
Abstract
Mechanisms leading to the observed immune dysregulation in chronic HIV infection are not well understood. The MHC-II class ligand, lymphocyte activation gene-3 (LAG-3, CD223), has been implicated in the complex regulation mechanism of immune functions. In this study, we describe a new population of HIV-specific CD8+ T cells expressing LAG-3. These LAG-3+CD8+ T cells do not display immunophenotypic patterns traditionally attributed to regulatory T cells. The LAG3+CD8+ T cells are CCR7+,CD127−, and display heterogeneous surface expressions of CD45RA and CD25. Interestingly, HIV-specific LAG-3+CD8+ T cells do not substantially express CTLA-4 and LAG-3 expression does not correlate with interleukin (IL)-10 or tumor growth factor (TGF)-β production. In addition, HIV-specific LAG3+CD8+ T cells do not produce interferon (IFN-γ) or express CD107a. The frequency of HIV-specific LAG3+CD8+ T cells negative correlated with plasma viral load. Our study introduces a new population of HIV-specific CD8+ T cells and proposes additional mechanisms of immune regulation in chronic HIV infection.
Introduction
L
Chronic HIV-1 infection is characterized by the progressive loss of immune function and subsequent failure of the immune system to eradicate the infection. Impaired CD8+ T cell function, a common feature of chronic HIV disease, leads to suboptimal viral control. 14 LAG-3 expression is associated with negative regulatory cell-surface patterns that are determinants of CD8+ T cell exhaustion in lymphocytic choriomeningitis virus and HIV infections. 15,16 Elevated plasma LAG-3 levels and lymphatic tissue expression have been associated with advanced HIV disease. 17,18 Simultaneous blockade of the T cell inhibitory receptors PD-1 and LAG-3 has shown improved T cell responses and viral control in vivo. 15,19 Interestingly, LAG-3 is also a marker for recently activated effector T cells, although differential expression of LAG-3 on T cells has not been fully elucidated in HIV infection. 9,15,20
In this study we evaluate the role of LAG-3 in CD8+ effector dysfunction and its association with other negative immune regulators, including PD-1, CTLA-4, and 2B4. We describe a new subpopulation of LAG-3+CD8+ T cells that is antigen specific. Expression of LAG-3 on HIV-specific T cells negatively correlates with viral load, suggesting a possible role in viral control.
Materials and Methods
Study subjects and samples
Peripheral blood mononuclear cell (PBMC) samples from HIV-positive volunteers (n=35) were obtained from the Research in Access to Care in the Homeless (REACH) cohort in San Francisco as previously described. 21 –23 Demographic information and CD4+ T cell count were obtained at the time of enrollment and blood draw. HIV-positive volunteers on antiretroviral treatment (ART) were usually on a protease inhibitor or a nonnucleoside reverse transcriptase inhibitor. CD4+ T cell count and viral loads were measured approximately every month. The CD4+ T cell counts for these samples were 422–1,283×106 cells/liter (median=494×106 cells/liter). Viral plasma loads were undetectable. Institutional Review Board approvals were obtained from the California Department of Public Health and University of California San Francisco Committee on Human Research. All participants in the study gave written and informed consent. None of the study participants had received antiretroviral therapy for at least 6 months. HIV RNA levels were determined from plasma using the Roche Amplicor 1.5 (generously donated by Roche Diagnostic Systems), per the manufacturer's instructions. PBMCs were separated and cryopreserved in liquid nitrogen until assay time.
Antigens
Peptides corresponding to the subtype B consensus sequences of HIV-1 for Gag and Nef were synthesized as 15 amino acids (a.a.) overlapping by 11 a.a. (AIDS Research and Reference Reagent Program, Division of AIDS, NIAID,
Flow cytometry reagents
The following antibodies were used in the assays per the manufacturer's instructions (Becton Dickinson, San Jose, CA): AmCyan-conjugated anti-CD3 (clone SK7), PerCP-Cy5.5 or PE-Cy7-conjugated anti-CD4 (clone SK3), APC-H7-conjugated anti-CD8 (clone SK1), PE-Cy5-conjugated CTLA-4 (CD152; clone BNI3), PE-conjugated anti-CD25 (clone 2A3), PE-Cy7- or APC-conjugated anti-CD45RA clone (clone L48; clone HI100, respectively), PE-conjugated anti-CD107a (clone H4A3), Alexa Fluor 647-conjugated anti-CD127 (clone hIL-7R-M21), or APC-conjugated anti-interferon (IFN)-γ (clone 25723.11). FITC-conjugated anti-LAG-3 staining was used according to the manufacturer's instructions (CD223; clone 17B4, Burlington, NC). All samples were acquired using a multichannel (eight-color) BD LSRII flow cytometer (Becton Dickinson, San Jose, CA).
