
Abstract
Mother-to-child transmission of HIV-1 subtype C can occur in utero, intrapartum, or via breast milk exposure. While not well understood, there are putative differences in the mechanisms involved with the distinct routes of vertical HIV transmission. Here, we address the question of whether specific viral characteristics are common to variants transmitted through breastfeeding that may facilitate evasion of innate or adaptive immune responses. We amplified the envelope gene (env) from the plasma of six infants during acute infection who were infected with HIV-1 subtype C through breastfeeding, and from three available matched maternal samples. We sequenced the full-length env genes in these subjects revealing heterogeneous viral populations in the mothers and homogeneous populations in the infants. In five infants, the viral population arose from a single variant, while two variants were detected in the remaining infant. Infant env sequences had fewer N-linked glycosylation sites and shorter sequences than those of the available matched maternal samples. Though the small size of the study precluded our ability to test statistical significance, these results are consistent with selection for virus with shorter variable loops and fewer glycosylation sites during transmission of HIV-1 subtype C in other settings. Transmitted envs were resistant to neutralization by antibodies 2G12 and 2F5, but were generally sensitive to the more broadly neutralizing PG9, PG16, and VRC01, indicating that this new generation of broadly neutralizing monoclonal antibodies could be efficacious in passive immunization strategies.
T
Studies of in utero and intrapartum transmission have shown a universal bottleneck in genetic diversity from mother to child, as well as differences in the characteristics of transmitted virus for in utero versus intrapartum transmission. 12,13 Data are very limited for breastfeeding pairs, but one study of three breast milk transmission events found a similar bottleneck for HIV-1 subtype A. 14 We previously demonstrated that the viral population found in infants infected intrapartum tended to be more heterogeneous than populations from infants infected in utero, and that viruses from infants infected intrapartum tended to have shorter variable loops and fewer glycosylation sites than the parent viral population. 13,15 Others had similar findings for infants mostly infected intrapartum, though they only found differences in glycosylation sites and not variable loop lengths. 16
In this article, we present data on viral sequences from three mother–child pairs plus three additional infants where the infant became HIV DNA positive between 6 and 12 weeks postpartum through exposure to breast milk, with the goal of exploring whether known selective pressures influence this route of mother-to-child transmission. Participant plasma samples were collected from the Malaria and HIV-1 in Pregnancy (MHP) prospective cohort. 17 –20 The MHP study was approved by the Malawi College of Medicine Research Committee and the Institutional Review Board at the University of North Carolina at Chapel Hill. Informed consent was obtained from all mothers. Women and their newborn infants received single-dose nevirapine according to the HIVNET 012 protocol 20 and breastfeeding was initiated. Plasma and cell pellets were isolated from blood collected from the mothers at labor-ward admission, and blood was collected via heel-sticks from the infants at three time points: within 48 h of birth, at 6 weeks, and at 12 weeks of age. Infants who were HIV-1 DNA negative by real-time polymerase chain reaction (PCR) 21 at 0 and 6 weeks, then positive at 12 weeks were classified as infected postpartum through breastfeeding (BF). Viral RNA and cell-associated DNA were isolated and amplified using single-genome env gene amplification, as has been described previously, 15,22 to ensure that most amplifications were initiated with a single template without artifactual recombination during PCR between multiple template sequences. The HIV-1 env DNA single genome amplification protocol was the same as the RNA protocol following reverse transcription (GenBank accession numbers JN983803–JN983805).
Sequences were aligned using the L-INS-I method in MAFFT version 5.8.
