
Abstract
Although it has been suggested that biological differences among HIV-1 subtypes exist, their possible influence on disease progression has not been fully revealed. In particular, the increasing emergence of recombinants stresses the need to characterize disease presentation in persons infected by these diverse HIV-1 forms. We explored this issue among 83 Brazilian subjects infected with either HIV-1 subtype B or recombinant subtype BF, all followed since incident infection in a cohort study. Viral subtypes were assigned by full length sequencing of HIV-1 genomes. We observed that the baseline measures for CD4+ T cells and viral load did not differ between the groups. However, longitudinal analysis revealed that subtype BF was clearly associated with a faster CD4+ T cell decline compared to infection with subtype B, in spite of a similar plasma HIV-1 load. While subtype B-infected subjects presented a loss of 3.6 CD4+ T cells/μl per month, subtype BF-infected individuals showed a monthly decay of 6.3 CD4+ T cells/μl (p<0.01). The time to reach 350 CD4+ T cells/μl and the time to start antiretroviral treatment were also shorter in subtype BF-infected persons. The elucidation of an accelerated CD4+ T cell loss associated with subtype BF suggests that this HIV-1 genetic form could be more pathogenic than subtype B.
R
Currently, BF CRFs are the most widespread form of HIV-1 after subtype B and reach a high prevalence (10–50%) in Argentina, Brazil, Chile, Paraguay, and Uruguay. 4 –7 Brazil has the highest number of people living with HIV-1 in the region (estimates from 460,000 to 810,000) 8 and, as in Europe and North America, subtype B is the major circulating genetic clade. In this country, five distinct BF CRFs have been described to date (CRF28_BF, CRF29_BF, CRF39_BF, CRF40_BF, and CRF46_BF). 6 By the year of 2006, 62% of AIDS cases reported in Brazil were confined to the Southeast region, with 70% of these cases from São Paulo, the most populous Brazilian city. Interestingly, up to 11% of the HIV-1 isolated in São Paulo to date has shown a mosaic B/F genome. 5,7
The increasing prevalence of new CRFs and URFs poses a problem in controlling the AIDS epidemics given potential differences in resistance profiles, response to antiretroviral therapy (ART), and other biological properties of the variants. Although different transmission effectiveness, preferential routes of infection, and intrahost replication fitness have long been suggested, 9,10 debate still remains as to whether differences among viral subtypes would influence disease progression. Of note, a small number of studies have examined the effect of HIV-1 subtype on clinical outcome. 11 –13 However, most of them included only seroprevalent cases or a mixture of incident and prevalent cases and have reported subtype assignment based on partial genome sequencing, which cannot exclude the presence of recombinants. To date, no study has investigated distinctive disease outcomes when comparing the two most prevalent HIV-1 forms in Southeast Brazil, subtypes B and BF.