Flow-based CD107a degranulation assay
The degranulation assay was performed as previously described. 23,26 Briefly, PBMCs (1×106) were stimulated with HIV (Gag or Nef) or human CMV peptide pools and incubated in the presence of anti-CD107a-PE (BD Biosciences), 1 μg/ml costimulatory anti-CD49d and anti-CD28, and Golgi-Stop (BD Biosciences) for 6 h at 37°C in 5% CO2. Staphylococcus enterotoxin B (SEB) (1 μg/ml; Sigma-Aldrich) and media alone were used as positive and negative controls, respectively. Cells were subsequently stained with anti-CD45RA PE, anti-CCR7 PE CY7, anti-CD3 PerCP Cy5.5, and anti-CD8 allophycocyanin Cy7, CD4 Pacific Blue (BD Pharmingen). A minimum of 30,000 CD8+ events/sample was acquired by flow cytometry. Percentages of CD3+CD8+ T cells expressing CD107a were determined after subtraction of background activity using preset gating. All volunteers demonstrated significant CD107a expression following SEB stimulation.
Intracellular cytokine flow cytometry
IFN-γ production was performed as previously described. 23 PBMCs (1×106) were incubated with SEB or peptide pools for 6 h at 37°C in 5% CO2 in the presence of costimulatory anti-CD28 and anti-CD49d (1 μg/ml, Becton-Dickinson) and Golgi-Stop (BD Pharmigen). Alternatively, PBMCs were stimulated for 24 h in the absence of Golgi-Stop followed by restimulation with SEB or peptide pools in the presence of Golgi-Stop for an additional 6 h. A minimum of 30,000 CD3+ T cells per sample was acquired using eight-color flow cytometer (LSRII, BD Biosciences) and analysis was performed by FlowJo software (TreeStar, San Carlos, CA).
LAG-3 surface expression on antigen-specific CD8+ T cells
Detection of HIV-specific LAG-3 or CTLA-4 expressing CD8+ T cells was performed using PBMCs (1×106) incubated with either Gag, Nef, or CMV pp65 peptide pools incubated for 24 h at 37°C in 5% CO2 in the presence of costimulatory anti-CD49d and anti-CD28 (1 μg/ml, BD Biosciences). Cells were further stained with the following abs: LAG-3-FITC, CTLA-4-PE-Cy5, CD3-AmCyan, CD8-APC-H7, 2B4 PE (CD244), CD25-PE, CD127-Alexa Fluor 647, PD-1 APC (CD279), and CD45RA-PE-Cy7. PBMCs were fixed with BD FACS Lysing Solution and a minimum of 30,000 CD3+ T cells per sample was acquired on the flow cytometer (BD LSRII) and analyzed using FlowJo software. Stimulation with SEB (1 μg/ml, Sigma-Aldrich) was used as a positive control for the expression of LAG-3. Media alone was used as a negative control. To detect CD107a and IFN-γ on LAG-3-expressing CD8+ T cells, PBMCs were stimulated as described above, washed twice with phosphate-buffered saline (PBS) buffer, and restimulated with peptides in the presence of anti-CD28, anti-CD49, and Golgi-Stop (BD Biosciences) for an additional 6 h and analyzed for the expression of CD107a-PE and IFN-γ-APC. A minimum of 30,000 CD3+ T cells was acquired on the flow cytometer (BD LSRII) and analyzed using FlowJo software.
Statistical analysis
Groups were compared using the Mann–Whitney U test and analysis was performed with PRISM software version 5.0 (Graph-Pad). Correlations were determined using the Spearman rank correlation test (Graph-Pad). This statistical test like the Mann–Whitney U test is a nonparametric test and does not assume normality, homoscedasticity, or linearity and is calculated by rank. Statistical significance was defined as p<0.05.
