
Abstract
Toll-like receptor 9 (TLR9) is one of the key sensors that recognize viral infection/replication in the host cells. Studies have demonstrated that methamphetamine (METH) dysregulated host cell innate immunity and facilitated HIV infection of macrophages. In this study, we present new evidence that METH suppressed TLR9-mediated anti-HIV activity in macrophages. Activation of TLR9 by its agonist CpG-ODN 2216 inhibits HIV replication, which was demonstrated by increased expression of TLR9, interferon (IFN)-α, IFN regulatory factor-7 (IRF-7), myeloid differentiation factor 88 (MyD88), and myxovirus resistance gene A (MxA) in macrophages. However, METH treatment of macrophages greatly compromised the TLR9 signaling-mediated anti-HIV effect and inhibited the expression of TLR9 downstream signaling factors. Dopamine D1 receptor (D1R) antagonists (SCH23390) could block METH-mediated inhibition of anti-HIV activity of TLR9 signaling. Investigation of the underlying mechanisms of the METH action showed that METH treatment selectively down-regulated the expression of TLR9 on macrophages, whereas it had little effect on the expression of other TLRs. Collectively, our results provide further evidence that METH suppresses host cell innate immunity against HIV infection by down-regulating TLR9 expression and its signaling-mediated antiviral effect in macrophages.
Introduction
M
Toll-like receptor 9 (TLR9) is a primary receptor for CpG DNA and synthetic CpG-containing oligonucleotides (CpG-ODN). 10 When class A CpG-ODN binds to TLR9, it stimulates the secretion of interferon (IFN)-α, 11,12 a potent antiviral cytokine that impedes HIV infection/replication in macrophages. 13 –15 Studies on TLR9 16 have revealed that postexposure treatment with CpG-ODN has an immunostimulatory effect on retrovirus-induced disease. Therefore, it has been proposed that CpG-ODN may constitute a treatment regime to restrain HIV infection/replication, as it can reactivate HIV-1/AIDS patients' immune response. 17,18 Studies have shown that METH dysregulated host cell innate immunity and facilitated HIV infection. Thus, we investigated whether METH treatment impairs TLR9-mediated anti-HIV activity in macrophages.
Materials and Methods
Monocyte isolation and culture
Peripheral blood samples from healthy adult donors were provided by the University of Pennsylvania Center for AIDS Research, which has Institutional Review Board review and approval for the sample collection. These blood samples were screened for all normal viral blood-borne pathogens and were certified to be pathogen free. To avoid donor variation, we pooled the blood samples from three donors for each experiment, and all experiments were independently repeated at least three times. Monocytes were purified according to a previously described technique. 19
In brief, heparinized blood was separated by centrifugation over lymphocyte separation medium (Organon Teknika, Durham, NC) at 400–500×g for 45 min. The mononuclear cell layer was collected and incubated with Dulbecco's modified Eagle's medium (DMEM) (Invitrogen, Carlsbad, CA) in a 2% gelatin-coated flask for 45 min at 37°C, followed by removal of the nonadherent cells with DMEM. Adherent monocytes were detached with 10 mM EDTA. After the initial purification, greater than 97% of the cells were monocytes, as determined by nonspecific esterase staining and flow cytometry analysis using monoclonal antibody against CD14, the marker specific for monocytes and macrophages. Isolated monocytes were plated in 24- or 48-well culture plates at a density of 5 or 2.5×105 cells/well, respectively, in DMEM containing 10% fetal calf serum (10% DMEM).
In this study, monocytes refer to freshly isolated monocytes (within 24 h), and macrophages refer to 7-day-cultured monocytes in vitro. The viabilities of monocytes and macrophages were monitored by Trypan blue exclusion and maintenance of cell adherence.
Reagents
METH and SCH 23390, a dopamine D1 receptor (D1R) antagonist, were purchased from Sigma-Aldrich Co. (St. Louis, MO). ODN 2216, a specific TLR9 agonist, and its control pCpG Giant were purchased from InvivoGen (San Diego, CA). TLR3 ligand poly(I:C) HMW, TLR4 ligand LPS-EB, and TLR7/8 ligand R848 were also purchased from InvivoGen (San Diego, CA). The purified monoclonal TLR9 (eB72-1665) antibodies were purchased from Imgenex (San Diego, CA). Fluorescein isothiocyanate-conjugated goat antirabbit IgG antibody was purchased from Southern Biotechnology Associates, Inc. (Birmingham, AL).