Here, we examined the effect of being infected with either HIV-1 subtype B or BF in a group of individuals followed since the incident infection for a mean of 5 years. Our cohort of recently HIV-1-infected subjects in São Paulo, Brazil 14,15 started accrual in 2002 based on the Serologic Testing Algorithm for Recent HIV Seroconversion (STARHS) strategy. The 113 volunteers who were enrolled in our cohort from June 2002 to January 2006 were selected for this study. All patients provided informed written consent and the study was approved by the University of São Paulo's and by the Federal University of São Paulo's Institutional Review Boards. Peripheral blood mononuclear cells and buffy coat samples were obtained at the patients' first clinical visit and were used for genomic DNA isolation with QIAamp DNA Blood (Qiagen, Hilden, Germany). Five overlapping fragments of the HIV-1 genome were amplified from proviral DNA in duplicate to avoid assembling a whole genome from heterogeneous targets. 7 Phylogenetic analysis and screening of recombinants were carried out as previously described. 7
Near full length genome (NFLG) sequences were obtained for viruses from 83 participants out of the 113. Thirty had only partial genomes sequenced and were not included in subsequent analysis. The epidemiological (gender, age, sexual behavior) and clinical (specific coinfections, CD4+ T cell count, plasma viral load) characteristics of the 83 individuals with NFLG sequencing did not differ from the 30 other individuals whose viruses' genomes were only partially sequenced, except by plasma viral load (VL) at first clinical visit (NFLG: 4.3±0.9 vs. partial: 3.6±0.8; mean±SD log10 copies/ml; p<0.001; Supplementary Table S1; Supplementary Data are available online at
Phylogenetic study of the 83 NFLGs revealed 70 pure (84.3%) and 13 recombinant (15.7%) subtypes. Sixty-eight (82%) sequences clustered with subtype B. Ten (12%) viral genomes with evidence of recombination were mosaic isolates of BF1 URF variants. Their sequences did not present recurrent locations for breakpoints and did not resemble the other BF mosaics already described. 7 Throughout the observed period of enrollment into the cohort, subtype B constantly presented the highest prevalence. Non-B subtypes were observed in the study population after September 2003. Since the enrollment of the first subtype BF-infected subject (November 2003), infections with this subtype have accounted for up to 12% (Fig. 1A). According to our findings, previous reports have shown a prevalence of 9% to 11% for subtype BF infection in São Paulo. 5,7,10
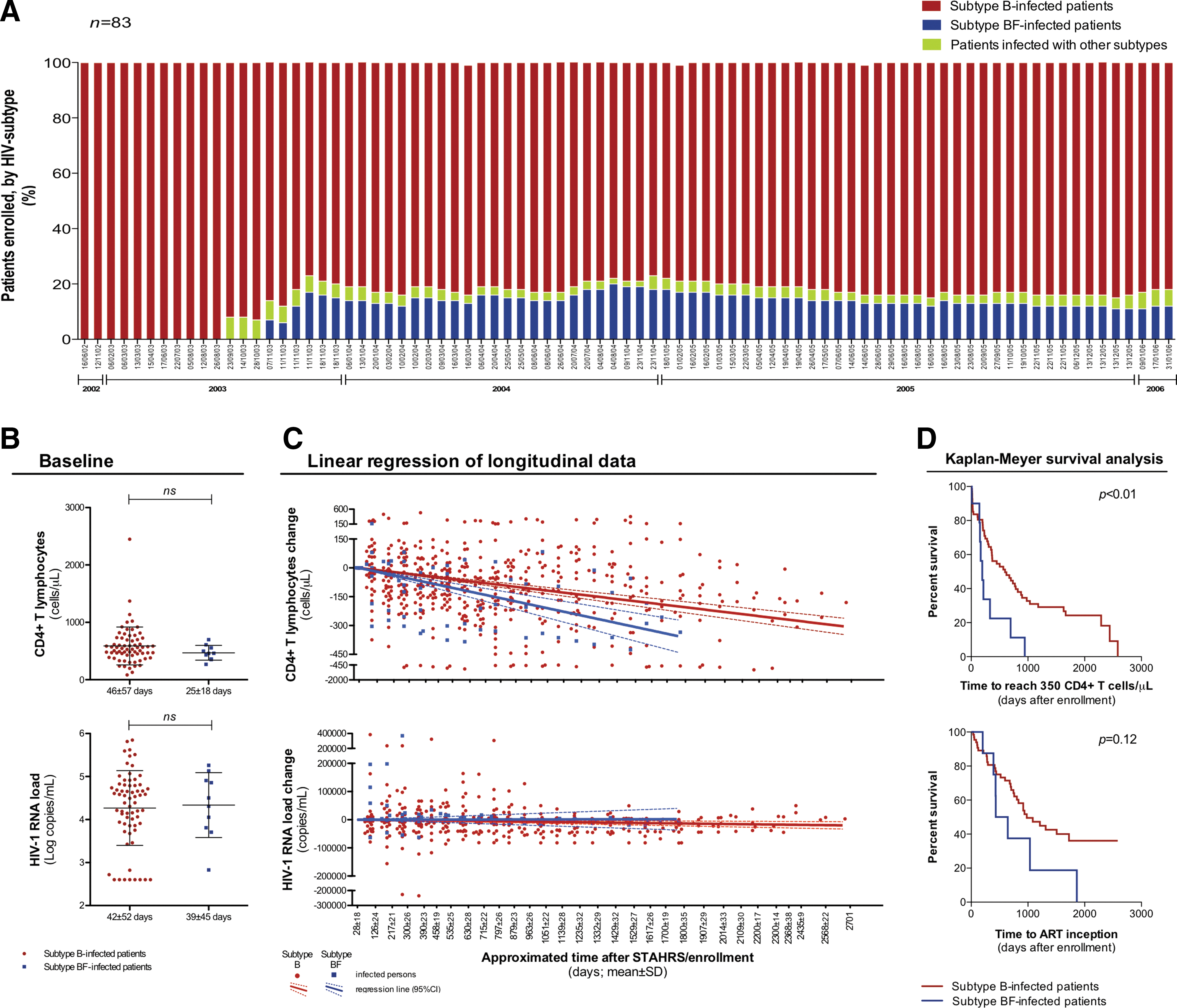
Enrollment of HIV-1 subtype B-infected and BF-infected subjects in a cohort study and comparative analyses of the clinical progression they presented.
