
Abstract
The relationship between exogenous contraceptive hormones and permissiveness of the female genital tract to human immunodeficiency virus type 1 (HIV-1) is the subject of renewed debate. To better characterize the effect of depot medroxyprogesterone acetate (DMPA) on HIV-1 cellular targets and epithelial integrity in the vagina, we compared leukocyte populations, markers of activation and proliferation, and the density of intercellular junctional proteins in the vaginal epithelium of women during the follicular and luteal phases of the menstrual cycle and approximately 12 weeks after receiving a DMPA injection. This prospective cohort study involved 15 healthy women. Vaginal biopsies were obtained in the follicular and luteal phases of the menstrual cycle, and approximately 12 weeks following a 150-mg intramuscular injection of DMPA. Leukocyte populations, activation phenotype, and epithelial tight junction and adherens proteins were evaluated by immunohistochemistry. After receiving DMPA, the numbers of CD45, CD3, CD8, CD68, HLA-DR, and CCR5 bearing immune cells were significantly (p<0.05) increased in vaginal tissues, compared to the follicular and/or luteal phases of untreated cycles. There were no significant differences in immune cell populations between the follicular and luteal phases of the control cycle. There were also no statistically significant differences in epithelial thickness and density of epithelial tight junction and adherens proteins among the follicular, luteal, and post-DMPA treatment sampling points. In this pilot study, vaginal immune cell populations were significantly altered by exogenous progesterone, resulting in increased numbers of T cells, macrophages, and HLA-DR- and CCR5-positive cells.
Introduction
D
Therefore, elucidating the interaction between exogenous hormones and mucosal susceptibility to HIV-1 is critical, as the main route of HIV-1 infection in women is via heterosexual intercourse. 10 Several large epidemiologic studies in Africa and Thailand have found that women using DMPA were significantly more likely to acquire HIV-1 as compared to women not using hormonal contraception, even after controlling for certain confounding variables such as frequency of intercourse, age, and number of sexual partners. 8,11 –14 As is the case with most observational studies, however, not all methodological considerations, e.g., consistent condom use or other important, unknown confounders, could be properly controlled. 15
Theories as to how exogenous progesterone might increase genital tissue susceptibility to HIV-1 infection center on epithelial thinning and alteration of local mucosal immunity. A decrease in the number of epithelial cell layers or the density of intercellular junction proteins potentially enhances exposure of cervicovaginal mucosal target cells to HIV-1. In nonhuman primate models of HIV infection, progesterone and DMPA administration cause a dramatic atrophy of the vaginal epithelium, and DMPA (at 30 mg) is routinely used to enhance the efficiency of vaginal infection by simian immunodeficiency virus (SIV) or a chimeric variant of HIV (SHIV). 16 –21 However, data regarding the effect of exogenous progestins on epithelial thickness in the human vagina are mixed, ranging from no change to either increased or decreased thickness, 5,22 –24 in short-term (3–6 months) 5,22 and long-term (2–3 year) 23 DMPA users.
DMPA could also affect mucosal susceptibility to HIV-1 infection through alterations of the cervicovaginal mucosal immune response. Although the cervix and vagina are likely the initial site of entry of HIV-1 in women, the effects of exogenous estrogen and progesterone on the local immune environment of the lower genital tract have not been clearly elucidated, 24 –29 with most data focusing on biological mechanisms within the endometrium. 30
The objective of this study was to compare immune cell populations and markers of epithelial integrity in women during the normal menstrual cycle and 3 months after receiving a single injection of DMPA, to determine if exogenous and endogenous progesterone alters the cellular immunoinflammatory status of the vaginal mucosa. To this end, we examined vaginal leukocytes bearing CD45, CD3, CD4, CD8, CD1a, and CD68 antigens, markers of activation and proliferation such as HLA-DR, CCR5, and Ki-67, and the epithelial junctional proteins zona occludin 1 (ZO-1) and E-cadherin. Because the dual need for contraception and protection from HIV-1 exists for so many women, especially in high-risk areas such as sub-Saharan Africa, the possible biological mechanisms underpinning a relationship between effective contraceptives such as DMPA and enhanced HIV-1 susceptibility need to be urgently explored.