METH and/or TLR ligand treatment
Seven-day-cultured macrophages (2.5×105 cells/well) were pretreated for 3, 6, and 24 h with different concentrations (1, 10, 100, and 250 μM) of METH, which are comparable to the levels found in the blood, urine, or tissue samples of METH-using subjects. 8,20,21 Various TLR ligands, including ODN 2216 (0.5 μM), poly(I:C) (2 μg/ml), LPS (100 ng/ml), and R848 (2 μg/ml) were added to macrophage cultures 1 h before METH treatment, respectively. The cell cultures were refed with fresh 10% DMEM containing METH and various TLR ligands every 4 days. To investigate whether D1R is involved in METH-mediated down-regulation of CpG-ODN's anti-HIV activity in macrophages, 10 μM SCH 23390, a D1R antagonist, was added to the macrophage cultures 1 h before METH treatment for 24 h. ODN 2216 (0.5 μM) was added to macrophage cultures 6 h after METH treatment and the treatment was maintained for 24 h. The cell cultures were refed with fresh media containing METH, ODN 2216, and/or SCH 23390 every 4 days. There were no cytotoxic effects of METH, ODN 2216, poly(I:C), LPS, R848, and SCH 23390 on macrophages at the concentrations used in this study, as demonstrated by Trypan blue dye staining (data not shown).
Infection of macrophages with HIV Bal strain
The macrophage-tropic R5 strain (Bal) was obtained from the AIDS Research and Reference Reagent Program (NIH, Bethesda, MD). Macrophages were infected with equal amounts of HIV Bal virus (p24 20 ng/106 cells) for 2 h at 37°C after treatment with or without various TLR ligands. The cells were then washed three times with plain DMEM to remove unabsorbed virus, and fresh media containing METH, ODN 2216, and/or SCH 23390 were added to the cell cultures. Culture supernatants were collected for HIV reverse transcriptase (RT) activity assay on days 8 and 12 after infection. Untreated cells served as a control. The cells were observed and photographed under a light microscopy (×200) on day 8 postinfection.
HIV RT assay
HIV RT activity was determined based on the technique of Guo et al. 22 and Ho et al. 23 with modifications. In brief, 10 μl of culture supernatants from macrophages infected with or without HIV was added to a cocktail containing poly(A), oligo(dT) (Amersham Biosciences, Inc., Piscataway, NJ), MgCl2, and [ 32 P]dTTP (Amersham Biosciences, Inc.) and incubated for 20 h at 37°C. Then 30 μl of the cocktail was spotted onto DE81 paper, dried, and washed five times with 2×saline-sodium citrate buffer and once with 95% ethanol. The filter paper was then air-dried. Radioactivity was counted in a liquid scintillation counter (PerkinElmer Life Sciences, Boston, MA).
Real-time RT-PCR
Total RNA was extracted from macrophages using Tri-Reagent (Molecular Research Center, Cincinnati, OH) as previously described. 24 Total cellular RNA (1 μg) was subjected to reverse transcription using the reverse transcription system from Promega (Madison, WI). Real time reverse transcriptase polymerase chain reaction (RT-PCR) was used to quantify mRNA expression of TLR1-10, IFN-α, MyD88, IRF7, MxA, and ISG56 that relate to the TLR9 signaling pathway, as well as glyceraldehyde-3-phosphate dehydrogenase (GAPDH), with the iQ SYBR Green Supermix (Bio-Rad Laboratories, Hercules, CA) as previously described. 24 The amplified products were visualized and analyzed using the MyiQ software provided with the thermocycler (iCycler iQ real time PCR detection system; Bio-Rad Laboratories). The levels of GAPDH mRNA were used as an endogenous reference to normalize the quantities of target mRNA. The special oligonucleotide primers used in this study were synthesized by Integrated DNA Technologies, Inc. (Coralville, IA). Sequences of these primers are listed in Table 1.