Results
LAG-3 is expressed on HIV-specific CD8+ T cells and does not correspond with IFN-γ or CD107a expression
In this study we assessed the frequency of LAG-3, IFN-γ, and CD107a expression on Gag- and Nef-specific CD8+ T cells in 35 HIV-1-infected volunteers. Through the standard 6-h intracellular protocol we are able to detect robust IFN-γ and CD107a (Fig. 1A); however, we are unable to detect surface expression of LAG-3 (data not shown). We therefore developed an assay that would allowed the simultaneous measurement of surface expression of LAG-3 and intracellular IFN-γ and CD107a (Fig. 1B). Twenty-three and six volunteers demonstrated significant LAG-3 expression on Gag- and Nef-specific CD8+ T cell responses, respectively [median=0.29% (range, 0.07–1.24%) and 0.21% (range, 0.09–0.35) for Gag and Nef, respectively]. Eight volunteers demonstrated significant LAG-3 expression on CMV pp65-specific CD8+ T cell responses [median=0.22% (range, 0.09–0.53%)]. No overlap between HIV-specific CD8+ T cells expressing LAG-3 and IFN-γ or CD107a was observed (Fig. 1C). LAG-3 frequencies were significantly higher on Gag- but not on Nef- or CMV pp65-specific CD8+ T cells compared to no antigen-stimulated cells (p<0.0001, p=0.73, and p=0.91, respectively; data not shown). Differences in the frequencies of LAG-3 expression on Gag-specific were significantly higher compared to Nef-specific and CMV pp65-specific CD8+ T cell responses (p<0.0001 and p=0.0002, respectively) (Fig. 1D).