Materials and Methods
Clinical study
This study was approved by the Institutional Review Board of Eastern Virginia Medical School. Details of the clinical study have been previously described. 5 In brief, 20 women were screened for the study. After verifying the absence of exclusionary factors, such as sexually transmitted infections (STIs), women were examined in the luteal phase of the menstrual cycle (days 22–26), the follicular phase of the menstrual cycle (days 8–12), and finally 12 weeks (range 81–96 days) after receiving an injection of 150 mg of DMPA intramuscularly. Phase of the menstrual cycle was verified by menstrual calendar and serum estradiol and progesterone measurements. At each visit, two vaginal biopsies, colposcopy, swabs of cervicovaginal secretions, and serum samples for estradiol, progesterone, and MPA were obtained. All women enrolled in this study were otherwise not at risk of pregnancy due to female tubal sterilization or had a male sexual partner with a vasectomy and were not using any exogenous hormones during the study. 5
Tissue analysis
Paraffin-embedded tissue blocks of the vaginal biopsies were cut into 5-μm sections. Immunohistochemistry (IHC) staining of tissue sections was performed using the ABC method [avidin:biotinylated enzyme complex from Vector labs (Burlingame, CA)]. Briefly, the slides were deparaffinized, dehydrated, and rehydrated followed by antigen retrieval in citrate buffer (pH 6.2, DAKO) at high temperature. Nonspecific binding was blocked using specific protein block for 30 min at room temperature. After washing with phosphate-buffered saline (PBS), the slides were incubated overnight with primary antibody at 4°C. Slides were then washed with PBS and subjected to biotinylated secondary antibody followed by ABC reagent. The antigen was localized by incubation with AEC chromogen–substrate (skyTek Labs, Mississauga, Ontario, Canada) and finally mounted with Accergyl mounting media (Accurate Chemicals, NY) with a cover slip. Positive stained cells were either counted under the microscope (Nikon E-800) or analyzed using Image J software (NIH, Bethesda, MD), as described below.
The main endpoint of this secondary analysis was the number of cell types associated with mucosal immunity and inflammatory responses and the density of epithelial junctional proteins (ZO-1 and E-cadherin) in the vaginal tissue biopsies. Cell phenotype was identified using specific unconjugated monoclonal antibodies against CD1a, CD3, CD4, CD8, CD45, CD68, CCL5, HLA-DR, and Ki67 markers and cell density was expressed per square millimeter of tissue (cells/mm2). The following mouse monoclonal antibodies were used: CD45 (X16/99 clone, Leica Microsystems), CD3 (LN10 clone, Leica Microsystems), and HLA-DR (TAL 1B5 clone, Santa Cruz). The following monoclonal mouse antihuman antibodies were used: CD4 (4B12 clone, DAKO), CD8 (1A5 clone, Leica Microsystems), CD68 (KP1 clone, ZYMED), CD1a (O10 clone, Gene Tex), and Ki-67 (MIB-1 clone, DAKO). The following rabbit monoclonal antibodies were used: E-cadherin (EP700Y clone, Abcam) and ZO-1 (Polyclonal, Invitrogen). Finally, for CCR5 a monoclonal antihuman antibody, 45523 clone, from R&D Systems was used.
All cell types were counted in the epithelial barrier, except CD4 and CCR5, which were counted in the lamina propria. For epithelial E-Cadherin and ZO-1, the immunolabeling of the protein was analyzed semiquantitatively, using Image J software (NIH, Bethesda MD). In brief, five to six areas were randomly selected using a Nikon E800 microscope from each section and these images were captured using a CCD camera (Spot Camera, Diagnostic Instruments, MI). The integrated optical density (IOD) in each area was calculated for the positive staining color. The IOD of the negative control (no primary antibody) was subtracted from the IOD values for each tissue and the mean value was calculated for the areas of each tissue sample.
Epithelial thickness was previously reported, 5 but the samples were reanalyzed in this study in association with intercellular protein expression. Briefly, epithelial thickness was measured using cross-line reticules (Klarmann Rulings Inc., Litchfield, NH), fitted to the objective of a Nikon Eclipse 600 microscope. The epithelium was measured from the basal lamina to the apical surface in micrometers in 10 random areas per tissue sample and the mean was reported. The mean number of cell layers in the same 10 selected areas of the tissue sample was also reported.
