Since CD4 mimetics are entry inhibitors that target the viral envelope protein rather than the human CCR5, they potentially should cause fewer severe side effects, making them attractive candidates as future therapeutic and prophylaxis agents.
Antibodies to the CD4 binding site can potentially induce conformational changes similar to CD4 and some have been reported to act in synergy with V3-directed antibodies. 32 Therefore, a combination of immunogens that can elicit antibodies that induce the CD4 bound conformation and expose the V3, with an optimal V3 immunogen that elicits high levels of anti-V3 antibodies, in principle should provide significantly better protection compared to each immunogen when used alone. The synergism between CD4bs and V3-elicited antibodies may compensate for lower than optimal neutralization titers obtained by immunization with the individual immunogens.
Altogether our results demonstrate a significant synergistic effect between the immune sera elicited by the constrained V3 peptide and CD4M33. This effect is of interest for treating AIDS patients and using the combination of an active immunization with a prophylactic treatment. Challenges for the future will include the development of more potent CD4 mimic compounds and maximization of the breadth and potency of the neutralizing antibody response.
Footnotes
Acknowledgments
We are most grateful to Dr. Orith Leitner and Mr. Ziv Landau from the antibody unit at the Weizmann Institute for the immunizations, animal handling, and good advice and to Drs. Julie Goss, Terri Wrin, and Pham Phung from Monogram Biosciences, South San Francisco, CA for their services in the HIV-1 neutralization assay.
Research reported in this publication was supported by the Kimmelman Center and by the National Institute of General Medical Sciences of the National Institutes of Health under award number GM53329 (J.A.) and GM22087 (F.N.). J.A. is the Dr. Joseph and Ruth Owades Professor of Chemistry and F.N. is the Leonard and Esther Kurtz Term Professor at the College of Staten Island at the City University of New York. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Author Disclosure Statement
No competing financial interests exist.