Lymphocyte activation gene-3 (LAG-3+) expression on HIV-specific CD8+ T cells does not express CD107a or produce interferon (IFN)-γ.
Immunophenotyping of HIV-specific LAG-3+CD8+ T cells
Specific CD8+ T cell functions have been ascribed to distinct populations defined by immunophenotyping, including expression of the homing receptor CCR7, the IL-7 receptor CD127, and different isoforms of CD45RA. 27 We examined the level of CD25, CD127, and CD45RA expression on HIV-specific CD8+ T cells expressing LAG-3. Representative plots of the phenotype of the HIV-specific CD8+ T cell population expressing LAG-3 are shown in Fig. 2. LAG3+CD8+ T cells were CD25 positive, CD127 negative and demonstrated a heterogeneous phenotype expression of CD45RA (Fig. 2A) and CCR7 (Fig. 2B).
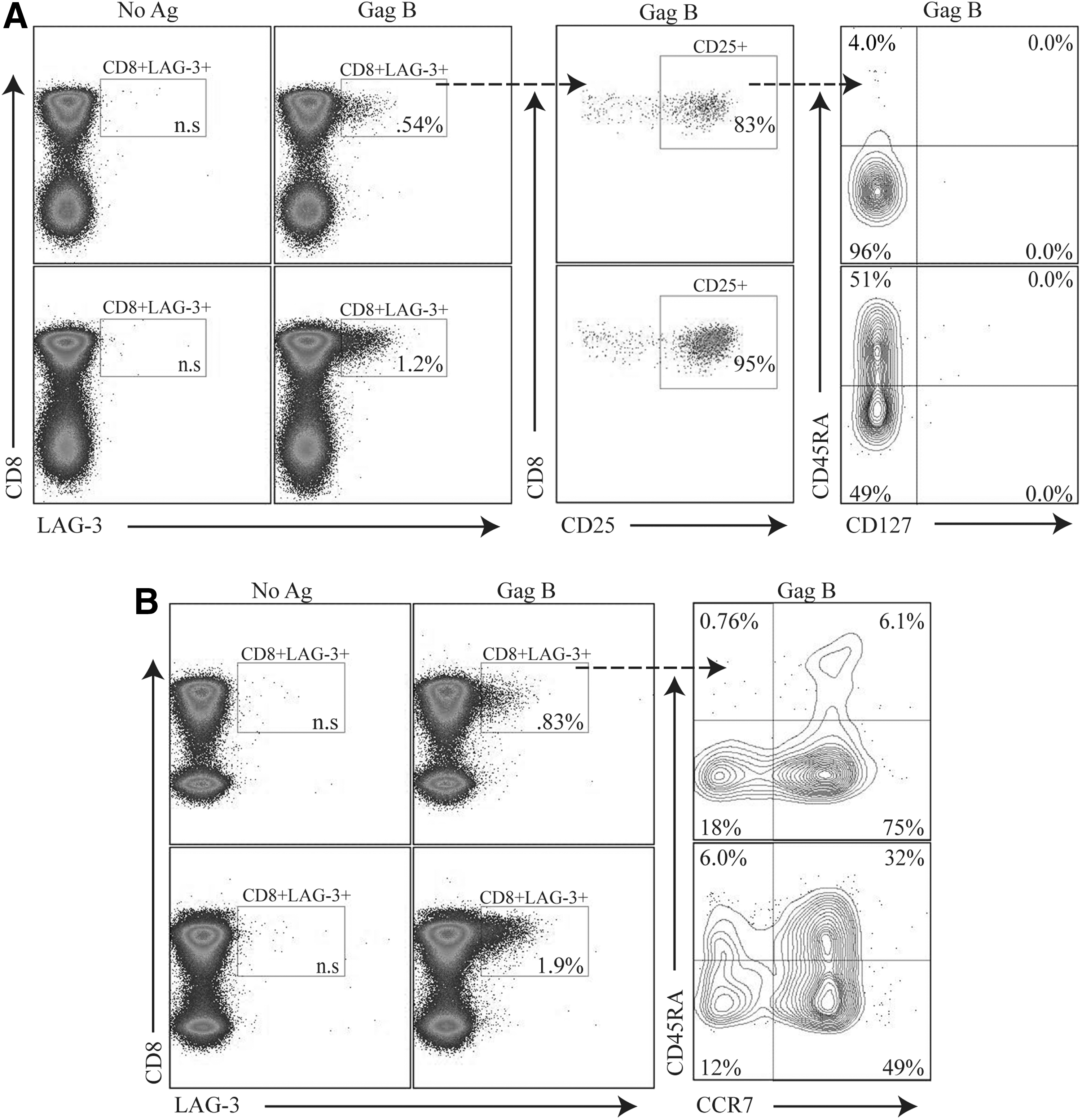
HIV-specific LAG-3+CD8+ T cells express a variable surface immunophenotype. PBMCs were stimulated with HIV peptides then stained with Vivid for live/dead cell exclusion, LAG-3 FITC, CTLA-4 PE-Cy5, CD3 AmCyan, CD8 APC H7, CD25 PE, CD127 Alexa Fluor 647, and CD45RA PE-Cy7 and analyzed by flow cytometry. Samples were first gated on SSC/FSC, followed by SSC/CD3+, and then on the CD3+/CD8+ lymphocyte population; the percent of LAG-3+CD8+ T cells was then determined.