Statistical analysis
Descriptive statistics were obtained by calculating the median, interquartile range, means, and standard deviations for each variable. Since each woman served as her own control, paired tests were used. Paired comparisons were done across the three phases: (1) luteal phase versus follicular phase, (2) luteal phase versus post-DMPA administration, and (3) follicular phase versus post-DMPA administration. For the immune cell and epithelial proteins, the data were not normally distributed and therefore statistical analyses were performed using Wilcoxon signed rank tests. For epithelial thickness and number of epithelial cell layers, the data were normally distributed and therefore a paired t test was used. The test results were interpreted with p-values of 0.05 as statistically significant. Distribution of the data was evaluated by examination of normal quantile plots and with the Shapiro Wilk test. Statistical analyses were performed by use of the SAS statistical analysis software package (SAS for Windows, Version 9.3; SAS Institute, Cary, NC).
Results
Of the 20 women enrolled in the study, results were originally reported on 16 women who completed all study visits. 5 Of these 16 women, 15 had archived paraffin tissue sections suitable for analysis in this study. Women were on average 35.9 (±3.1) years-old, weighed 158.5 (±25.0) pounds, and had an average menstrual cycle length of 29.0 (±1.4) days. All 15 women had previously been pregnant with an average gravidity of 2.7 (±1.1).
We found no significant differences between numbers of vaginal immune cells, activation markers, epithelial junction proteins, number of cell layers, or epithelial thickness in tissues from the luteal and follicular phases of the control menstrual cycle (all p>0.05).
Table 1 shows the differences in the median numbers of immune cell populations between the follicular and luteal phases and after treatment with DMPA. There were significant increases in the median numbers of CD45, CD3, CD8, CD68, CCR5+, and HLA-DR+ immune cells after DMPA treatment compared to the follicular and/or luteal phases of the control cycle. Immune cells displayed a random distribution in the epithelial tissue, with no observed clustering or differential localization across any of the phases of the menstrual cycle or after DMPA treatment. Figure 1 (dot plots) shows increased number of immune cells in tissues obtained after DMPA exposure when compared to the follicular phase. It also demonstrates interindividual variability in the response to DMPA, with some subjects showing a greater change in immune cells than others. Compared with luteal phase samples, vaginal tissues exposed to DMPA showed an increased density of CD3+, CD8+, and CCR5+ immune cells. They also showed an increase in HLA-DR+ cells, which is statistically significant when compared with tissues taken during the follicular phase. Figure 2 shows a composite picture of these cells in the vaginal mucosa during follicular, luteal, and DMPA phases.