Immunofluorescence assay
The macrophages were cultured on glass coverslips at a density of 0.25×106/well in 24-well plates. The macrophages were washed with 1×cold phosphate-buffered saline (PBS) (with Ca2+ and Mg2+) twice. Cells were fixed at 4°C in 4% paraformaldehyde–4% sucrose in PBS for 20 min and then permeated in cold methanol (100%) for an additional 10 min followed by 0.2% Triton X-100 for an additional 10 min. Cells were blocked in Block Solution (Pierce, Rockford, IL) for 1 h at room temperature. Mouse purified monoclonal antibody (1:100) against TLR9 was used as the primary antibody. After washing five times with 1×PBS, the cells were incubated with fluorescein isothiocyanate-conjugated goat antimouse IgG antibody (green, 1:500) for 1 h and rewashed five times with 1×PBS. Then the cells were mounted on glass coverslips in mounting media (Biomeda, Foster City, CA) and viewed by fluorescence microscopy (Zeiss, Jena, Germany). Hoechst 33342 was used for nuclei morphology.
Enzyme-linked immunosorbent assay
Total cell lysates from the cultured macrophages were prepared using a radioimmune precipitation assay buffer (Promega). Enzyme-linked immunosorbent assay for IFN-α was performed according to the protocol provided by the manufacturer (PBL Biomedical Laboratories).
Statistical analysis
Student's t-test was used to evaluate the significance of difference between groups, and multiple comparisons were performed by one-way analysis of variance. p values of less than 0.05 were considered significant. Data are presented as the mean±SD. Statistical analyses were performed with SPSS 11.5 for Windows.
Results
METH inhibited CpG-ODN's anti-HIV activity in macrophages
As shown in Fig. 1A, METH treatment of macrophages resulted in an increase in HIV RT activity, whereas ODN 2216 treatment inhibited HIV RT activity. However, the anti-HIV effect of ODN 2216 is inhibited when macrophages were pretreated with METH, and the highest inhibition was observed at day 8 postinfection (Fig. 1A). In addition to TLR9 ligand ODN 2216, the ligands for other TLR (TLR3, TLR4, and TLR 7/8)-related HIV recognition were also demonstrated to have an inhibitory effect on HIV replication, which was evidenced by significantly decreased HIV RT activity in macrophage cultures treated with these TLR ligands (Fig. 