
Abstract
The subcellular localization of JAK3 was examined by quantitative image analysis. For the first time, JAK3 was found to be located in the nuclei of primary CD4 lymphocytes. A comparable quantity of JAK3 was recovered in CD4 lymphocytes from healthy donors and HIV-infected patients. By contrast, far more phosphorylated JAK3 (pJAK3) was found in the nuclei of CD4 lymphocytes from HIV-infected patients than from healthy donors. The correlation detected between the quantity of pJAK3 in the nuclei of CD4 lymphocytes and the increase in HLA-DR at their surface suggests that pJAK3 may play a role in the deleterious immune activation characterizing HIV-infected patients.
D
Here, we report on the subcellular distribution of JAK3 in healthy donors and viremic patients. For the first time, we show that JAK3 is located in comparable quantities in the nuclei of primary CD4 lymphocytes from healthy donors and HIV-infected patients. But when we measured phosphorylated JAK3, we found higher quantities in the nuclei of CD4 lymphocytes from HIV-infected patients than from healthy donors, and we then analyzed the correlation between the intensity of this signal and immune activation.
JAK3 subcellular localization was determined by quantitative image analysis on CD4+ T cells from viremic patients and from healthy donors. Peripheral blood mononuclear cells (PBMCs) were prepared and imaged as previously described. 14 Cells were first incubated for 1 h at room temperature with rabbit anti-JAK3 (sc-513, Santa Cruz Biotechnology) or rabbit anti-pY980-JAK3 (sc-16567-R, Santa Cruz Biotechnology) both diluted at 1:200 in staining buffer [phosphate-buffered saline (PBS) 2%, fetal bovine serum (FBS) 0.1%, and Triton X-100 0.1%]. Cells were then washed and incubated with secondary goat antirabbit IgG (H+L)-AF594. As control, anti-JAK3 or anti-pY980-JAK3-specific antibodies were replaced by purified rabbit IgG (dilution 1:200). CD4 lymphocytes were stained with mouse anti-CD4 (BD Pharmingen, diluted at 1:100) followed by goat antimouse IgG (H+L)-AF488 antibodies (Invitrogen). An antibody directed against a protein exclusively located in the cytoplasm, e.g., α-tubulin (sc-8035, Santa Cruz Biotechnology, dilution 1:200), was also used as a control. Coverslips were mounted on glass slides in Vectashield mounting medium with DAPI (Vector Laboratories) to stain nuclei. Images were acquired after optical sectioning on an Apotome system (Zeiss). A script was implemented in Acapella 2.0 software (PerkinElmer) to determine the subcellular location of JAK3. Total cell area was defined as the area enclosed by the CD4-FITC stain. The nucleus was defined as the closed area containing the DAPI stain. JAK3 accumulation in this compartment was defined as the sum of gray values corresponding to the AF594 staining in the nuclear area. All analyses, including cell segmentation and total gray value computation, were carried out automatically on at least 50 CD4+ T cells per slide.
FACS analysis of the CD4 activation markers HLA-DR and CD127 was analyzed to determine the degree of immune activation in the viremic patients. Experiments were carried out on whole blood cells labeled for 30 min at 4°C with an antibody combination consisting of anti-CD3-V500 (BD Biosciences), anti-HLA-DR-PECy7, anti-CD127-APC-AF750 (eBioscience), and anti-CD4-ECD (Beckman Coulter). After red cell lysis and fixation PBMCs were washed and resuspended in PBS before fluorescence acquisition on the same day on a CYAN flow cytometer (Coulter) using SUMMIT V4.3.02 software. Flow cytometry data were analyzed by FlowJo V8.8.4 software (Tree Star).
Statistical analyses were performed using GraphPad Prism 5.0 software (La Jolla, CA) and nonparametric tests in all cases. Differences between subject groups were analyzed by the Mann–Whitney U test. Horizontal bars on data plots indicate median values. Correlations were analyzed using Spearman's rank test. They were considered significant when p values were <0.05.
The viremic patients (VIR) studied had a viral load >10,000 HIV-1 RNA copies/ml plasma, a CD4+ T cell count >100/mm3, and were either antiretroviral treatment naive or had discontinued such treatment at least 6 months previously. The control group (HD, for healthy donors) consisted of anonymous healthy individuals who donated blood at the Etablissement Français du Sang (Paris, France). The study was sponsored by the Agence Nationale de Recherches sur le SIDA et les hépatites virales (ANRS) and was approved by the Ile de France Medical Ethics Committee under number 05–15.
