
Abstract
Recent studies suggest that HIV-specific antibody-dependent cell-mediated cytotoxicity (ADCC) antibodies contribute to protective immunity against HIV. An important characteristic of future HIV vaccines will, therefore, be the ability to stimulate production of these antibodies in both men and women. Early studies suggest that men may have a better ADCC antibody response against HIV than women. Our objective was to determine whether men and women differ with respect to their ADCC response to HIV-1 gp120. HIV-positive, asymptomatic untreated men and women were matched for race, age, CD4+ T cell number, HIV-1 viral load, and treatment and HIV-1 gp120 ADCC antibody titers were compared. A standard 51Cr-release assay was used to determine HIV-1 gp120 ADCC antibody titers in HIV-1-seropositive individuals from the Multicenter AIDS Cohort Study (MACS; n=32) and the Women's Interagency HIV Study (WIHS; n=32). Both sexes had high ADCC titers against HIV-1 gp120: 34.4% (n=11) and 40.6% (n=13) of men and women, respectively, had titers of 10,000; 62.5% (n=20) and 56.3% (n=18) had titers of 100,000. Groups did not differ in percent specific release (% SR), lytic units (LU), correlations of titer to viral load, or titer to CD4+ T cells in men or women. Both groups also had similar cross-clade ADCC antibody responses (p>0.5 for % SR and LU). Comparable groups of asymptomatic HIV-1-infected men and women had comparable HIV-1 gp120 ADCC antibodies. Both sexes had significant cross-clade reactivity. Differences between men and women may become evident as disease progresses; this should be evaluated at later stages of HIV-1 infection.
Introduction
Results from RV144, a human vaccine trial for HIV in Thailand, indicate that approximately 30% of subjects may have been protected. This is perplexing because the two major immune effector mechanisms thought to be responsible for protection, cytotoxic T lymphocytes (CTL) and neutralizing antibodies, were not present in immunized individuals. 1 Further analysis revealed an inverse correlation between HIV-env-specific antibody-dependent cell-mediated cytotoxicity (ADCC) antibodies and risk of infection. 2,3 This has spurred a renewed interest in ADCC.
In ADCC, Fc receptor-bearing cells, such as natural killer (NK) cells and monocytes, together with virus-specific antibody, bind to HIV envelope on infected cells and kill them. This reduces the number of cells producing virus, lowers viral load, and slows pathogenesis.
It is not surprising that ADCC antibodies would be considered an important component of a vaccine or a therapeutic tool since monoclonal antibodies that have functional ADCC activity are used therapeutically to treat cancer. 4 There have been efforts to enhance ADCC-mediated killing by removing fucose from ADCC antibodies 5 and to characterize the HIV epitopes that are recognized by these antibodies. 6 Strong support for ADCC as an important component of vaccines against retroviruses comes from a study by Alpert et al. in which rhesus macaques were vaccinated with a live attenuated vaccine and challenged with neutralization-resistant simian immunodeficiency virus (SIV). Animals with envelope-specific ADCC antibodies were protected, suggesting that ADCC antibodies were essential for protection mediated by this retrovirus vaccine. 7 Other recent studies supporting a role for ADCC in host defense against HIV include evidence of its ability to prevent HIV transmission through breastfeeding, 8 confirmation that ADCC killing correlates inversely with viral load, 9 and the suggestion that ADCC exerts immune pressure on HIV replication. 10 –12 If ADCC antibodies are an important part of the immune defense against HIV, it is important to determine whether HIV vaccines that are designed to stimulate an ADCC response against HIV will work equally well in men and in women.
The Multicenter AIDS Cohort Study (MACS) is an ongoing, NIH-sponsored, prospective study of HIV infection in men who have sex with men (MSM). It was initiated in 1984. The more than 7,000 participants in this study are homosexual and bisexual men who report participation in activities that put them at high risk of HIV infection; once enrolled, they have a semiannual interview, physical and mental evaluation, and HIV testing and treatment. They also provide clinical samples that have been used to establish a national repository that can be accessed by basic and clinical investigators in order to study different aspects of HIV disease. MACS sites include Los Angeles, Chicago, Baltimore/Washington, and Pittsburg.