1B). The pretreatment of macrophages with METH, however, had little effect on the HIV inhibitory activity of these ligands (Fig. 1B). Morphologically, characteristic giant syncytia were observed in untreated and METH-treated HIV Bal-infected macrophage cultures. In contrast, ODN 2216-treated macrophages failed to develop HIV-induced giant syncytia (Fig. 1C).
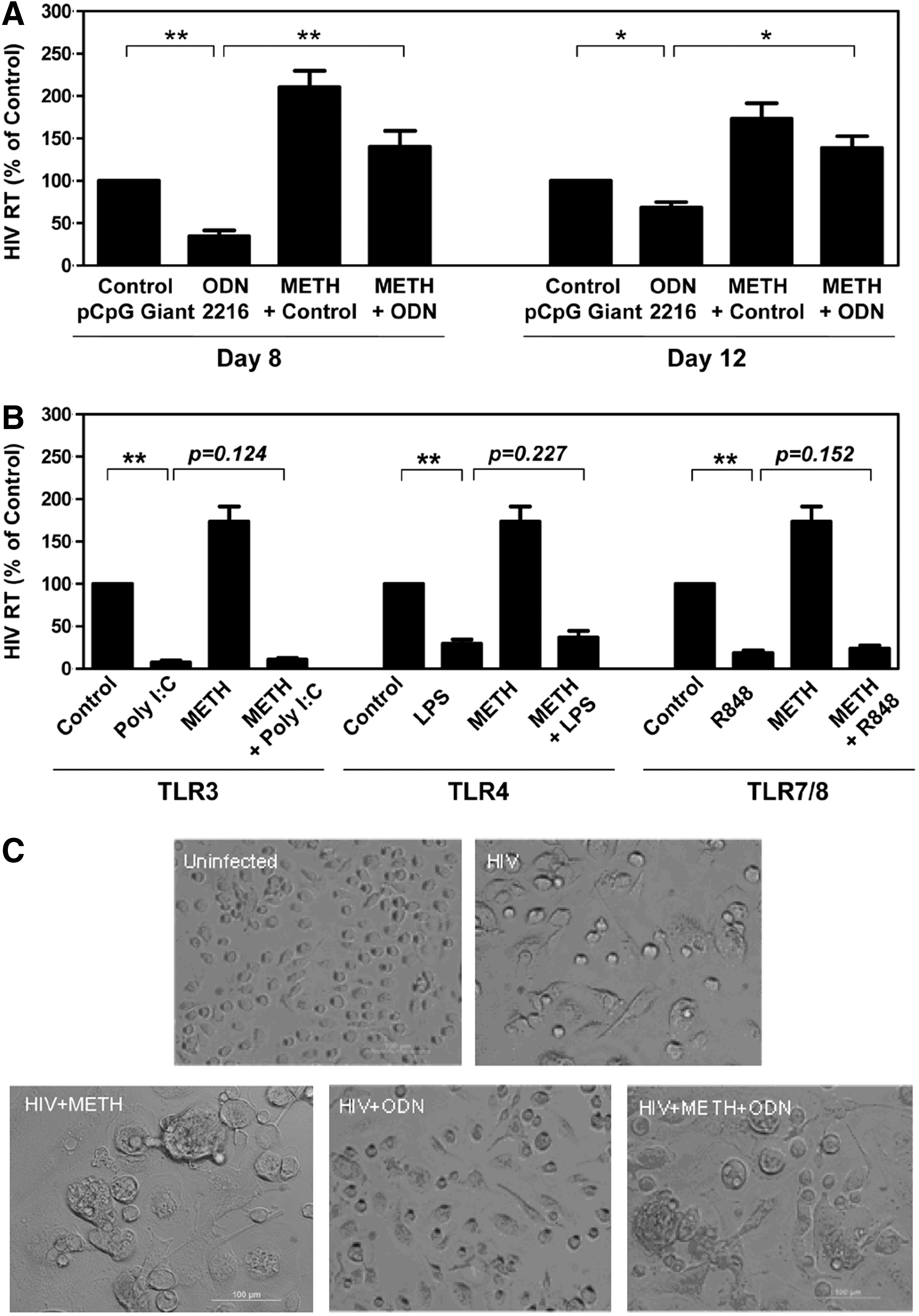
Methamphetamine (METH) compromises ODN-mediated inhibition of HIV Bal infection.
D1R was involved in METH-mediated down-regulation of CpG-ODN's anti-HIV activity in macrophages
D1R has been shown to be implicated in the pathological effect of METH on the target cells. 9 Thus, it is of importance to determine whether METH-mediated dysregulation of CpG-ODN activity is through D1R. The D1R antagonist (SCH23390) could abrogate the inhibitory effect of METH on ODN 2216-mediated HIV inhibition (Fig. 2).

Dopamine D1 receptor (D1R) antagonist blocks the inhibitory effect of METH on CpG-ODN's anti-HIV activity in macrophages. Seven-day-cultured macrophages were incubated with or without 10 μM SCH 23390 for 1 h, followed by 100 μM METH for 24 h, then 0.5 μM ODN 2216 for 6 h, and then infected with the HIV Bal strain. Culture supernatants were collected on day 8 postinfection for HIV RT assay. HIV RT activity was expressed as a percentage of HIV RT activity in cells without METH, ODN, and SCH treatment, which was defined as 100% (the first bar on the histogram). The results shown in this figure are of the three independent experiments, being represented as the mean±SD. Statistical analysis was performed using one-way analysis of variance, and significance was shown with **p<0.01.
METH selectively suppressed the expression of TLR9
To further determine potential mechanism(s) involved in METH-mediated inhibition of host cell innate immunity, we examined the impact of METH on TLR1-10 expression in macrophages. As shown in Fig. 3A, METH treatment of macrophages had little effect on the expression of TLRs 1–8 and TLR 10 (Fig. 3A). However, METH significantly suppressed the expression of TLR9 (Fig. 3A–C). This inhibiting effect of METH on TLR9 was dose and time dependent (Fig. 3B and C). The highest inhibitory effect by METH was observed with a concentration of 250 μM at 24 h of exposure (Fig. 3B and C).
