
Abstract
A new generation of extremely broad and potent neutralizing antibodies (bNAbs) has been isolated from HIV-infected subjects. This has refocused interest in the sites of vulnerability targeted by these bNAbs and in the potential for designing Envelope (Env) immunogens that display these sites. Standard methods for evaluating HIV-1 vaccine candidates do not enable epitope mapping on the HIV Env spike, the target for NAbs. To meet the need for rapid analysis of Ab specificity, we designed a multiplexed, quantitative mapping assay that can test for serum Ab competition for the binding of an HIV-1 Env gp120 to a panel of bNAbs directed to different sites of vulnerability on the Env that do not compete for one another in the assay. Using serum samples from rabbits immunized with various DNA prime/gp120 protein boost vaccines we were able to detect serum Ab competition for multiple classes of bNAbs in the postimmune samples that were significantly higher than background competition detected in samples obtained prior to vaccination. Importantly, application of this novel assay to our ongoing HIV-1 Env viral vector studies in mice has allowed us to distinguish qualitative differences in the Ab elicited by various regimens that ELISA cannot. Furthermore, pooled immunoglobulin from HIV-infected donors (HIVIg) competes for binding to the bNAb panel whereas a control pool from HIV-negative donors does not, highlighting the utility of this assay for human studies. This novel assay will add value in rational immunogen design and in the detailed, qualitative evaluation of binding and, potentially, neutralizing Abs elicited by natural infections and HIV-1 vaccine candidates.
Introduction
W
Materials and Methods
BG505 HIV-1 Env gp120
The BG505 HIV-1 Env was identified as part of a phylogenetic screen for Envs related to those isolated from study subjects exhibiting potent neutralization activity, as detailed by Hoffenberg et al. 8 The BG505 gp120 sequence was cloned with a C-terminal AviTag peptide sequence into the pCI-neo expression vector using the GeneArt Gene Synthesis service at Life Technologies (Grand Island, NY). BG505 proteins were produced by transient expression from transfected HEK 293T/17 (293T) cells (ATTC # CRL-11268) as described by Hoffenberg et al. Proteins were biotinylated using the Biotin-protein ligase BirA enzyme system according to the manufacturer's protocol (Avidity, LLC, Aurora, CO). This allowed a single biotin molecule to be enzymatically linked to the C-terminus of the protein to engender greater sensitivity and to avoid random biotinylation that could interfere with Ab-epitope interactions. 9
BG505 capture ELISA using MSD plates
Fifty nanograms of Ab was spot-coated in 5 μl of phosphate-buffered saline (PBS) by reverse-pipetting to the center of each well of an MSD High-bind plate (Meso Scale Discovery, Gaithersburg, MD). Plates were left at room temperature (RT) in a laminar flow hood to dry overnight before being washed three times with PBS–0.02% Tween (PBST) in an automated plate washer. Plates were subsequently blocked with 150 μl of 3% bovine serum albumin (BSA) per well with shaking for 1 h at RT. Then 100 ng BG505 was added to each well in 50 μl of 1% BSA and incubated with shaking for 1 h at RT. Plates were washed as above, followed by incubation with 50 μl of 0.5 μg/ml MSD Streptavidin-SULFO-Tag [ruthenium tris(bipyridine), (Ru(bpy)3)] for 1 h at RT with shaking. Plates were washed as above, followed by addition of 150 μl MSD tripropylamine read buffer. Plates were read in an MSD Sector Imager 2400 (Meso Scale Discovery, Gaithersburg, MD).
Cross-competition and serum competition experiments (single-plex)
Serial dilutions of competing Ab (200 μg/ml–0.0003 μg/ml) or serum (1:4–1:2,048) in 30 μl 1% BSA were preincubated with 120 ng of BG505 in 30 μl of 1% BSA per well for 1 h at 20°C with shaking before addition of 50 μl of Ab/BG505 mixture to coated and blocked MSD High Bind plates. Percent competition was calculated using the formula {[1 – (test well ECL/BG505 only control well average ECL)]×100}. Percent competition values at each Ab concentration/serum titer were fitted with four-parameter curves from which ID50 or ID80 concentrations were interpolated using GraphPad Prism 5.03.