The Women's Interagency HIV Study (WIHS) is an ongoing, NIH-sponsored, prospective study of HIV infection in women. It was established in 1993. Women who enroll in the WIHS are interviewed and receive a physical examination including a gynecologic examination and laboratory testing. They also provide clinical samples that contribute to a valuable repository that can be used to study the HIV epidemic. Until recently there were six WIHS sites: New York City/Bronx, Brooklyn, Washington, D.C., Chicago, Northern California, and Southern California, with a Data Coordinating Center in Baltimore. Four new sites have recently been established that represent areas of the South Eastern regions of the United States where HIV infection rates in women are high and increasing.
Our previous work demonstrates that in HIV-1-infected individuals with a wide range of viral loads, ADCC antibody titer correlates with the absolute CD4+ T cell number. 13 Furthermore, we show that ADCC antibodies against HIV-1 are beneficial to the host and that high titers of antibodies correlate not only with higher CD4 T cell numbers, but also with a lower viral load. 14 These were separate studies of ADCC activity in 394 carefully chosen samples from 118 HIV-1-infected men from the Multicenter AIDS Cohort Study (MACS) 14 and 330 women from the Division of AIDS Treatment and Research Initiative (DATRI 009). 15 Even though the purpose of the MACS study was to show a correlation with disease progression and the purpose of the DATRI study was to report ADCC activity in the genital fluids of HIV-infected women, one striking observation from these studies was that men from the MACS had higher % specific release (% SR), higher lytic unit (LU) values, and higher ADCC antibody titers than women from the DATRI 009 study. For example, men had LU values as high as 1,101 and a LU of 500–600 was frequent. The focus of the DATRI study was genital ADCC, but paired serum ADCC activity was evaluated for each woman. While greater than 90% of women had serum ADCC activity, women more commonly had LU values 10-fold lower than men. While each cohort was composed of HIV-1-infected subjects from a single sex, they had different demographic and clinical criteria for enrollment.
The participants in the MACS were primarily homosexual or bisexual white men whereas the participants in the DATRI 009 cohort were predominantly heterosexual African-American women. The MACS samples were longitudinal samples from carefully selected groups of rapid progressors, moderate progressors, and long-term survivors while the DATRI 009 samples were cross-sectional samples from women at various stages of disease progression. The men had not been treated at the time the samples were collected since we utilized archived samples that had been stored prior to the availability of antiretroviral medications. Samples from the DATRI 009 study were actually collected a decade later than the MACS samples and included participant women some of whom had been treated with antiretroviral drugs because of their more advanced HIV disease.
Since there were many possible explanations for the apparent sex difference in ADCC activity in previous studies, we designed a more controlled study to compare ADCC antibodies in men and women. We obtained current matched samples from the MACS and from one of the cohorts that had contributed to the DATRI 009 sample set, the Women's Interagency HIV Study (WIHS), in order to determine whether the apparent differences between serum ADCC activity in men and women were related to sex as opposed to other factors such as race, age, stage of disease, viral load, or antiretroviral treatment. Serum samples in this study were all from HIV-1-infected asymptomatic men and women who were not on retroviral treatment. These samples were matched for age, race, and absolute numbers of CD4+ T cells and assayed for ADCC antibody activity. This study was not designed to correlate ADCC activity with disease progression since the study participants were relatively healthy and the ranges of CD4+ T cells and viral loads were narrow.
In addition to determining if men and women had similar ADCC antibody titers against HIV-gp120, we also wanted to determine whether there were differences in cross-clade reactivity between HIV-1-infected men and women. Following an initial assessment of sex differences in ADCC activity against homologous clade B-derived HIV-1 gp120, which is prevalent in North, Central, and South America and most of Europe, 16,17 we looked for differences in ADCC activity against heterologous clade A/E-derived HIV-1 gp120, which is prevalent in Thailand. 18
Evidence from separate published studies that suggest that sex differences in the ADCC antibody response to HIV, coupled with differences in HIV-1 disease progression in men and women, 19,20 lead to the concern that vaccines designed to stimulate the production of ADCC antibodies might not work equally well in men and women. In this study we directly compared ADCC antibody activity against HIV in groups of HIV-1-infected men and women who were carefully matched with respect to age, race, and CD4 T cell number.