Individual changes in vaginal immune cell populations comparing follicular phase and after depot medroxyprogesterone acetate (DMPA) treatment. Total cell density (cells/mm2) for each subject was plotted for the selected immune cell markers. In spite of substantial variability in individual responses, the majority of the samples showed increased numbers of immune cells and activation markers. In some cases, the increase was several fold in magnitude. All cells were quantified in the epithelium, except for CCR5+ and CD4+ cells, which were detected only in the lamina propria.
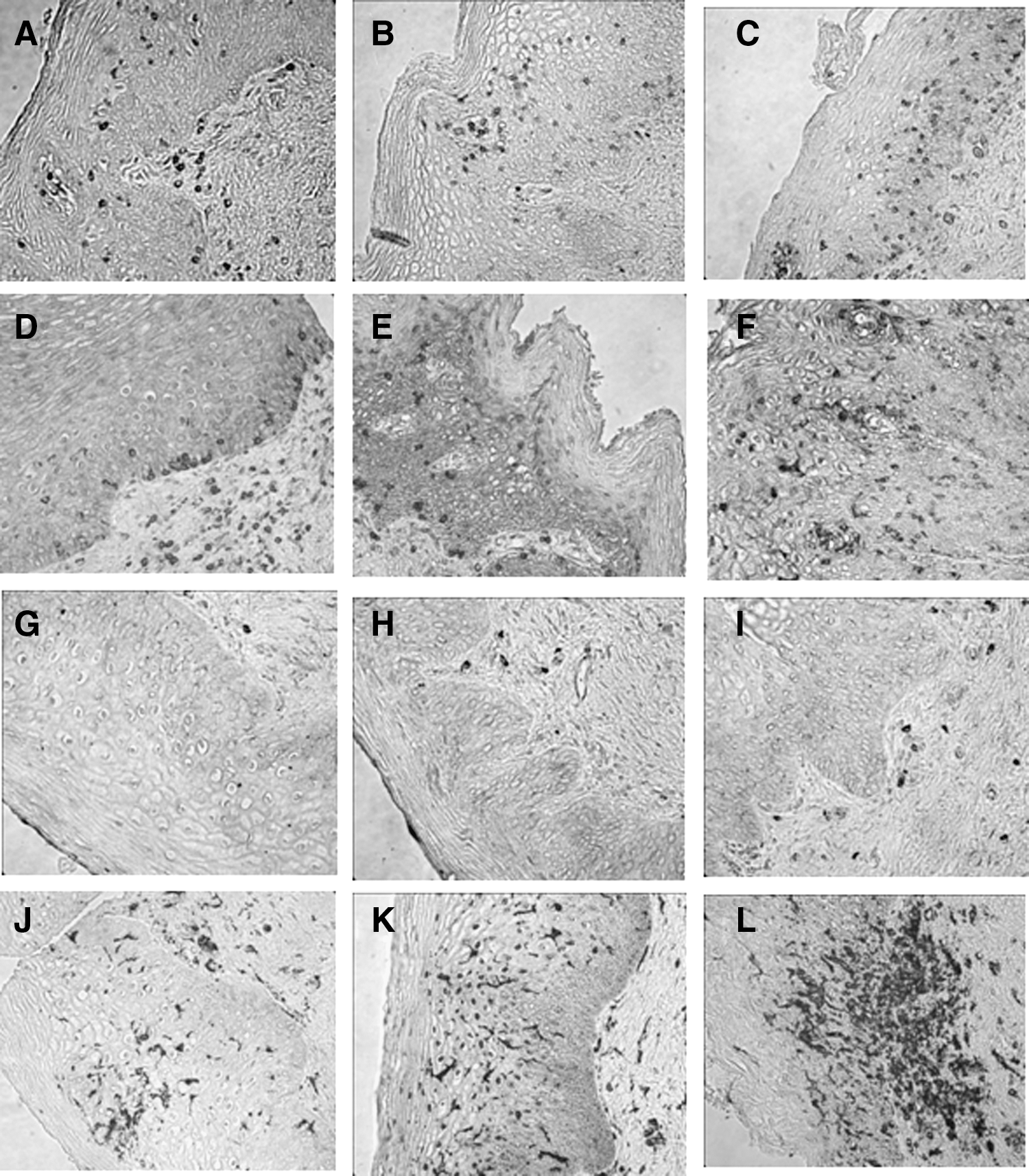
Vaginal immune cells in follicular, luteal, and post-DMPA treatment. Immunohistochemistry staining of paraffin-embedded tissue blocks of vaginal biopsies taken during the follicular phase
Data are not normally distributed and are thus expressed as median (25th percentile, 75th percentile).
CD4 and CCR5-positive cells counted in lamina propria.
Bold numbers indicate statistically significant differences.
DMPA, depot medroxyprogesterone acetate.
Notably, we identified very few CD4+ cells in only three vaginal tissue samples (from a total of 43 evaluable samples for CD4). These cells were exclusively confined to the lamina propria in each tissue sample. CD4+ cell counts were 11.6 cells/mm2 (for one subject in the follicular phase) and 9.3 and 6.3 cells/mm2 (for two subjects after DMPA treatment). Of note, in the three tissue samples where CD4+ cells were seen, there was histological evidence of inflammation, with increased numbers of leukocytes.
In terms of measures of epithelial integrity, there was a trend, although not statistically significant, toward decreased epithelial thickness and number of cell layers in the luteal phase and after DMPA administration when compared to the follicular phase (Table 2). In fact, there was an increase in Ki-67+ epithelial cells, a marker of cell proliferation, when comparing DMPA treatment to either the follicular or luteal phase baselines (Table 2). There were no differences in the density of epithelial tight junction and adherens proteins (E-cadherin and ZO-1) among any of the groups (Table 2 and Fig. 3).

DMPA treatment does not alter epithelial junction protein density. Immunolabeling for the epithelial tight junction protein E-cadherin
Data are not normally distributed and are thus expressed as median (25th percentile, 75th percentile).
Bold numbers indicate statistically significant differences.