It was determined that ID80 titer was a superior readout for serum competition compared to ID50 titers, as preimmune rabbit sera did not achieve 80% competition even at low dilutions, and median increases in preimmune to postimmune competition were 43% higher than when ID50 was used as a readout (not shown).
Multiplex CBA
Initially, using an MSD 7-spot Plate Optimization Package (MSD, Gaithersburg, MD), 18 prototype high-bind 7-spot MSD plates were machine coated by proprietary means at MSD headquarters with 75, 150, or 300 μg/ml of each Ab (5.6, 11.2, 22.4 ng/spot, respectively) in PBS or proprietary MSD BSA stabilizer coating buffer (i.e., six coating conditions were tested). In addition to a 3% BSA control spot, the Abs selected for the 7-plex panel included 2F5 (second binding control), PG9, b6, 17b, PGV04, and PGT126, i.e., those tested with rabbit sera by single-plex CBA. Cross-competition and serum experiments were performed exactly as described for the single-plex CBA (above), using 2.4 μg/ml BG505 (2.1×molar excess of total Ab coated per well).
Reverse-format CBA
One hundred nanograms of PG9 was spot-coated to MSD High Bind plate wells as described above. After blocking, plates were incubated at RT with 100 ng unbiotinylated BG505 in 50 μl of 1% BSA for 1 h with shaking. Plates were washed, followed by incubation with serum titrated 2-fold (1:8–1:16,384) in 50 μl 1% BSA. Plates were then washed and incubated with 100 ng biotinylated PGV04 in 50 μl 1% BSA. After washing, plates were then treated with Strep-SULFO-Tag and read as described above.
Multiplex CBA with HIVIg and control purified, pooled human IgG
HIVIg was obtained from the NIH AIDS Reference and Reagent Program (catalog number 3957) in a 50 mg/ml solution. Pooled purified human IgG (Hizentra, CSL Behring, Switzerland) from HIV-negative donors was used as a control. Reagents were titrated 2-fold (1,000–0.24 μg/ml) in 1% BSA prior to incubation with 2.4 μg/ml BG505. The CBA was then performed as described above.
Naive sera tests
Pooled HIV-naive serum from mice, ferrets, Chinese rhesus and Indian rhesus macaques, and humans (from human male AB plasma; Sigma-Aldrich, St. Louis, MO) were titrated 2-fold (1:4–1:2,048) in 1% BSA and the CBA was performed as described above.
ELISA and Multiplex CBA with sera from vaccinated mice
ELISA was performed using polystyrene, flat-bottom, high-binding ½ area plates (Corning, Corning, NY) coated with 50 ng of vaccine-matched HIV-1 Env gp120 antigen and incubated at 4°C overnight. Plates were then washed and blocked as described above. Plates were washed and serial dilutions of mouse serum (1:100–1:5,904,900) were added. Plates were incubated at 37°C for 1 h. Plates were washed and incubated with 50 μl horseradish peroxidase-conjugated antimouse IgG (Jackson ImmunoResearch, PA) at a dilution of 1:5,000. Plates were washed and developed with 50 μl of 3,3′,5,5′-tetramethylbenzidine (TMB) added (Thermo Scientific, Rockford, IL), followed by a 5 min incubation at room temperature. Sulfuric acid (2 N) was added, and plates were read in a VERSAmax plate reader (Molecular Devices, Sunnyvale, CA) at 450 nm according to the manufacturer's protocol. OD values were fit to four-parameter curves and an OD cutoff of 0.2 was used for endpoint interpolation using GraphPad Prism 5.03. p-values were determined by a one-tailed Mann–Whitney test. The multiplex CBA using mouse sera from vaccinated mice was performed as described above for rabbit serum.