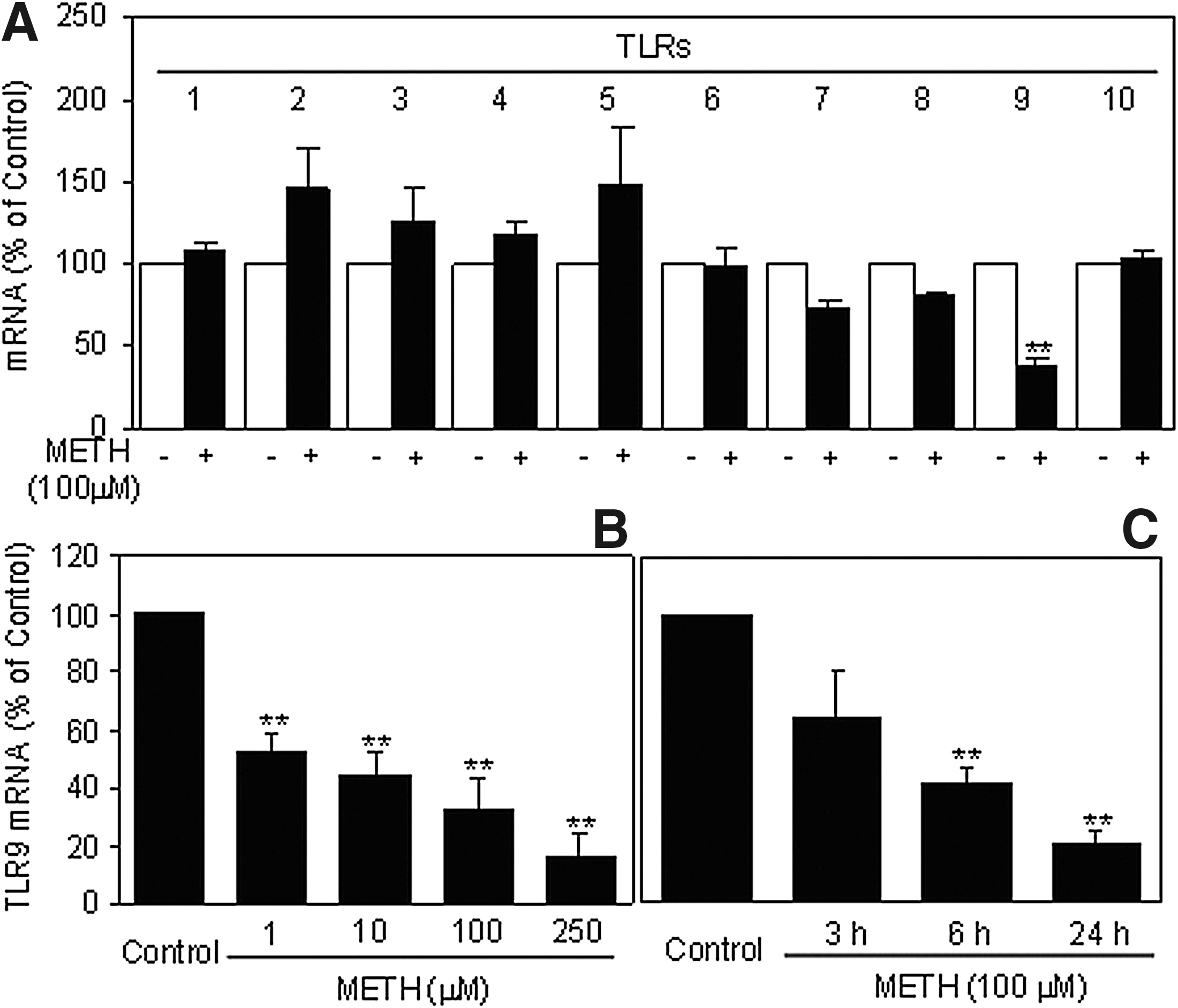
Effects of METH on Toll-like receptor (TLR) expressions in macrophages. Seven-day-cultured macrophages were incubated with METH at indicated concentrations for 24 h
METH reduced CpG-ODN-stimulated expression of TLR9 and IFN-α, which was blocked by the D1R antagonist
We observed that ODN 2216 treatment could induce TLR9 expression at both mRNA and protein levels (Fig. 4). However, when macrophages were pretreated with METH, ODN 2216-induced TLR9 expression was inhibited (Fig. 4). In addition, METH inhibited ODN 2216-mediated IFN-α expression (Fig. 5). However, the pretreatment of macrophages with the D1R antagonist (SCH23390) could prevent METH-mediated inhibition of TLR9 and IFN-α expression (Figs. 4 and 5).