LAG-3 expression on HIV-specific CD8+ T cells does not correlate with markers of T cell exhaustion
Exhausted HIV-specific CD8+ T cells express high levels of programmed death-1 (PD-1). 28 –30 The upregulation of CTLA-4 on HIV-specific T cells is also associated with immune dysfunction. 31 Expression of 2B4 on CD8+ T cells has been associated with HIV disease progression and decreased killing activity. 32,33 However, recent studies on HIV-specific CD8+ T cell exhaustion has been shown to be associated with the complex expression of PD-1, CTLA-4, and 2B4 but not LAG-3. 16 We found no evidence of PD-1, CTLA-4, or 2B4 coexpression on CD8+ T cells that express LAG-3 (Fig. 3), consistent with previous studies. 16 In addition, no tumor growth factor (TGF)-β or interleukin (IL)-10 production was detected upon peptide stimulation in these Lag3+CD8+ T cells (data not shown). These results suggest that HIV-specific LAG-3+CD8+ T cells represent a unique population distinct from previously described effector or suppressor CD8+ T cells. 26,34,35
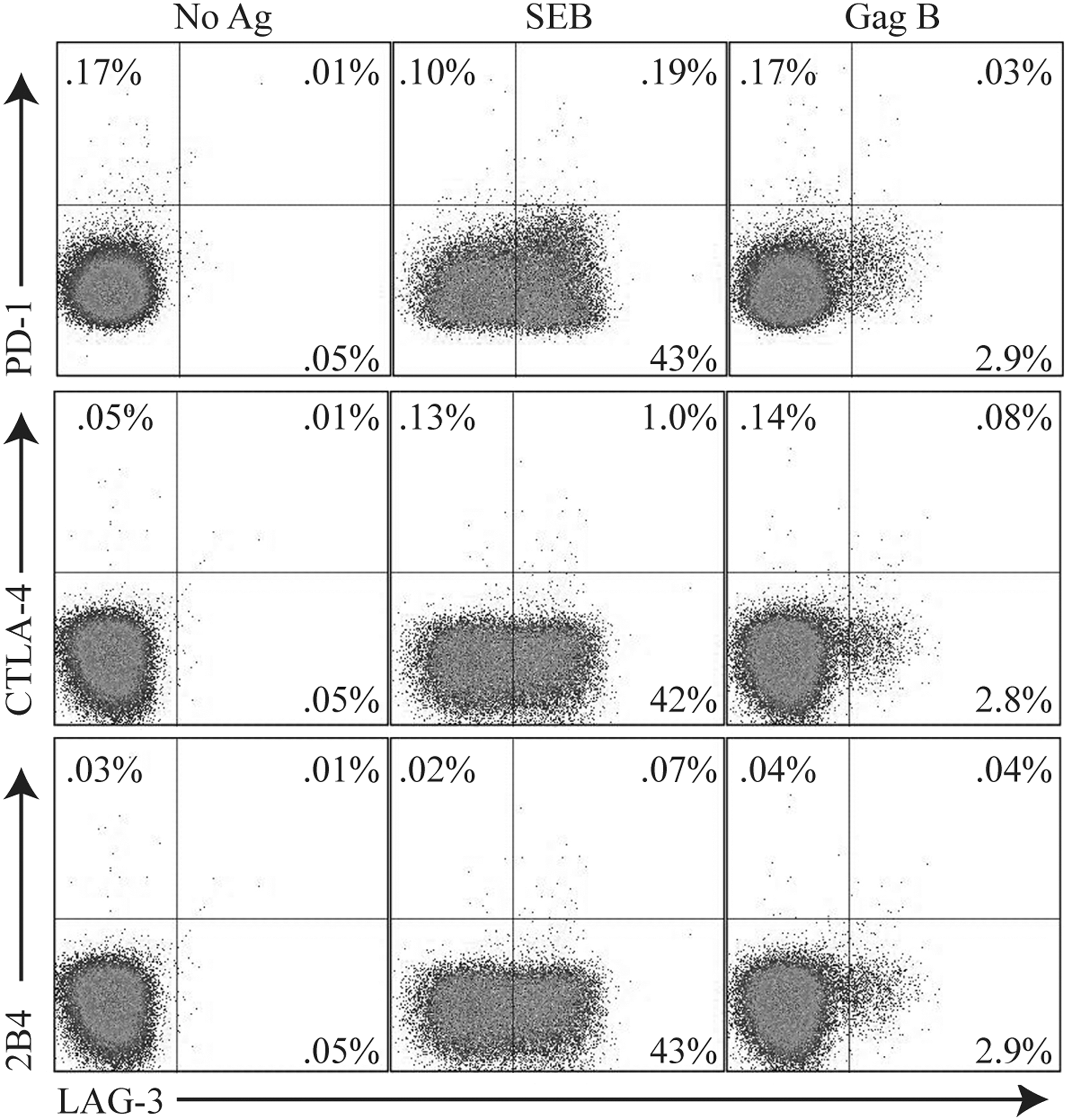
LAG-3+CD8+ T cell populations are distinct from PD1+, CTLA4+, or 2B4+ CD8+ T cells. PBMCs were stimulated with HIV-specific peptide Gag B peptides then stained with Vivid for live/dead cell exclusion, LAG-3 FITC, 2B4 PE, CD3 AmCyan, PD-1 APC, CTLA-4 PE-Cy5, CD4 PE-Cy7, and CD8 APC-H7. Samples were first gated on the CD3+/CD8+ lymphocyte population; the percent of LAG3+, PD-1+ CTLA4+, or 2B4+ cells was then determined. Representative plots of the frequencies of the LAG-3+ Gag-specific CD8+ T cells and expression of PD-1+ CTLA4+ or 2B4+ are shown. Responses ≥0.1% and two times the background were considered positive.