Animals
New Zealand rabbits were maintained at Covance (Denver, PA) under specific pathogen-free conditions. All experimental protocols were approved by the Covance Institutional Animal Care and Use Committee. C57BL/6 mice were maintained at SUNY Downstate Medical Center under specific pathogen-free conditions. All experimental protocols were approved by the SUNY Downstate Institutional Animal Care and Use Committee.
Immunizations
Rabbits were anesthetized and then immunized by intramuscular electroporation with 250 μg HIV-1 Env DNA using the TriGrid Delivery System (Ichor Medical Systems, Inc., San Diego, CA). Rabbits were primed with DNA twice at weeks 0 and 4, before an intramuscular boost at week 12 with HIV-1 Env protein adjuvanted with Iscomatrix formulated according to the manufacturer's protocol (CSL, Kankakee, IL). Mice were anesthetized and then immunized by intramuscular electroporation with HIV-1 Env DNA with plasmid IL-12 using the TriGrid Delivery System (Ichor Medical Systems, Inc., San Diego, CA). Mice were primed with DNA twice at weeks 0 and 3, before boosting at week 6 with a replicating viral vector encoding HIV-1 Env or with a third DNA-IL-12 injection.
Results
The CBA in single-plex format
Initially, the CBA was set up in a 96-well single-plex format in order to establish Env binding and optimal serum concentration. MSD high-bind plate wells were coated with a single anti-HIV-1 Ab (see Table 1 for list of Abs used and their specificities), and an excess of biotinylated BG505 was added, followed by Streptavidin-SULFO-Tag. Plates were read in the Sector Imager 2400. We found that BG505 bound to gp120-specific Abs with strong affinity while exhibiting virtually no binding to 2F5, a gp41(MPER)-directed antibody, nor to a 3% BSA control (Fig. 1A). We next aimed to evaluate the specificity of the CBA by assessing the ability of Abs in solution to compete for binding of BG505 to homologous capture Abs, as well as to identify any nonspecific competition by heterologous Abs. Therefore, we performed a cross-competition experiment by preincubating BG505 Env with titrated competing Abs in solution before addition to the plate. As expected, most Abs exhibited homologous competition, with IC50 values of ≤15 μg/ml, whereas heterologous competition between Abs directed at different regions of vulnerability required concentrations 6- to 83-fold greater, with most not achieving 50% competition even at the highest concentration tested (Table 2). Notably, cross-competition by the nonneutralizing CD4bs Ab b6 for BG505 binding to the neutralizing CD4bs Ab PGV04 required 22-fold more Ab than homologous b6 competition. Importantly, during early assay development it was determined that while preincubation of Ab with BG505 at 37°C engendered increased heterologous competition between Abs, 20°C was optimal for specific Ab/BG505 interactions, particularly with regard to b6 competition for coated Abs (Supplementary Fig. S1; Supplementary Data are available online at

BG505 gp120 Env binds a diverse panel of HIV-specific antibodies.
Denotes antibodies chosen for the 7-plex competition binding assay. Potency/breadth as previously described. 3
Based on this cross-competition data, five Abs were selected to further characterize serum antibody responses, including three bNAbs that bind to different sites on gp120: PG9 (anti-V1/V2 quaternary epitope), PGV04 (CD4bs), and PGT126 (anti-glycan/V3 loop), as well as two nonneutralizing Abs, b6 (CD4bs), and 17b (CD4-induced). Additionally, anti-C-terminal AviTag Ab was used as a control for nonspecific serum reactivity. This Ab panel was used to test the serum of four rabbits immunized with experimental DNA-prime/Env-gp120 protein-boost regimens. Titrated preimmune and postimmune serum was incubated with BG505 prior to the addition to the Ab-coated plate. It was found that postimmunization, serum had varying but largely detectable CBA titers to all five anti-HIV Abs in the panel relative to preimmune levels (median fold increase, 7.6), whereas reactivity to the anti-AviTag Ab did not change (Fig. 2).