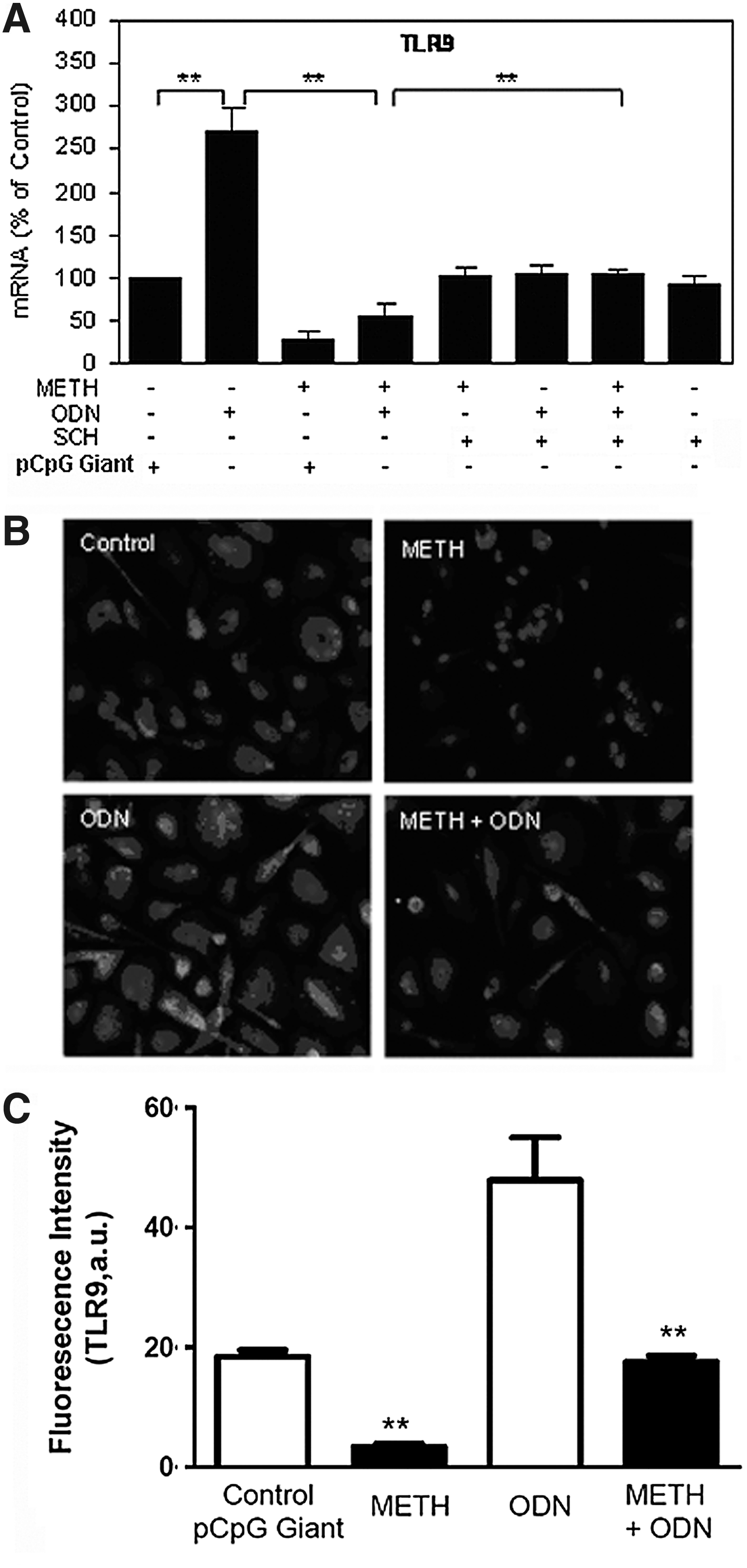
METH compromises ODN-mediated up-regulation of TLR-9.