LAG-3 expression on HIV-specific CD8+ T cells negatively correlates with HIV RNA
Plasma HIV-1 viral load is closely correlated with disease progression and the level of immune activation. 36 Functional impairment of HIV-specific T cells during chronic HIV infection is closely linked to viral replication and is thought to be due to T cell exhaustion. 28,37 Surprisingly, we found a significantly negative correlation between the frequency of the LAG-3 positive, HIV-specific CD8+ T cells, and plasma HIV RNA (r s=−0.51, p=0.0008) (Fig. 4A). No correlation between GAG-specific LAG-3-positive and CTLA-4-positive CD8+ T cells was observed (data not shown).
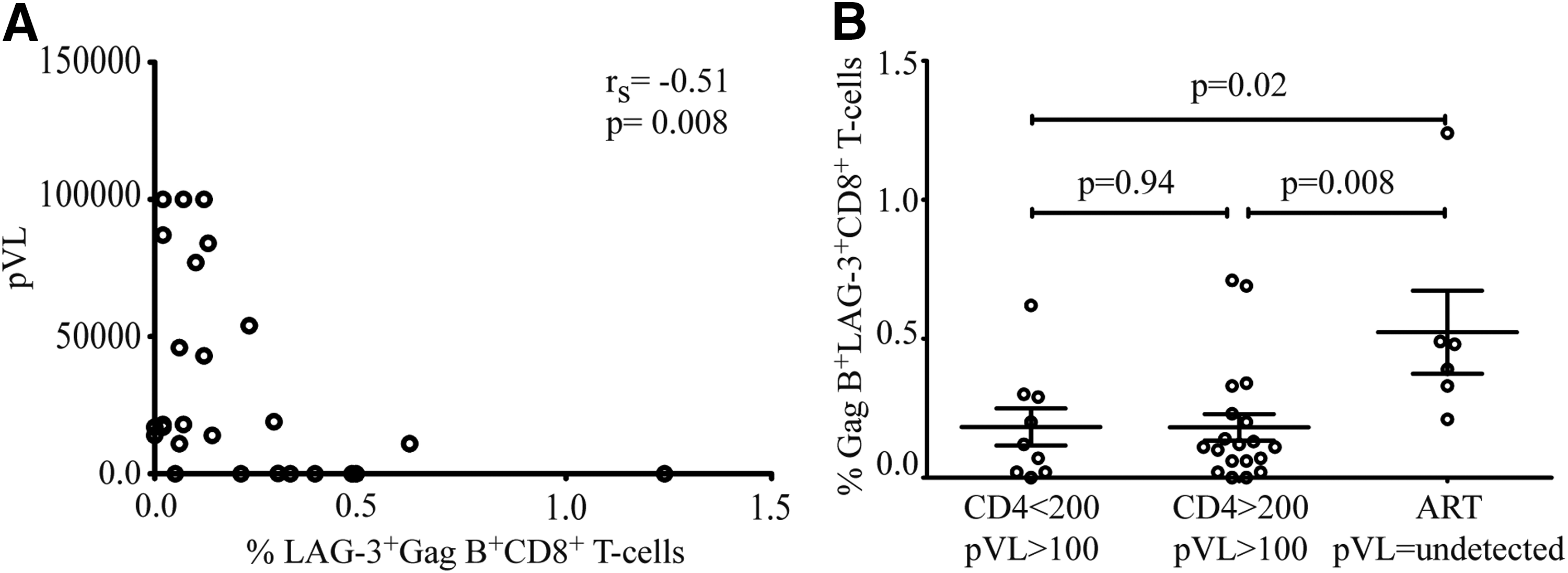
Negative correlation between the frequency of HIV-specific LAG-3+CD8+ T cells and HIV plasma RNA. The percentage of Gag-specific LAG-3 is plotted against the HIV plasma RNA. A significant negative correlation between HIV plasma RNA and Gag-specific LAG-3+CD8+ T cells in
Next we stratified our data based on CD4 count and viral suppression into three groups and analyzed Gag B LAG-3+CD8+ frequency. We find that ART-mediated viral suppression correlated with a higher expression of LAG-3 on Gag B-specific CD8+ T cells in individuals with CD4 counts >200 compared to samples from individual not receiving ART with CD4 counts <200 or CD4 >200 counts (p=0.02 and p=0.008, respectively).