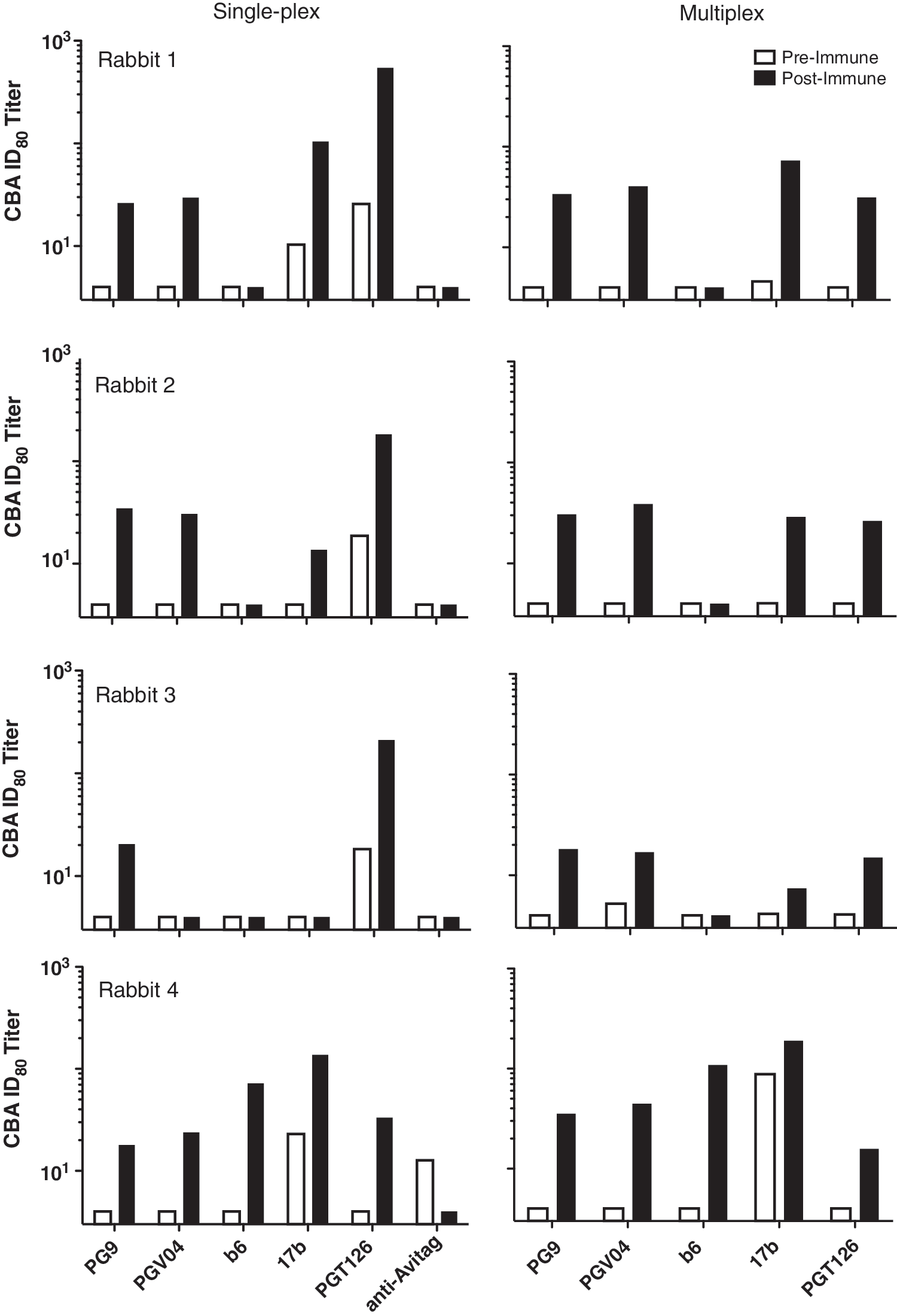
Competition binding assay using serum from four immunized rabbits. Serial dilutions (1:4–1:2,048) of preimmune or postimmune serum obtained from rabbits vaccinated with an HIV-1 Env DNA prime, gp120 protein boost regimen were preincubated with 120 ng (single-plex format) or 72 ng (multiplex format) of BG505 per well as described in Materials and Methods and added to blocked MSD high-bind single-plex or multiplex plates. Percent competition values were fit to four-parameter curves from which ID80 values were interpolated using GraphPad Prism 5.03.