Figure 1a and b shows that JAK3 localizes to the nuclei of CD4+ T cells in both healthy donors and viremic patients. As a control we measured the subcellular distribution of α-tubulin, which was found to be almost exclusively located in the cytoplasm (Fig. 1e). The median fluorescence intensity measured for JAK3 was quantitatively the same in the nuclei of CD4+ T cells from healthy donors and viremic patients, indicating that HIV infection does not affect the nuclear localization of JAK3 (Fig. 2a; p=NS). These results suggest that JAK3 is constitutively located in the nucleus of CD4 T lymphocytes.
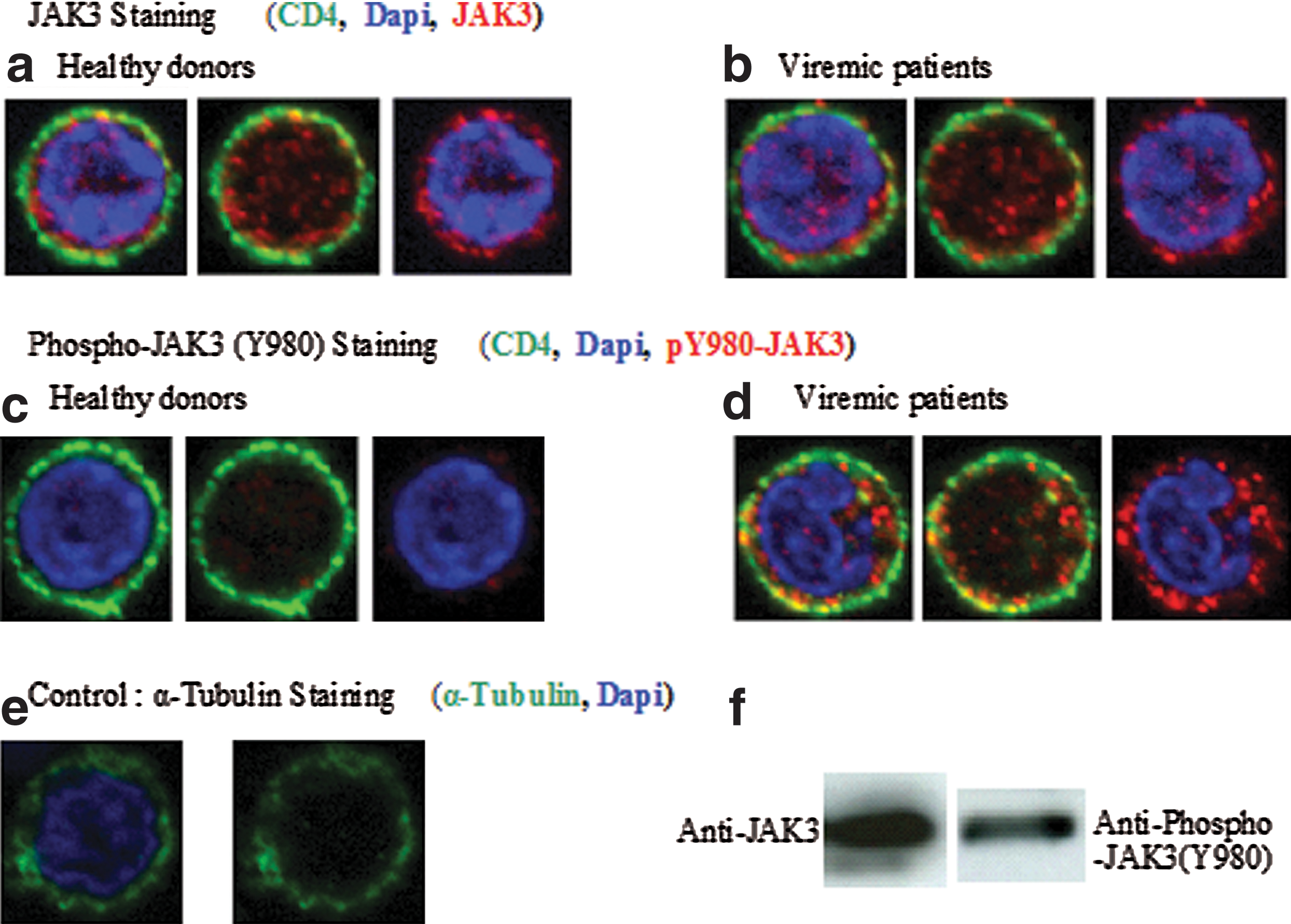
Nuclear localization of JAK3 and pJAK3 in CD4 lymphocytes. Total JAK3 and phosphorylated JAK3 were detected by immunofluorescence in CD4+ T cells. Each cell is represented with and without Dapi nucleus labeling and