Discussion
Exposure to a single dose of DMPA induced changes in the immune cell populations of the human vaginal mucosa, as compared with the follicular and luteal phases of the normal menstrual cycle. These changes consisted of increased numbers of leukocytes (CD45+ cells) and immune cells bearing CD3, CD8, CD68, CCR5, and HLA-DR. Since CD4+ cells were so scarce in the vaginal tissue biopsies, we could not detect statistically significant differences in this cell subset among any of the sampling times. Contrary to the alterations observed in mucosal immune cell populations, we did not see a statistically significant change in epithelial thickness, number of cell layers, or intercellular junctional proteins.
Changes in the mucosal immune system may contribute to the increased risk of acquiring HIV-1 infection reported in certain epidemiological studies of women using DMPA and other progestin-only injectable contraceptives. 8,11 –14 An expansion in CCR5 expressing cells is one of the key changes in the vaginal mucosa that could potentially enhance susceptibility to HIV-1 infection. We found a significant increase in cells bearing this marker in vaginal tissues after DMPA treatment. In addition to being a receptor for the chemokine CCL5 (RANTES), CCR5 is the main coreceptor for mucosal HIV-1 infection in humans, and is thought to be the predominant target for viral entry in the mucosal tissue of the lower female reproductive tract. 31 –33 Human cervical mucosal CD4+ T cells express CCR5, often in conjunction with other secondary receptors for HIV-1 entry. 34,35 In addition, within the lower reproductive tract of both humans and macaques, CCR5 is expressed on mucosal dendritic cells and macrophages, also primary HIV cell targets. While most groups have found that CCR5 is not expressed on intraepithelial CD1a+ Langerhans cells, there is some evidence that the receptor may be present in these cells also. 36 –38 Our findings are in accordance with results from an in vitro model showing that progesterone treatment of peripheral blood mononuclear cells (PBMCs) caused a 5- to 10-fold up-regulation of CCR5 in CD14+ monocytes/macrophages. 39 Furthermore, women in various progesterone-dominant states have been found to have increased expression of cervical and vaginal lymphocytes expressing CCR5. 39 –41 Interestingly, they have also been shown to have increased susceptibility to acquire HIV-1. 42 –45
CCR5 is known to be expressed by activated lymphocytes. 46 Another marker of lymphocyte activation is the histocompatibility antigen HLA-DR. HLA-DR+ T cells are present in the early phases of HIV-1 infection 47 –49 and are thought to account for the majority of the cell population responsible for dissemination of HIV-1 from the mucosal portal to draining lymph nodes and distant sites. 50 Animal models show that HLA-DR+-activated T cells and macrophages are productively infected during the early stage of SIV/HIV infection and constitute one of the main targets for the virus. 51,52
In our study, DMPA increased CD3+ T cells and HLA-DR+ cells. Our findings are consistent with a large longitudinal study that found that white blood cells (WBCs) and polymorphonuclear monocytes (PMNs) were increased in the cervicovaginal fluid lavage (CVL) of women using hormonal contraception. 53 CD3+ cells are widely reported to be the predominant lymphocyte population of the vagina. 54 –57 Although not as numerous in the cervix and vagina as in the upper reproductive tract, vaginal CD3+ T cell populations are not known to be affected by hormonal fluctuations of the menstrual cycle. 54,56 The two main subsets of CD3+ T cells are CD4+ and CD8+ cells 56,57 ; however, CD8+ T cells can outnumber CD4+ T cells in the vaginal epithelium by as much as 8:1. 58,59 CD4+ T cells are a key target for cervicovaginal mucosal HIV-1 infection. 32 Other CD4-bearing cells in the lower female genital tract are dendritic cells (DCs) and macrophages. 37 In vivo and ex vivo data indicate that intraepithelial and submucosal DCs and CD4+ T lymphocytes and macrophages are the first cells targeted by HIV-1. 32,50,60 –63 We detected few vaginal tissue biopsies containing CD4+ cells, and the observed cells were confined exclusively to the lamina propria. Of note, in the three tissue samples in which CD4+ cells were detected, subclinical inflammation was noted. This is in agreement with previous reports describing limited numbers and distribution of CD4+ cells in the vaginal epithelium, especially in the absence of infections or other inflammatory conditions. 28,34,36,59,64 In this study, the presence of STIs or other symptomatic inflammatory vaginal infections such as bacterial vaginosis or trichomoniasis was exclusionary. We have found similar low numbers and confined localization of CD4+ cells to the lamina propria in the mucosa of fresh, noninflamed vaginal tissue obtained from patients undergoing anterior and posterior surgical repairs (data not shown). Furthermore, parallel positive controls using lymph node tissue displayed robust labeling of CD4+ cells, indicating our findings were not due to technical issues in detection (data not shown).