We next aimed to determine the effect of having the CBA Env antigen in the solution phase, which we hypothesized would facilitate competition in a physiologically relevant format. As such, we reversed the assay configuration and coated BG505 directly on the MSD plate. In this configuration, we were unable to detect competition using postimmune rabbit serum (Supplementary Fig. S2).
Optimization of the multiplex CBA
The goals of this optimization were to assess ECL levels of BG505 binding to the coated Abs at each coating condition, to determine which coating condition gives the best reproducibility within and between plates, and to compare competition titers obtained for each coating condition. Predictably, ECL tended to increase with increasing coat concentration, and plates coated with Abs in the proprietary MSD BSA stabilizer tended to exhibit reduced ECL compared to plates coated with Abs in PBS (Fig. 1B and Supplementary Fig. S3A). However, there were certain exceptions to these trends; for example, the presence of BSA stabilizer exhibited little or no effect at the 75 μg/ml coating concentration for PG9 and b6, while for PGT126 and somewhat for PGV04, the presence of the stabilizer actually increased ECL, suggesting a “hook effect” phenomenon in the absence of the buffering effects of the BSA stabilizer. Furthermore, it was found that intraplate and interplate reproducibility was similar among all coating conditions, with the exception of the 75 μg/ml plate without BSA stabilizer, which exhibited the greatest ID80 variation, particularly within a plate (∼1.2- to 1.5-fold difference between replicates; Supplementary Fig. S3B). It was also observed that postimmune ID80 titers decreased with increasing Ab coat concentration for PGV04 and PGT126 competition (Supplementary Fig. S3C). Given these findings, the 75 μg/ml+BSA stabilizer condition was ultimately selected for the multiplex CBA.
The CBA in multiplex format
Cross-competition experiments were repeated using the multiplexed CBA plates. Consistent with the single-plex assay, homologous Abs were found to compete with themselves in the multiplex format at relatively low concentrations (IC50 ≤6.4 μg/ml), and 80% competition was achieved for four of five Abs, requiring ≤8 μg/ml. Interestingly, homologous competition in the multiplex assay required 3- to 22-fold less Ab compared with the single-plex assay (Table 3). This is likely the result of having a lower amount of antibody concentrated in each spot, thereby requiring less exogenous antibody for competition. Rabbit sera tested in the single-plex CBA were retested using the 7-plex format. We found that serum ID80 titers determined using the multiplex CBA were similar to those determined using the single-plex assay for PG9 and b6; however, rabbit 4 exhibited 1.5- to 2-fold higher postimmune titers using the multiplex assay. PGV04 competition titers were found to be roughly 2-fold higher on average using the multiplex assay, though this ranged from a 1.3- to 4.2-fold difference, depending on the serum tested. Generally, the effect of the multiplex format on 17b competition was quite variable, with rabbit 1 exhibiting 1.4- to 2.3-fold reductions in ID80 titer compared to the single-plex assay, while rabbits 2–4 exhibited 1.4- to 3.8-fold increases. In contrast, PGT126 competition was notably reduced for virtually all sera tested, with an average 7.1-fold ID80 titer decrease (range, 2.1- to 17-fold decrease), suggesting the multiplex format and coating conditions exhibited a unique effect on PGT126, drastically reducing the competition observed with the single-plex format (Fig. 2). Overall, it was evident from the variability observed between the two assay formats that data cannot be compared if both formats are used, and that only one format should be employed to analyze a single vaccine study or sample set.