23
A maximum likelihood phylogenetic tree was constructed with PHYML
24
using the general time reversible plus gamma (α ∼0.25 for each tree) evolutionary model chosen by FindModel (
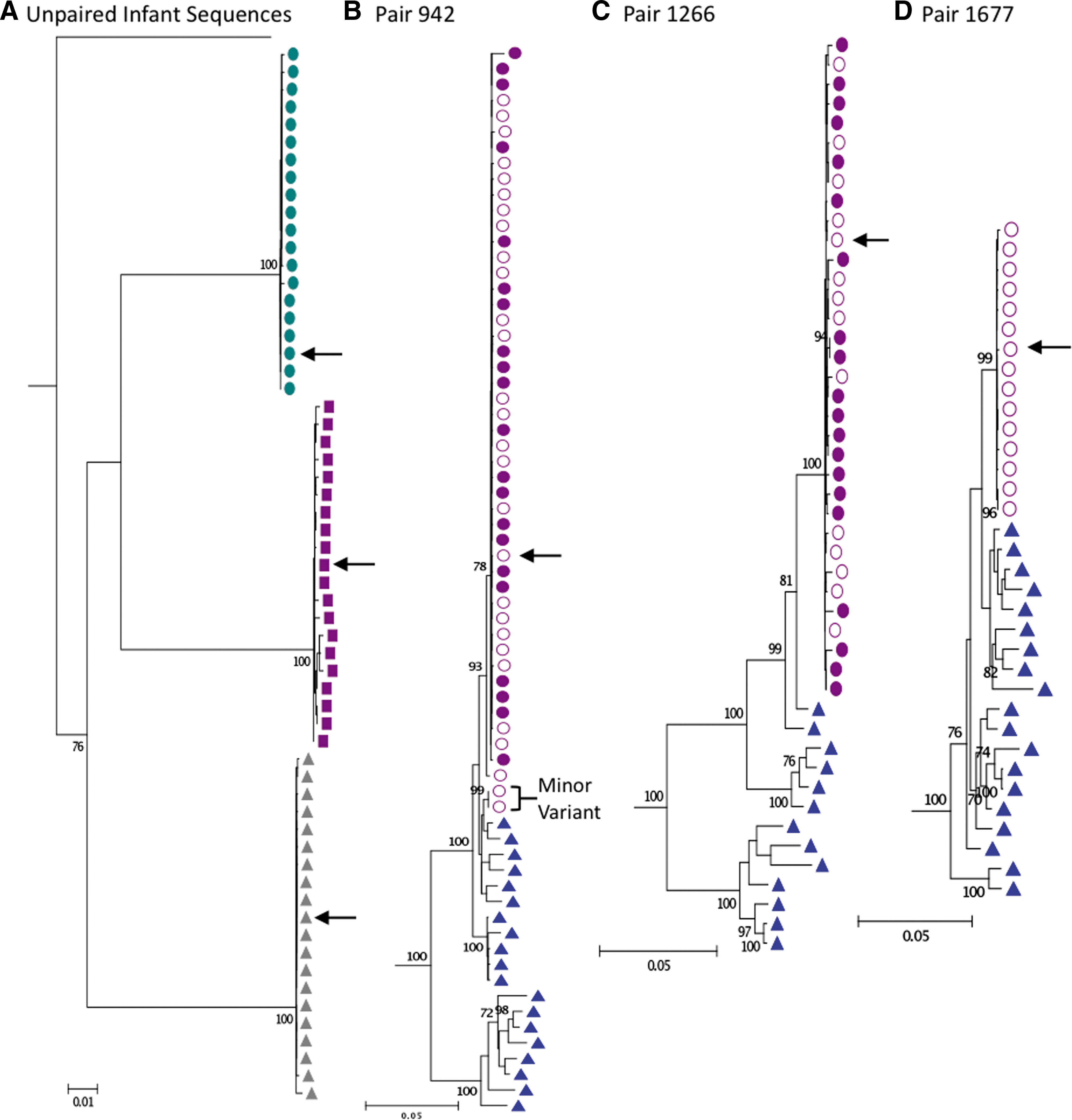
Maximum likelihood phylogenetic trees for
In maternal–infant pair 942 there is evidence for the transmission/replication of two maternal variants in the infant (Fig. 1). Using a Highlighter plot (
We next compared the number of putative N-linked glycosylation sites (N-glycosite program,
The presence of multiple variants in infant 942 could represent separate transmission events, or multiple variants simultaneously transmitted in a single event. It is worth noting that the two variants clustered on the same tree branch compared to the more diverse maternal viral population (Fig. 1). This phenomenon was also seen in an IP transmission pair for HIV-1 subtype B. 36 This similarity between variants could be stochastic or it could suggest selection for certain characteristics (in breast milk, during transmission, or through selective amplification in the infant). 37 –39 Other studies have presented conflicting results testing the hypothesis that selection is influenced by neutralizing antibodies, and transmitted variants are generally resistant to neutralization. 14,36,39 Thus, we investigated the neutralizing antibody resistance profile of these subtype C infant viral variants transmitted through breast milk for comparison.