We then attempted to investigate differences in clinical progression between subtype B-infected and BF-infected patients. To avoid biases in this approach, we first compared gender, age, date of enrollment/STARHS result (which mirrors approximately 170 days after infection
14
), time of follow-up, and sexual behavior. We also assessed the polymorphism ccr5Δ32 in the human gene ccr5 and viral tropism for CCR5 or CXCR4 coreceptors, assuming they could play a role in disease progression. Furthermore, HIV-1 coinfections with human T cell lymphotropic virus (HTLV)-I/II, herpes simplex virus (HSV)-2, and GB virus C (GBV-C) were also investigated, as they have been associated with a higher CD4+ T cell count,
16
augmented cell susceptibility to HIV-1,
17
and lower HIV-1 loads,
18
respectively. Briefly, HIV-1 coreceptor tropism was inferred from analysis of the env V3 sequence using the geno2pheno online tool (
Related to subtype BF-infected individuals, the group of subtype B infection presented fewer women (p=0.01) and fewer heterosexual men and women (p<0.01). One of the major characteristics of our targeted population is that the majority is constituted of men who have sex with men (MSM). In the city of São Paulo, MSM account for approximately one-third of the overall reported AIDS cases. 15 Other demographic characteristics did not differ. Also, neither host and viral genetic parameters nor frequency of coinfections were differentially represented between the two groups (Table 1).
Fisher's exact test.
At the enrollment visit.
Mann–Whitney U test.
Up to Sep 2010.
SD, standard deviation; ns, not statisticaly significant (p>0.05); MSM, men who have sex with men; HTLV, human T cell lymphotropic virus; HSV, herpes simplex virus; GBV-C, GB virus C.
Next, we focused on the well-established markers for progression of HIV-1 infection: peripheral CD4+ T lymphocyte count and plasma HIV-1 RNA load. CD4+ T cells were measured with the Tritest monoclonal antibodies cocktail and TruCount Tubes using a FACSCalibur flow cytometer (BD Biosciences, San Diego, CA). HIV-1 RNA was quantified by an Amplicor HIV-1 Monitor (Roche, Indianapolis, IN) before January 2007, which was subsequently replaced by the branched DNA assay (Bayer, Tarrytown, NY). The cohort protocol projected blood collections every 4 months and data from absent volunteers were still considered if health and medication information from the period of absence were provided. Clinical visits occurred at a mean±SD interval of 118±87 days, yet considerable individual variation came about (min: 16; max: 830 days). Nine (±6; min: 1; max: 25) measurements of CD4+ T cells and HIV-1 load were compiled for each patient. Data after ART inception were not compiled. Demographic and epidemiological data and baseline CD4+ T cell count (cells/μl) and VL (log10 of copies/ml) were compared by using either Fisher's exact test or Mann–Whitney U tests. Kaplan–Meier survival curves from the date of STAHRS enrollment to the time of reaching ≤350 CD4+ T cells/μl and to the time of starting ART were compared by the Cox proportional hazards model.
Linear regression was used to model the effect of HIV-1 subtype on CD4+ T cell count and plasma HIV-1 RNA load changes, longitudinally. Generalized linear mixed modeling (GLMM) was further executed as it takes the multiple CD4+ T cell and VL measurements for each patient into account (see Supplementary Fig. S1 for additional methodological information about GLMM's execution settings, input data, and results). For both linear regression and GLMM analyses, data from subjects contributing three or more measurements were used (B: n=50; BF: n=7). Models derived a slope (the average change per day in CD4+ T cell count or VL during follow-up) and an intercept (the average CD4+ T cell count or VL change observed at 1 year after the estimated date of infection). Analyses were performed with Prism and IBM SPSS software packages.
Baseline CD4+ T cell count and HIV-1 load were quantified from samples collected at 46±57 and 42±52 days after enrollment, respectively, for subtype B-infected subjects, and at 25±18 and 39±45 days for subtype BF-infected patients. Discordant mean times for baseline CD4+ T cell count and HIV-1 load are explained by the occurrence of two exceptional cases (one B-infected and one BF-infected individual) of independent blood collections for baseline CD4+ T cell counting and for baseline VL quantification. We observed that neither baseline CD4+ T cells nor HIV load diverged between the two groups (Fig. 1B). However, longitudinal analysis of changes in CD4+ T cell count and VL indicated a faster decline of CD4+ T cells in subtype B-infected subjects compared to subtype B-infected subjects, in spite of a similar viral load change (Fig. 1C).