The presence of CD4+ cells in a small percentage of biopsies does not rule out their importance in cervicovaginal HIV-1 acquisition, given the low incidence of HIV transmission, the ability of HIV to penetrate intact epithelium, and the increase in CD4+ cell numbers at mucosal sites of inflammation. 65,66 Furthermore, the average increased susceptibility to HIV-1 reported in observational studies of DMPA users has an approximate mean adjusted HR of 1.50 (1.07–2.09). 6,14,67,68 Therefore, an increase in the number of HIV cell targets, even if present only in a small percentage of the users, may justify the relatively small increased risk for acquiring the infection seen in the population of DMPA users. In our study, although not consistently across all markers, certain subjects showed marked increases of T cells, macrophages, or activated immune cells (HLA-DR+ and CCR5+) (Fig. 2).
We also found significant increases in CD8+ cells in vaginal tissues after DMPA treatment, in agreement with a previous study of long-term DMPA users. 24 CD8+ T cells can exert cytolytic or suppressive functions. Cytolytic activity has been demonstrated in the cervix and vagina throughout the menstrual cycle. 69 However, vaginal CD8+ T cells largely lack TIA1, a marker indicative of cytolytic function in lymphocytes. 26 In addition to being more prevalent than CD4+, CD8+ T cells in the cervicovaginal tissue have a higher expression of activation markers and an increased effector memory phenotype. 70 A sequestered population of CD8+ tissue-resident memory T (TRM) cells appears to be common at skin and mucosal surfaces, including that of the vaginal, and can be recalled by antigen-specific and antigen-nonspecific stimuli. 71,72 Although the presence of CD8+ T cells in the vaginal epithelium appears to be protective against various pathogens, 73 CD8+ T cells coexpressing the transcription factor Foxp3 can also exert suppressive functions through a regulatory T cell phenotype (Treg). 74 Progesterone and its derivative medroxyprogesterone have been associated with immunosuppression of the reproductive tract, inhibiting antibody production, cell-mediated immune responses, and innate immunity. 7 Innate antimicrobial levels decrease during the luteal phase of the menstrual cycle, and exogenous progesterone inhibits human beta defensin-2 production in vaginal epithelial cells. 75,76 MPA also decreases TLR9-induced interferon (IFN)-α production by human and mouse dendritic cells, thereby attenuating potential antiviral responses. 77 It is possible that DMPA suppresses mucosal antiviral mechanisms via CD8+ T cells. Treatment with DMPA stimulated the circulating CD8+ Treg response in macaques. 78 Since we only measured the density of CD8+ cells in this study, and could not perform functional assays or determine cofactor expression, we are unable to clarify the immune function of these cells in the vaginal epithelium.
DMPA has been associated with thinning of the vaginal epithelium. 22 We observed a trend toward decreased epithelial thickness in the luteal phase and DMPA biopsies as compared to those from the follicular phase. This trend, however, was not statistically significant as analyzed by ANOVA with paired t tests for comparison of each of the three sampling intervals. The trend in the data suggests that, on average, women lose cell layers and thus epithelial thickness, when comparing the follicular phase with the luteal phase or DMPA use. However, individual variability is present. For example, 7 of 15 women increased the mean number of vaginal epithelial cell layers, when comparing the follicular phase to after DMPA treatment or from the follicular to the luteal phase. In the previous analysis of these tissues, epithelial thickness and number of cell layers were measured by two different methods and it was noted that there was up to a 34% interobserver variability in measurements. 5 The issue of changes in epithelial thickness after DMPA administration has been investigated by several groups, and it is likely that the inconsistent results reported reflect differences in the method of measurement, statistical analyses, observer bias, timing of tissue sampling, pharmacokinetics, and interindividual variability in the biological response to DMPA. In addition, DMPA administration in humans does not appear to lead to the consistent and marked epithelial atrophy observed in monkey models, although a better dose–response study should be carried out in monkeys to determine a dose of DMPA that leads to pharmacokinetic equivalence with humans.