with and without CD4 labeling to better visualize the presence of JAK3 and pY980-JAK3 within the nucleus and within the cytoplasm, respectively. CD4, green labeling; JAK3, red labeling
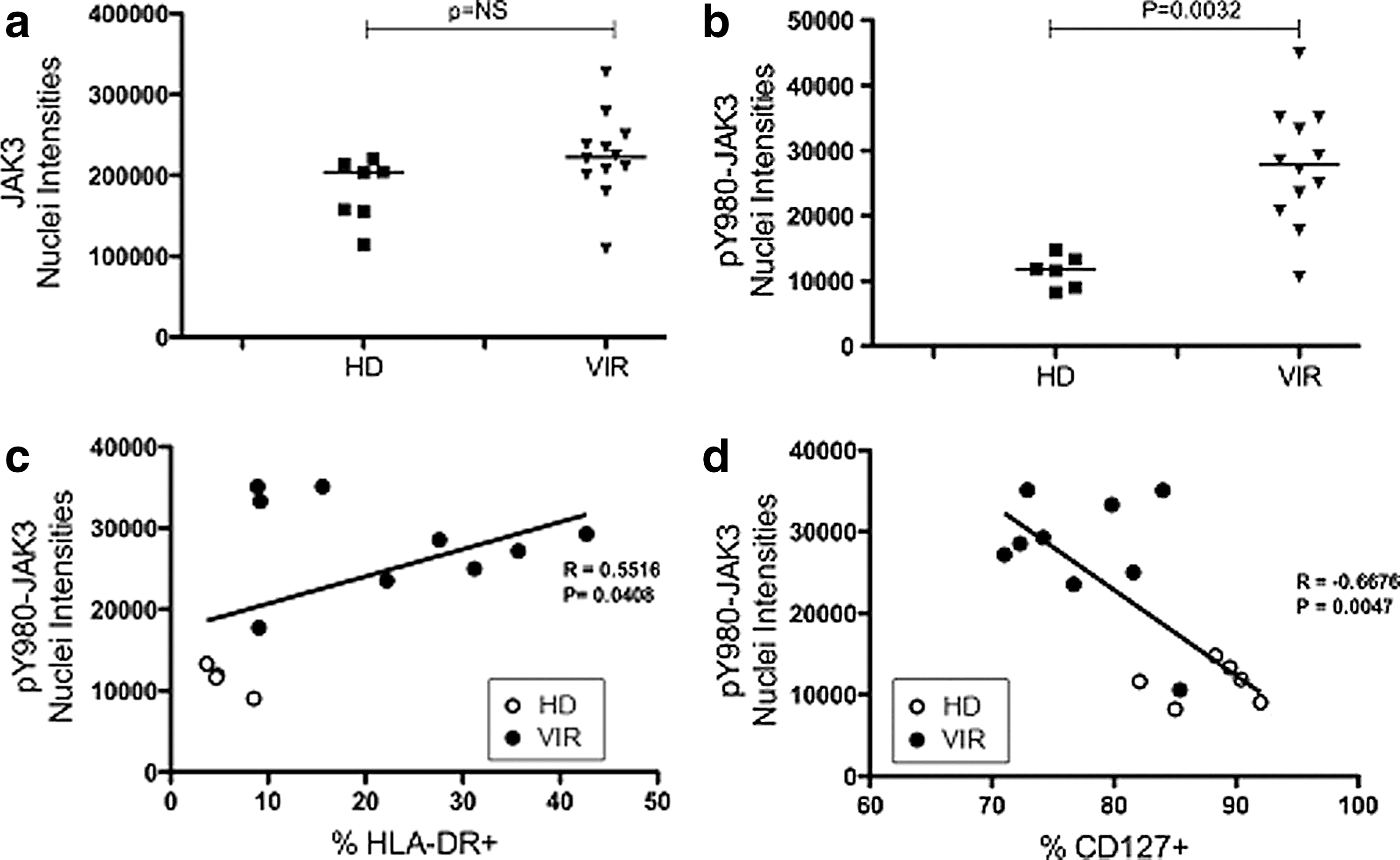
Measurement of JAK3 and pJAK3 in the nucleus and correlation with the activation of CD4 lymphocytes.
JAK3 was found to be hyperphosphorylated in the cytoplasm of CD4+ T cells from viremic patients (Fig. 1c and d). More significantly, fluorescence intensities for pY980-JAK3 were found to be far higher in the nuclei of CD4+ T cells from viremic patients than in those from healthy donors. Corresponding data are analyzed in Fig. 2b (p=0.0032). This conclusively demonstrated that JAK3 phosphorylated at Y980 residue is abundant in the nuclei of CD4+ T cells of viremic patients, and may be involved in the pathogenesis of HIV infection.