Discussion
LAG-3, a transmembrane protein that binds MHC class II, has been previously associated with negative regulation of cellular proliferation, activation, and homeostasis. 38 Although LAG-3 has been reported on regulatory T cells, it remains unclear whether LAG-3 expression on CD8+ T cells confers similar regulatory functions in HIV infection. In vitro studies have shown that CD8+LAG-3+CD25+ T cells have a suppressor function imparted through CCL4. These cells also expressed CTLA-4, FoxP3, and CD107a but no IFN-γ. Additionally these cells were described to be mostly CD45RA− and CD127−. 38 In these studies we show that Gag-B-specific CD8+LAG-3+CD25+ cells were induced and are CTLA-4, CD107a, and IFN-γ negative. Unlike the above mentioned study we found CD45RA to be heterogeneously expressed and CD127−. In addition, these cells did not produce IL-10 or TGF-β.
Although it is tempting to label these Gag-B specific CD8+LAG-3+CD25+ T cells as suppressor T cells, we cannot rule out the possibility that these cells are activated effector. LAG-3 expression has also been shown to be a marker of recently activated effector cells. 9 The expression of CD25 has also been shown to be upregulated on activated T cells. 27 Moreover, phenotypic analysis of these cells shows them to be CD127− and a heterogeneous expression of CD45RA and CCR7. The mix phenotypic profile of these Gag-B-specific CD8+LAG-3+ T cells points to an effector-like [effector (CD45RA+CCR7−) or effector memory (CD45RA−CCR7−)] phenotype. 39,40
Here, we show that LAG-3 expression is antigen specific and can be delineated from other CD8+ T cells, both inhibitory and effector. Surface LAG-3 expression on activated human T cells is upregulated by interleukin IL-2, IL-7, and IL-12 and thus may play an essential role during activation. 41 LAG-3 has been associated as an inhibitory marker and in immune suppression. 3,8,9,11,15,16,42,43
Given the similarities in downstream targets, it is tempting to speculate that inhibitory molecules may act antagonistically against other positive costimulation. The presence of LAG-3+ HIV-specific-positive CD8+ T cells was associated with a significantly lower HIV RNA level in our study, arguing that these cells do not have a similar regulatory effect previously ascribed to suppressor T cells. Analysis of the surface marker expression of the LAG-3+ Gag-specific CD8+ T cells displayed a heterogeneous pattern of surface immunophenotypes that does not allow distinction between inhibitory and effector phenotypes. It is not yet clear whether accumulation of T cells expressing various inhibitory or effector molecules is a consequence of chronic HIV-specific immune activation. The direct and indirect role of LAG-3+ HIV-specific CD8-positive cells in cytokine production and CD8+ lytic ability remains unknown. It is likely that many pathways in addition to those activated by inhibitory receptors are involved in the events leading to T cell exhaustion.
We propose that LAG-3 expression by HIV-specific CD8+ T cells identifies a new antigen-specific CD8+ T cell population. Larger longitudinal studies will provide more conclusive answers on the functional role of these CD8+ T cells in T cell homeostasis and immune recovery.
In conclusion, our data show that LAG-3 is upregulated on CD8+ T cells that are HIV specific. Levels of LAG-3 are highest in individuals with lower viral load. A mechanistic understanding of how immune modulation receptors restrain or promote T cell responses would not only aid in our understanding of HIV immunopathogenesis, but could also provide novel targets for therapeutics.
Footnotes
Acknowledgments
This work was supported by NIH grants AI43885, MH54907, AI71772, and K24 MH87227.
Author Disclosure Statement
No competing financial interests exist.