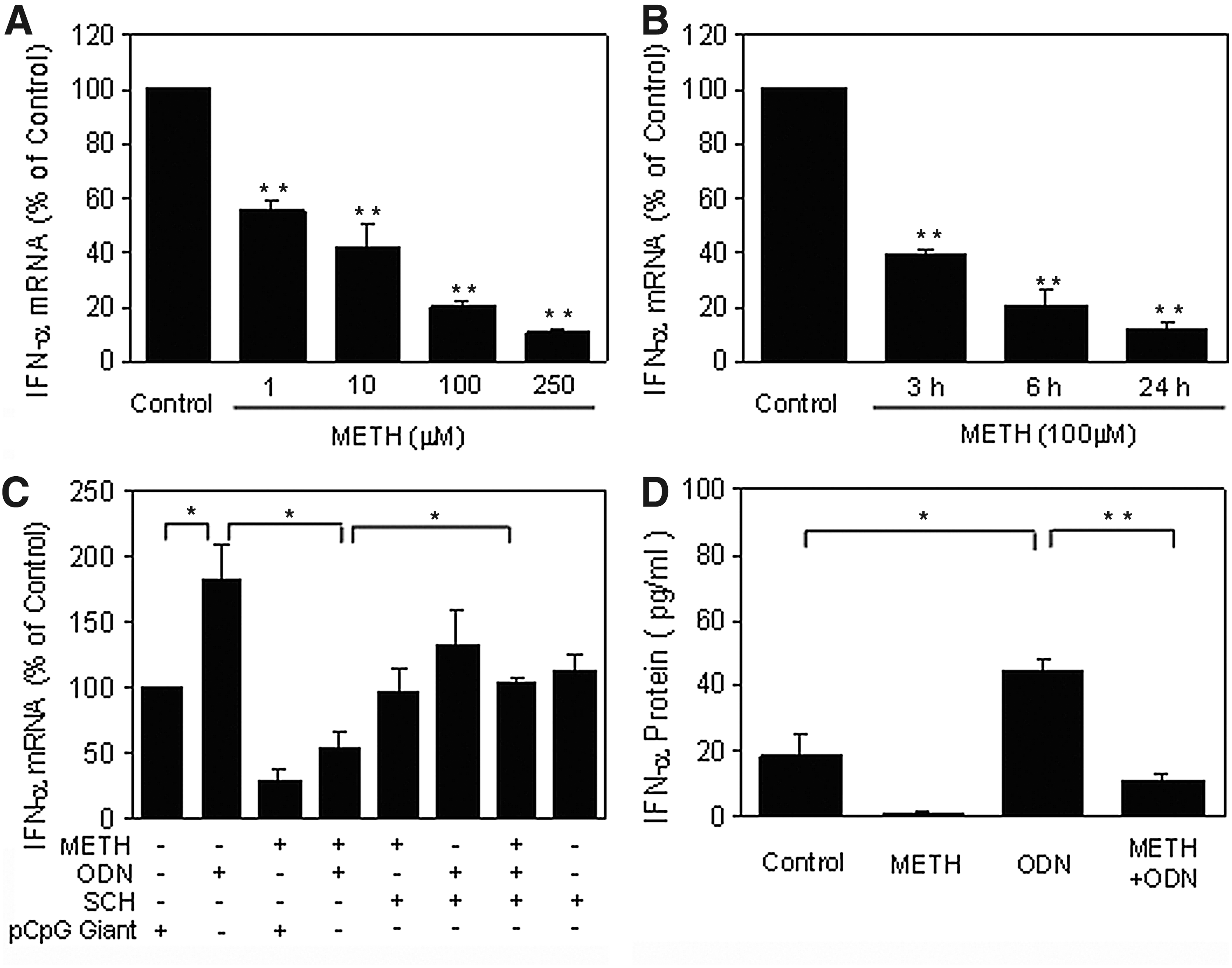
METH compromises ODN-mediated up-regulation of interferon (IFN)-α.
METH inhibited the expressions of TLR9 downstream signaling molecules in macrophages
As shown in Fig. 6, ODN 2216 induced higher levels of IRF-7, MyD88, and MxA, but did not significantly induce ISG56 expression. However, induction of IRF-7, MyD88, MxA, and ISG56 was inhibited in macrophages treated with METH prior to ODN 2216 stimulation. The D1R antagonist SCH 23390 could counteract the inhibitory effects of METH on the expression of MyD88 and ISG56, but not on IRF-7 and MxA (Fig. 6).

METH compromises ODN-mediated up-regulation of TLR9 downstream signaling molecules. Seven-day-cultured macrophages were treated with or without 10 μM SCH 23390 for 1 h, followed by 100 μM METH for 24 h, then 0.5 μM ODN 2216 for 6 h. Cellular RNA was subjected to real-time RT-PCR for IRF-7 mRNA
Discussion
Synthetic ODNs are generated based on the structural properties of the physiological ligands of TLR9, bacterial DNA rich in CpG motifs. 10 Because of its ability to activate TLR9, inducing antiviral factors, 25 CpG-ODN has been shown to suppress HIV replication either in vivo 26 or in clinical studies. 27 TLR9 could activate the type I IFN pathway, 28,29 which is critical for the control of viral infection/replication, including HIV. Clinically, higher HIV loads in plasma and brain were found in METH users. 30,31 We and others have shown that METH treatment of macrophages enhances HIV infection. 5,9 The investigation of the underlying mechanisms of METH action showed that METH inhibits the expression of endogenous IFN-α and STAT1 in macrophages. 9
To further determine the mechanisms involved in METH-mediated HIV enhancement, the present study investigated the impact of METH on the TLR9 signaling pathway in macrophages. We showed that METH compromised the anti-HIV activity of ODN 2216, as a strong inducer of IFN-α 32 and a potent inhibitor of HIV infection of macrophages. 13 In addition, we observed that METH treatment of macrophages inhibited the expression of TLR9 (Fig. 3). This inhibitory effect of METH on TLR9 is specific, as other TLRs tested were not affected by METH in macrophages (Fig. 3A). This finding provides an additional mechanism for METH-mediated impairment of intracellular antiviral signaling pathways.