Supplementary Fig. S1 (Supplementary Data are available online at
The nuclear fluorescence intensities of pY980-JAK3 measured by quantitative image analysis correlated positively with the percentage of HLA-DR expression at the surface of CD4 T lymphocytes as measured by flow cytometry in viremic patients (Fig. 2c; R=0.5516, p=0.0408). In addition, the amount of active JAK3 in the nuclei of CD4+ T cells was inversely correlated with CD127 expression in viremic patients (Fig. 2d; R=–0.6676, p=0.0047). Since HLA-DR expression and CD127 downregulation are hallmarks of activated CD4+ T cells, these findings demonstrate that increased amounts of pY980-JAK3 in the nuclei of CD4+ T cells are related to the chronic immune activation found in viremic patients.
The putative relationship between nuclear STAT5 and nuclear JAK3 was investigated (see Supplementary Fig. S2). It shows that nuclear tyrosine phosphorylated STAT5 levels correlate negatively with immune activation during HIV infection (Supplementary Fig. S2a). Similarly, the nuclear tyrosine phosphorylated STAT5 level correlates negatively with nuclear phosphorylated JAK3 during HIV infection (Supplementary Fig. S2b).
Our data confirm and extend previous results concerning nuclear localization of molecules of the JAK family. The present study has shown nuclear localization for JAK3. Likewise, other JAK molecules such as JAK1, JAK2, and Tyk2 have also been previously detected in the nuclei of various cell types. 20 –22 In addition, two recent studies have debated the nuclear localization of JAK2. 23,24 JAK2 molecules have been reported to have a nuclear function 21 in that they specifically phosphorylate histone H3 at tyrosine 41 (H3Y41), enabling HP1α to be excluded from heterochromatin. Since H3Y41 phosphorylation occurs in the first H3 helix, in a region known to control nucleosome remodeling, this may have a profound effect on the expression of some normal or abnormal genetic programs related to CD4 lymphocyte activation.
In a recently published article it has been indicated that JAK3 localizes to the nuclei of different cell lines. 25 Moreover, this is the first time that the expression of JAK3 has been characterized in the nuclei of primary cells, and this was achieved by quantitative imaging of CD4 lymphocytes, using a specific antibody against JAK3 (Fig. 1f). This critical observation was further confirmed by using another antibody for the detection of phosphorylated JAK3 in the nuclei of activated cells from HIV-infected individuals. Interestingly, whereas the total quantity of nuclear JAK3 noted in these individuals was the same as in healthy individuals, their nuclear phosphorylated JAK3 content was far higher, thus confirming the presence of an activation process in HIV-infected individuals. It is very unlikely that this process is due to an increase in IL-7 in the plasma of viremic patients 26 as the mean IL-7 concentration found in these patients is around 10 pg/ml, far below the K d of the IL-7 receptor. 27
We have previously reported a regulatory dysfunction of the JAK/STAT pathway in HIV-infected patients. 2,3,13,14 Most of these defects are detected after IL-2 or IL-7 stimulation, explaining decreased responses to these cytokines. 1 –6 More recently we have shown poor nuclear localization of phosphorylated STAT5 in HIV-infected individuals, a defect negatively correlated with the levels of immune activation. 13,14 Therefore, under those conditions, phosphorylated STAT5 cannot directly participate in the mechanisms leading to CD4+ T cell activation. By contrast, the correlation reported here between the recovery of phosphorylated JAK3 in the nucleus and membrane over expression of HLA-DR or down modulation of CD127 points to a new and compensatory mechanism that may play a role in the induction of the abnormal activation of the CD4 compartment observed in HIV-infected patients. It was recently proposed that nuclear JAK1 and JAK2 28 may activate some genes either directly as transcription factors or by opening heterochromatin and allowing access to known transcription factors already present in the nucleus. 29 It may therefore be imagined that nuclear hyperphosphorylated JAK3 increases the function of certain promoters, thereby enhancing genetic programs related to deleterious activation of the immune system from HIV-infected patients.
This report further emphasizes the role of the JAK/STAT pathway in HIV pathogenesis. Further studies are now required to decipher the role of nuclear hyperphosphorylated JAK3 in the abnormal activation of CD4 lymphocytes from HIV-infected patients.
Footnotes
Acknowledgments
We wish to thank Dr. Lisa Chakrabarti (Unité d'Immunogénétique Cellulaire, Institut Pasteur) and Prof. Olivier Lambotte and Jean-François Delfraissy (Assistance Publique Hôpitaux de Paris, Centre Bicêtre) as PIs from the ANRS EP33-2 study. We also wish to thank Huguette Berthé and Thomas L'Hyavanc for their help in recruiting patients and Alain Cosson for advice and reagents. We are especially grateful to the patients who took part in the study.
I.L. was supported by a fellowship from the Panamanian Government (Senacyt). This work was part of study EP33-2, which was supported by the Agence Nationale de Recherche sur le SIDA et les Hépatites Virales. I.L. designed and performed the experiments with technical support from Florence Bugault for the FACS analysis. V.N.S analyzed data and gave statistical support. J.T. analyzed the data and wrote the paper.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.