To determine the suitability of the CBA assay for use with serum from other species, we tested pooled, HIV-naive sera from mice, ferrets, Chinese and Indian rhesus macaques, and humans in the multiplex format. As was observed with naive rabbit serum, these sera did not achieve 80% competition, even at the lowest serum dilution tested (1:8), with the exception of human serum, which exhibited very low nonspecific ID80 titers (Table 4). As a future goal of this novel assay is to employ it to map Ab specificities in human samples, we aimed to determine whether the CBA could detect Ab responses in HIV-infected humans. Pooled, purified immunoglobulin from HIV-infected donors (HIVIg) was therefore assayed in the multiplex format. HIVIg exhibited IC50 values of 67–127 μg/ml for binding to PG9, PGV04, PGT126, and b6, but did not bind to 17b. A control pool of HIV-naive purified human serum immunoglobulin did not achieve 50% competition in the CBA (Table 5).
ID80 titers are shown. Ch.RM, Chinese rhesus macaques; In.RM, Indian rhesus macaques.
Values shown in μg/ml.
Since pooled, naive mouse serum exhibited low baseline competition in the CBA, we assayed sera from groups of mice immunized with either PBS (mock control), or one of four HIV-1 Env DNA-prime/replicating viral vector-boost regimens obtained from two separate experiments. In both experiments, ELISA endpoint titers of total anti-HIV-1-Env IgG showed no quantitative difference between the two Env regimens tested (Fig. 3A and C); therefore, we applied the CBA in order to determine if the Abs elicited by these vector regimens might be qualitatively unique. It was found for each experiment that the two regimens tested elicited qualitatively different Abs. In the first experiment, it was determined that CD4bs-directed Abs were significantly increased in one regimen compared to the other (p=0.04; Fig. 3B

Competition binding assay using serum from vaccinated mice. Mice were vaccinated with either PBS (mock vaccine) or one of four HIV-1 Env DNA-prime/replicating viral vector-boost regimens obtained from two separate experiments.
Discussion
Various methods for mapping the specificities of serum Abs reactive to HIV-1 Env have been developed, including peptide interference, reactivity to scaffolded Env regions, pseudoviral mutagenesis, and competition by mAbs with known specificities in an ELISA format. 5,10 –17 Though these methods have proven valuable for specific applications, they have intrinsic limitations that diminish their utility. For example, each assay can typically test only one specificity, thus testing for multiple epitopes requires high sample volume. Furthermore, peptide-based mapping assays detect antibodies that recognize linear epitopes but ignore responses to discontinuous or conformationally sensitive epitopes.
The identification of a “new generation” of bNAbs that can collectively neutralize >90% of viruses tested has engendered new interest in the design of an HIV vaccine candidate that elicits similar Abs. 3,6 Rapid, quantitative evaluation of such vaccines for elicitation of antibody responses to known sites of vulnerability on the virus will clearly benefit rational immunogen design. To meet this need, we developed a novel, high-throughput, multiplexed CBA that can rapidly assess multiple Ab specificities in a single well using only 15 μl of serum.
The present study suggests that the CBA will be suitable tool for analysis of Ab quality elicited in preclinical studies using various animal models, such as mice, rabbits, and rhesus macaques, as well as analysis of human samples. The CBA may be particularly useful when neutralization assays are not feasible, such as with mouse serum, which is obtained in low volumes and tends to exhibit high levels of nonspecific neutralization. Without neutralization data, assessment of Ab is typically based on ELISA titers alone, which, as was the case in the highlighted mouse experiments described herein, do not necessarily provide an accurate picture of the quality (i.e., specificity) of the Abs elicited. In the mouse experiments presented, critical information regarding immunogen selection and the effects of each vaccine regimen would have been lost without the benefit of the CBA. Furthermore, even when neutralization assays can be performed effectively, providing a more traditional assessment of Ab quality, additional, rapid analysis of Ab specificity by the CBA will similarly provide critical qualification of ELISA data, acting as a bridge between binding and neutralization studies, to the benefit of rational immunogen design.