Materials and Methods
Study population
Serum samples were obtained from the MACS (n=32) and from the WIHS (n=32); all subjects were asymptomatic HIV-1-seropositive individuals who reported no prior antiretroviral therapy. The MACS is a prospective continuing study designed to evaluate the impact of HIV-1 infection in men; it has sites in Baltimore, Pittsburg, Chicago, and Los Angeles. The WIHS is a prospective cohort study designed to evaluate the impact of HIV-1 infection in women; it has sites in New York, Washington, D.C., San Francisco, Southern California, and Chicago. Both studies conducted interviews, physical examinations, and specimen sampling at enrollment with similar follow-up visits every 6 months. 19,20 The women in this study were taking a variety of oral contraceptives. The MACS and WIHS seropositive samples selected for this study were matched for CD4+ T cell number, plasma viral load, age, and ethnicity (Table 1). Since individuals selected for this specific study were not seroconverters, but were seroprevalent when they entered these prospective studies, we do not know whether the men and women we studied were infected for comparable lengths of time. All of the 64 individuals studied are most probably HIV-1 clade B since all of the men and 29 of the women were from North America. The remaining three women were from Granada, Honduras, and Jamaica. Clade B is also the predominant clade in these countries. 21 –23 In addition, five female and five male HIV-1-seronegative serum samples were obtained from healthy donors who were not participants of either the MACS or WIHS cohort.
Characteristics of Study Participants
Mean±standard deviation.
Study participants from the Multicenter AIDS Cohort Study (MACS) and Women's Interagency HIV Study (WIHS) (n=32/32) were systematically matched for age, HIV-1 viral load, CD4 cell count, and ethnicity prior to evaluating sex differences in antibody-dependent cell-mediated cytotoxicity (ADCC) against HIV-1 gp120.
Target cells
A natural killer (NK) cell-resistant CD4+ cell line, CEM.NKR (NIH AIDS Research and Reference Reagent Program, Germantown, MD), was used as target cells in a standard 51chromium (51Cr)-release assay. When cells are labeled in log phase, this cell line has less than 5% SR activity in the absence of antibody (background NK cell activity) and more than 90% of these cells bind and retain HIV-1 gp120 for the duration of the assay. 14 In chromium-release assays, 6×106 target cells in RPMI-1640 (Invitrogen Corp., Carlsbad, CA) with 10% fetal bovine serum (FBS) (Atlanta Biologicals, Lawrenceville, GA) were incubated with 100 μCi of Na[51Cr]O7 (Perkin Elmer, Downers Grove, IL) for 1 h at 37°C in a humidified 5% CO2 incubator. Following incubation, 1×106 cells were washed and pulsed with 0.225 μg of recombinant HIV-1 MN gp120 (Immunodiagnostics, Inc., Woburn, MA) or 0.225 μg of recombinant HIV-1 CM gp120 (Protein Sciences Corp., Meriden, CT) at room temperature for 1 h. The cells were washed and 1×104 of target cells bearing rgp120 were added to each microtiter well.
Effector cells
Peripheral blood mononuclear cells (PBMC) were freshly isolated from healthy HIV-1-seronegative donors by Ficoll gradient centrifugation using lymphocyte separation medium (Mediatech Inc., Manassas, VA). The PBMC were rotated with carbonyl iron (Sigma, St. Louis, MO) for 1 h followed by magnetic separation to remove the monocytes and isolate the peripheral blood leukocytes (PBL). The PBL were washed and resuspended at a concentration of 8×106 cells/ml, 4×106 cells/ml, and 2×106 cells/ml in complete media to achieve effector to target (E:T) cell ratios of 40:1, 20:1, and 10:1, respectively.
Antibody source
Sera from HIV-1-seropositive MACS (n=32) and WIHS (n=32) participants were used as the source of antibodies. To accurately compare HIV-1 gp120-specific ADCC titers in males and females, 10-fold serum dilutions ranging from 1:100 to 1:100,000 were evaluated. Since the serum concentrations with maximum activity were determined in the initial analysis, these peak dilutions were assayed to evaluate gp120 cross-clade reactivity. Pooled IgG from HIV-1-seropositive individuals, HIVIg (North America Biologicals, Miami, FL), was used as a positive control in all 51Cr-release assays at a 1:2,000 dilution, which was previously determined to be optimal. HIV-1-seronegative sera from healthy male and female donors (m/f=5/5) were obtained and diluted the same way as the experimental samples for all assays.