The consensus (or transmitted/founder) infant env gene (or a close match with 1 nt change for 942 and 1266) from each infant was cloned as previously described 15 (Fig. 1). Viruses were assayed as Env pseudotyped viruses for sensitivity to soluble CD4 (sCD4) and to a panel of monoclonal antibodies [2G12, 2F5, 4E10, IgG1b12, PG9, PG16 (gifts of Dennis Burton), VRC01 (gift of John Mascola), CH31 (gift of Bart Haynes)], and subtype C HIVIG. Titers are reported as the antibody concentration or reciprocal serum dilution with a 50% reduction in relative luminescence units, as previously described. 40 Values were interpolated using 5 parameter curve-fitting. As expected for subtype C virus, no pseudotyped infant virus was sensitive to monoclonal antibodies 2G12 or 2F5 up to a concentration of 25 μg/ml. 40 Five of the six pseudotyped infant viruses were sensitive to 4E10 and PG16 within this range, while 1677 was resistant to both. All infant viruses were sensitive to PG9, and all but 942 were sensitive to VRC01 and CH31.
The panel of neutralizing antibodies used in this study represents both the previous generation of monoclonal antibodies (mAbs) with potent neutralization of subtype B HIV-1, yet importantly also includes more recently identified mAbs with extensive breadth even against the previously “difficult to neutralize” HIV-1 subtype C. The resistance profiles found for the infant clones described here are similar to other studies of newly infected infant viral variants of several HIV-1 subtypes. 14,16,36,41 While we saw little neutralization to 1b12 (Table 1) compared to some other studies where 50% or more of viruses were sensitive, 41,42 the results were similar to our previous study of uncultured Envelope pseudotypes from virus newly transmitted from mother to child. 15 Only one study of three infants infected through breast milk 39 also analyzed sensitivity to the recently identified broadly neutralizing PG9 and PG16 antibodies. 43,44 Similarly, all six infant clones tested herein were sensitive to PG9, and all but one were sensitive to PG16, which have a conformational epitope in the V1/V2 region. In addition, the sensitivity of these variants to the CD4 binding-site antibodies VRC01 and CH31 further demonstrates that transmitted infant subtype C viruses are not inherently resistant to neutralization and that recently transmitted viruses may have neutralizing epitopes in common among all prevalent subtypes. These antibodies clearly have enhanced breadth that includes neutralization of subtype C, and for this reason may represent a useful tool for passive immunization in regions where clade C HIV-1 is prevalent. 41 These data in particular support the CD4 binding site as a promising target for infant vaccine design.
MHP, malaria and HIV-1 in pregnancy.
In summary, we analyzed env sequence data from six infants infected with HIV-1 subtype C through breastfeeding, including three transmission pairs. There was a strong genetic bottleneck during transmission, and more than one variant was transmitted in only one of six infants. Viral envelopes were generally resistant to the monoclonal neutralizing antibodies 2G12 and 2F5, yet were generally sensitive to 4E10 and the more recently identified antibodies PG9, PG16, VRC01, and CH31. The neutralizing sensitivity of these postnatally transmitted virus variants supports the use of the new generation of broadly neutralizing antibodies directed against quaternary V1/V2 epitopes or the CD4 binding site for infant passive immunization strategies, and also implicates these epitopes in infant HIV vaccine design.
Footnotes
Acknowledgments
We thank the Malawian mothers and infants for their participation, the MHP nursing staff and technicians for excellent logistical and technical support, and Jesus Salazar Gonzalez for helpful advice in the preparation of this manuscript. This research was supported by NIH awards to S.R.M. (R21-AI49084) and R.S. (R37-AI44667), and by the UNC CFAR (P30-AI50410). E.R. was supported by an NIH Training Grant (T32 AI07419).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.