The BG505 Env was selected for its ability to bind to a wide array of anti-HIV-1 Abs, as described extensively by Hoffenberg et al. Though outside the scope of this study, any Env antigen and competition panel could be used in the CBA, as long as the antigen is capable of binding the Abs chosen. It is likely that extensive Ab qualification could be achieved by performing the CBA using various clade-matched and clade-mismatched antigens. Furthermore, the antigen need not be gp120, as biotinylated gp140 monomers, Foldon or other gp140 trimers, or, potentially, trimer-expressing viruses could also potentially be employed in the CBA. Use of trimers would be particularly useful for studying serum competition for trimer-specific Abs, such as PGT145 and PG16, and would certainly provide a more complete and relevant picture of elicited Ab, particularly with regard to those facilitating virus neutralization. 3,4 Use of HIV-1 Env trimers in the CBA is under investigation and will be the subject of further studies. The CBA Ab panel described in the present study represents three sites of HIV Env vulnerability that do not cross-react in the CBA, as detailed in the Ab cross-competition studies performed initially. Depending on the antigen used, additional Abs could be added to the panel while maintaining competition specificity. Though not yet explored, inclusion of the nonneutralizing b6 as well as the neutralizing PGV04 in the Ab panel, which both target the CD4bs yet do not cross-compete in the CBA, may yield useful data when compared to neutralization data in future studies.
Optimization of assay conditions was essential for reducing Ab cross-competition in the CBA, particularly of b6 for coated PGV04, and to a lesser degree, PGT126 and PG9. To minimize cross-competition, it was necessary to reduce the preincubation temperature from 37°C to 20°C and to perform all steps in PBS–1% BSA. Previous work by Leaman et al. 18 with b6 and the neutralizing CD4bs Ab b12 similarly showed that higher temperatures may engender b6 promiscuity. Though we expected that “stronger” buffering systems such as milk or FBS would be the most effective at limiting Ab cross-competition, ultimately it was the BSA buffering system recommended by MSD that reduced b6 cross-competition most efficiently. Inclusion of additional Abs in future CBA panels may require similar or additional optimization, particularly if the Abs recognize similar specificities. Furthermore, care must be taken when Abs are included in a competition panel that do not exhibit homologous competition in the CBA, such as was seen in the present study for 17b. This Ab recognizes the chemokine coreceptor binding site on Env, requiring soluble CD4 for efficient binding or neutralization. 19 Like b6, 17b was included in the panel as a nonneutralizing Ab for future investigational purposes. However, cross-competition experiments, when performed in the single-plex format, indicated that 17b in solution could not compete for coated 17b, while in the multiplex assay, ID50 was achieved, but 2.5-fold or more Ab was required compared to the rest of the panel, and this difference between assay formats was likely due to the reduced amount of 17b coated in multiplex format compared to single-plex (5.6 ng vs. 50 ng/spot). As well, nearly 3-fold more HIVIg was required to achieve ID50 for 17b binding compared to the other panel Abs, suggesting this Ab may not be binding BG505 with sufficient specificity in the absence of soluble CD4 and that it may not be suitable for inclusion in future panels.
The CBA is highly innovative as it enables simultaneous, quantitative mapping of serum antibody responses to multiple regions of vulnerability on the HIV-1 virus targeted by bNAbs using gp120 antigen in a native, soluble configuration. Herein we have shown that vaccination of rabbits and mice with various HIV-1 Env immunogens can elicit serum Abs that compete for the binding of gp120 Env to a panel of bNAbs, indicating that these vaccine regimens have produced Abs that target one or more important sites of Env vulnerability. Ongoing studies using this novel assay system are directed at the evaluation of specific vaccine candidates and the correlation of CBA titers with virus neutralization among a large panel of human sera, so that the assay may be further developed for clinical use.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.