To investigate the issue of epithelial integrity further, we measured the density of the intercellular adhesion proteins, E-cadherin and ZO-1, which have both been localized in the human vagina and cervix, 79 hypothesizing that a decrease in these proteins could lead to a more permeable epithelium. It has been previously shown that vaginal estrogen alters the concentration of vaginal epithelial tight junctions, 80 but no data exist on the effect of DMPA on epithelial tight junction proteins. We found that a single administration of DMPA did not significantly alter the density of E-cadherin or ZO-1 in the vaginal epithelium. It is known that intercellular adhesion proteins are less concentrated in the apical portions of the vaginal epithelium, as these epithelial cells lose cell-to-cell adhesions and are eventually sloughed into the lumen. 79 We also found that epithelial cell proliferation, as determined by the Ki-67 marker, was significantly increased after DMPA administration, as compared to the luteal and follicular phases. However, the increased proliferation was not reflected in any morphological changes in the number of cell layers or epithelial cell thickness. Women in this study were biopsied at 12 weeks post-DMPA administration, which is the standard length of DMPA treatment. Our findings of no statistical differences in epithelial thickness and number of cell layers, with a statistically significant increase in the Ki-67 marker, could also suggest that at the 12-week time point the vaginal tissue may be in the initial process of reverting back to its normal cyclic state, thereby increasing proliferation, thickness, and cellular junction integrity. It would be ideal to examine epithelial integrity at shorter intervals post-DMPA administration or after prolonged DMPA use. Proliferation of vaginal epithelium after long-term DMPA use has been previously reported by one group. 81 Taken as a whole, our data do not suggest that DMPA causes significant epithelial thinning or compromises epithelial integrity at the 12 week sampling interval. It is unknown if a relatively small reduction in the thickness of the epithelial barrier may facilitate access of virions to mucosal-activated HIV-1 target cells
The data presented in this report represent a pilot study conducted on archived tissue. Since HIV-1 infection was not an endpoint, our data cannot provide a conclusive explanation as to how DMPA use may increase a woman's susceptibility to HIV-1 acquisition. These data, however, do demonstrate a significant effect of exogenous MPA on the number of immune cell populations within the vagina. Importantly, they show changes in immune cell populations known to be involved in the early phases of HIV-1 infection, following exposure to a single injection of DMPA. Lacking functional endpoints and dual marker labeling, we cannot determine if these changes are associated with a mucosal proinflammatory or immunosuppressive state. However, either through increased numbers of activated target cells or reduced antiviral responses, DMPA may be associated with increased cervicovaginal mucosal susceptibility to HIV-1 infection.
These findings are even more critical in the context of recent HIV-1 prevention trials. The majority of these trials require enrolled participants to be on effective contraception; for example, in the CAPRISA 004 trial, 80% of participants used injectable progestin-only contraception, primarily DMPA. 82 Of note, it is not clear if our findings are applicable to other exogenous progestins, widely used as contraceptives. Because millions of women need dual protection from unintended pregnancy and HIV-1, it is essential to characterize the impact of the exogenous hormones on mucosal susceptibility to HIV-1 and the effect of exogenous hormones on microbicide efficacy. A prospective study of approximately 70 women who select combined oral contraceptives or DMPA is planned to further elucidate the effect of exogenous hormones on mucosal susceptibility to HIV-1.
Footnotes
Acknowledgments
This work was supported by intramural CONRAD funds from the U.S. Agency for International Development (Grant GPO-A-00-08-00005-00). The views expressed by the authors do not necessarily reflect those of the funding agency or CONRAD.
Neelima Chandra, PhD, performed the immunohistochemistry (IHC) analyses of all samples. Andrea Ries Thurman, MD, analyzed the IHC data and wrote the manuscript. Sharon Anderson, PhD, analyzed the IHC data and edited the manuscript. Tina Duong Cunningham, PhD, performed the primary statistical analyses of the data and edited the manuscript. Nazita Yousefieh, PhD, performed quality control analyses of the IHC experiments. Christine Mauck, MD, MPH, designed the clinical study and edited the manuscript. Gustavo F. Doncel, MD, PhD, designed the clinical study, analyzed the IHC data, and edited the manuscript.
Author Disclosure Statement
No competing financial interests exist.