The selective action of METH on TLR9 is interesting. However, we are still investigating the mechanisms involved in the action of METH on TLR9. The different cellular and subcellular locations may contribute to the selective effect of METH on TLR9. It is well known that TLR4 is localized on the outer cell surface membrane, while TLR9 is an intracellular receptor, which is positioned on endosomes or lysosomes. 33 In addition, although TLR3, TLR7, TLR8, and TLR9 that recognize foreign nucleic acid are all intracellular receptors and are mainly positioned on endosomes, the mechanisms that dictate their location are different. 33 The intracellular location of TLR3 is determined by a 23-amino acid sequence present in the linker region between the transmembrane and TIR domain. 34 TLR7 has been shown to be colocalized with TLR3-containing compartments, however, intracellular localization of TLR7 is defined by its transmembrane domain. 34 To be different from TLR3 and TLR7, a tyrosine-based motif in the cytoplasmic domain of human TLR9 dictates its localization. 35 These findings indicate that the subcellular compartmentalization of TLR9 on endosome is distinct from TLR3 and TLR7/8, which may result in different regulatory mechanisms to these TLRs, including TLR expression, trafficking, proteolysis, and autophagy.
Our further investigation showed that METH treatment of macrophages could suppress the expression of IRF7 and MyD88. The IRFs are important regulators in type I IFN-mediated antiviral immunity. These regulators not only recognize the elements of the DNA promoter to promote IFN gene expression, but also regulate the IFN-stimulated response element (ISRE) in some of ISGs, leading to induction of an antiviral state. 36 Among IRFs, IRF7 can activate IFN-α-dependent promoters. 37,38 IRF7 interacts with MyD88 to form a complex, which provides the foundation for the induction of IFN-α. 39 More importantly, TLR9 signaling-mediated IFN-α induction is shown to be entirely dependent on MyD88. 40 Therefore, the down-regulation of IRF7 and MyD88 by METH provides the justification for the action of METH in facilitating HIV replication in macrophages.
As one of the key sensors that recognize viruses and viral products, activating the type I IFN pathway, TLR9 signaling can induce the production of a number of antiviral factors, including MxA protein, a key mediator of type I IFN-medicated antiviral response to a wide range of RNA viruses and some DNA viruses as well. 41,42 Clinically, elevated levels of MxA have been observed in whole blood lysates from patients with viral infections, including HIV. 43,44 We found that ODN 2216 treatment of macrophages enhanced MxA expression. This effect of ODN 2216, however, was compromised by METH (Figs. 1A and 2). In this study, we found that ISG56 could not be induced by ODN 2216, which is another indicator of IFN-α. 45,46 One possible reason may be that the levels of IFN-α induced by CpG ODN under our study conditions were relatively low. In this study, low concentrations of CpG ODN were used to avoid higher concentrations of CpG ODN masking the inhibitory effect of METH.
Collectively, our study provides compelling experimental evidence that METH inhibits TLR9-mediated anti-HIV activity in macrophages by blocking the TLR9-MyD88-IRF7-IFN-α signaling pathway. These additional observations indicate that METH use impairs intracellular innate antiviral mechanisms in macrophages, contributing to cell susceptibility to HIV infection. Because METH exerts profound and detrimental effects on host cell innate immunity, which is critical in restricting HIV replication, it is likely that METH use is a cofactor in facilitating HIV disease progression. However, clinical studies are required not only to validate the in vitro observations but also to design innate immunity-based intervention and treatment strategies for METH users with HIV infection.
Footnotes
Acknowledgments
We are grateful for financial support from the U.S. National Institutes of Health (NIH; grants DA012815 and DA027550, to Wen-Zhe Ho) and the Guangxi Natural Science Foundation (2010GXNSFD013045, to Hao Liang).
Author Disclosure Statement
No competing financial interests exist.