Slope values derived from linear regression models showed a monthly decay of 3.6 CD4+ T cells/μl for subtype B-infected and of 6.3 CD4+ T cells/μl for subtype BF-infected subjects (p<0.01). At the intercept of 1 year after the first CD4+ T cell counting, subtype B-infected individuals had lost 39±5 CD4+ T cells/μl while subtype BF-infected persons had lost 69±16 CD4+ T cells/μl (p=0.01). Such a faster decline of CD4+ T cells among those infected with subtype BF did not appear to be an effect of plasma HIV-1 load, either at set point or over time as slopes for VL change did not differ from zero for both the B-infected and BF-infected groups. Further investigation of CD4+ T cells changes over time was carried out by GLMM analysis assuming the HIV-1 subtype as a fixed effect and testing viral tropism and VL as possible random effects. By this modeling approach, subtype B-infected patients presented a greater ability to preserve their CD4+ T cells over time (days) compared to the subtype BF-infected individuals, by a positive coefficient of 0.34 (95% CI: 0.15–0.52; p<0.001; Supplementary Fig. S1 and Table S1). This notably compares with the results from linear regression analysis, as by GLMM it was calculated that a decay of one CD4+ T cell/μl would take about 3 days longer in subtype B-infected subjects than in subtype BF-infected subjects.
By Kaplan–Meyer survival analysis, we compared the time to reach 350 CD4+ T cells/μl and the time to start ART from the enrollment date. We observed a median time to reach 350 CD4+ T cells/μl of 195 days in subtype BF-infected patients, shorter than the median time of 596 days seen in subtype B-infected subjects [hazard ratio (HR)±95% CI=3.9±1.4 to 11.0; p<0.05]. Analysis of the time to start ART also indicated a faster progression among subtype BF-infected patients, although the difference did not reach statistical significance (median=B: 980 vs. BF: 538 days; HR±95% CI=2.3±0.8 to 6.4; p=0.33; Fig. 1D). We also compared the incidence and the prevalence of cases in need of ART among the 78 studied subjects over time. Both parameters were unfavorable for the subtype BF-infected individuals (data not shown). It is important to note that Brazilian ART guidelines have changed from the observed period of 2002 to 2006, when therapy was started at CD4+ T cell count ≤200 cells/μl, to the present, with patients presenting a CD4+ T cell count ≤500 cells/μl being eligible to start ART.
It is obvious that the unremitting emergence of recombinants of HIV-1 alters the AIDS epidemic. This stresses the need for the characterization of disease presentation in persons infected by these diverse HIV-1 forms. Nonetheless, the association between HIV-1 subtype and disease progression has been difficult to define. Studies intended to categorically analyze such an association must assess data from well-characterized and controlled cases of incident infection, since they happen over large follow-up periods. Moreover, full viral genome sequencing is needed to avoid misclassifying a virus as a pure subtype when it is a recombinant. To our knowledge, our study is the first to compare clinical outcome between infections with subtype B and subtype BF. These HIV-1 forms represent the most prevalent strains in Southeast Brazil, where the AIDS epidemic involves about 300,000 cases.
Here, 78 HIV-1-infected patients were studied, all from a single cohort, and were followed since their recent infection. In addition, their infecting viruses were fully sequenced. This dataset allowed us to reveal a more accelerated rate of CD4+ T cell loss in cases of infection with subtype BF compared to subtype B, in spite of a similar plasma viral load at baseline and over follow-up. This observation provides further evidence that subtype BF might be more pathogenic than subtype B. However, the biological mechanisms involved in this phenomenon remain unclear and could be essentially virological, like coreceptor tropism, transmissibility, or replicative capacity, or could be related to host immunity, such as bearing gp120 molecules more able to bind CD4 or less exposed to antibodies, or carrying immunodominant CTL epitopes that fail to elicit efficient CD8+ T cell-mediated responses. Likewise, elevated virus-mediated cell damage and induction of apoptosis could be acting as a mechanism and this hypothesis has already begun to be investigated by our group.
Lastly, we suggest that our findings and future studies of this nature may add to our understanding of the dynamics and evolution of HIV-1 epidemics, further influencing treatment and vaccine development. It may be conceivable that subtype assignment among recently diagnosed individuals could also help to predict those more likely to present faster disease progression, thus resulting in closer follow-up and/or earlier initiation of antiretroviral therapy.
Footnotes
Acknowledgments
We thank all the study participants and the dedicated clinical research staff at the collaborating sites. This study was supported by the Brazilian Program for STD/AIDS–Ministry of Health (914/BRA/3014 to E.G.K.), the São Paulo City Health Department (2004-0.168.922-7 to E.G.K.), and the Fundação de Amparo à Pesquisa do Estado de São Paulo (2004/15856-9 to E.G.K. and 2010/51609-7 to L.F.T.).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.