Chromium release assay
A total of 1×104 target cells (in 100μl) were added to wells of a 96 V-bottom plate (Corning, NY) and 50 μl of study serum was added to give final dilutions of 1:100, 1:1,000, 1:10,000, and 1:100,000. After a 10-min incubation of target cells and serum, 50 μl of the effector cells was added to each well at the appropriate concentration to give the three E:T ratios. To determine spontaneous and maximum release values, target cells were incubated with either 100 μl of complete media or 100 μl of 0.1% Triton X-100 (Sigma, St. Louis, MO). Plates were then incubated for 3.5 h in a humidified 37°C/5% CO2 atmosphere. Cultures were assayed in quadruplicate. After incubation, 100 μl of the supernatant was harvested from each well and γ-irradiation was measured as counts per minute (cpm) using a 2470 Wizard Gamma Counter (Perkin Elmer, Torky, Finland). cpm were converted to % SR according to the following formula: % SR=[(experimental CPM – spontaneous CPM)/(maximum CPM – spontaneous CPM)]×100. Percent SR from three E:T values was used to determine lytic unit20 (LU20) values. A LU20 is roughly based on the number of effector cells required to lyse 20% of the target cells added to a well. This number was calculated for each dilution using the exponential fit method developed by Pross and Maroun. 24
ADCC titer
We assessed the serum of 10 healthy HIV-1-seronegative donors (m/f=5/5) to establish a baseline ADCC response. HIV-1 gp120-specific ADCC antibody titers were obtained from % SR and LU20 values at each antibody dilution. Background NK activity (activity in the absence of antibody) for the individual assay was subtracted to standardize results prior to comparison of data. ADCC antibody titers were determined by taking the inverse of the highest antibody dilution at which HIV-1 gp120-specific ADCC activity was more than three standard deviations greater than the mean activity of the seronegative donors and greater than the no antibody control. Percent SR, LU20, and ADCC titers were used since not all of the antibody present in an individual's serum is capable of mediating functional ADCC activity.
Statistical analysis
Percent SR and LU20 values of all serum samples at each antibody dilution were calculated. Means and medians±standard deviations or standard errors (SD, SE) of % SR and LU20 values from each cohort were obtained. A positive ADCC response was assigned to samples with an activity >3 SD above the mean of our seronegative subjects. After determination of gp120-specific antibody titers, differences between groups were assessed using a Student's two-tailed t-test and confirmed using Mann–Whitney. Only one individual from each cohort had no ADCC response. Their data were included in the statistical analysis. When simultaneously comparing between and within groups, two-way ANOVA measures were used. Relationships between groups were assessed using the Pearson's correlation. In all statistical analysis, results were considered significant when the p value was <0.05. In this study, all statistical analysis and graphing were done using GraphPad Prism v.5 software (La Jolla, CA).
Results
ADCC antibody titers against HIV-1gp120 in infected men and women
The demographic and clinical characteristics of matched participants are described in Table 1. The mean absolute CD4+ T cell counts of both men and women were greater than 500 cells/μl and the log of the viral load was <4.0. Two-tailed Student's t-tests between men and women revealed no significant differences in serum ADCC activity when results were calculated as % SR (p=0.97) or LU20 (p=0.87) (Fig. 1). In fact, these results were remarkably similar, which makes it likely that comparable results would be seen in a similar comparison even if the sample size were considerably larger. Furthermore, there were no variations between men and women in % SR or LU20 values at any of the serum dilutions evaluated (p>0.1 in all two-tailed Student's t-tests and Mann–Whitney tests). Both groups had high ADCC antibody titers of 10,000 or 100,000 against HIV-1 gp120 when compared to seronegative controls, with the exception of two seropositive individuals (one from each cohort), who by definition have antibodies to HIV, but had an HIV-1 gp120 ADCC antibody titer of 0, indicating that no functional ADCC antibody activity was detected (Table 2).
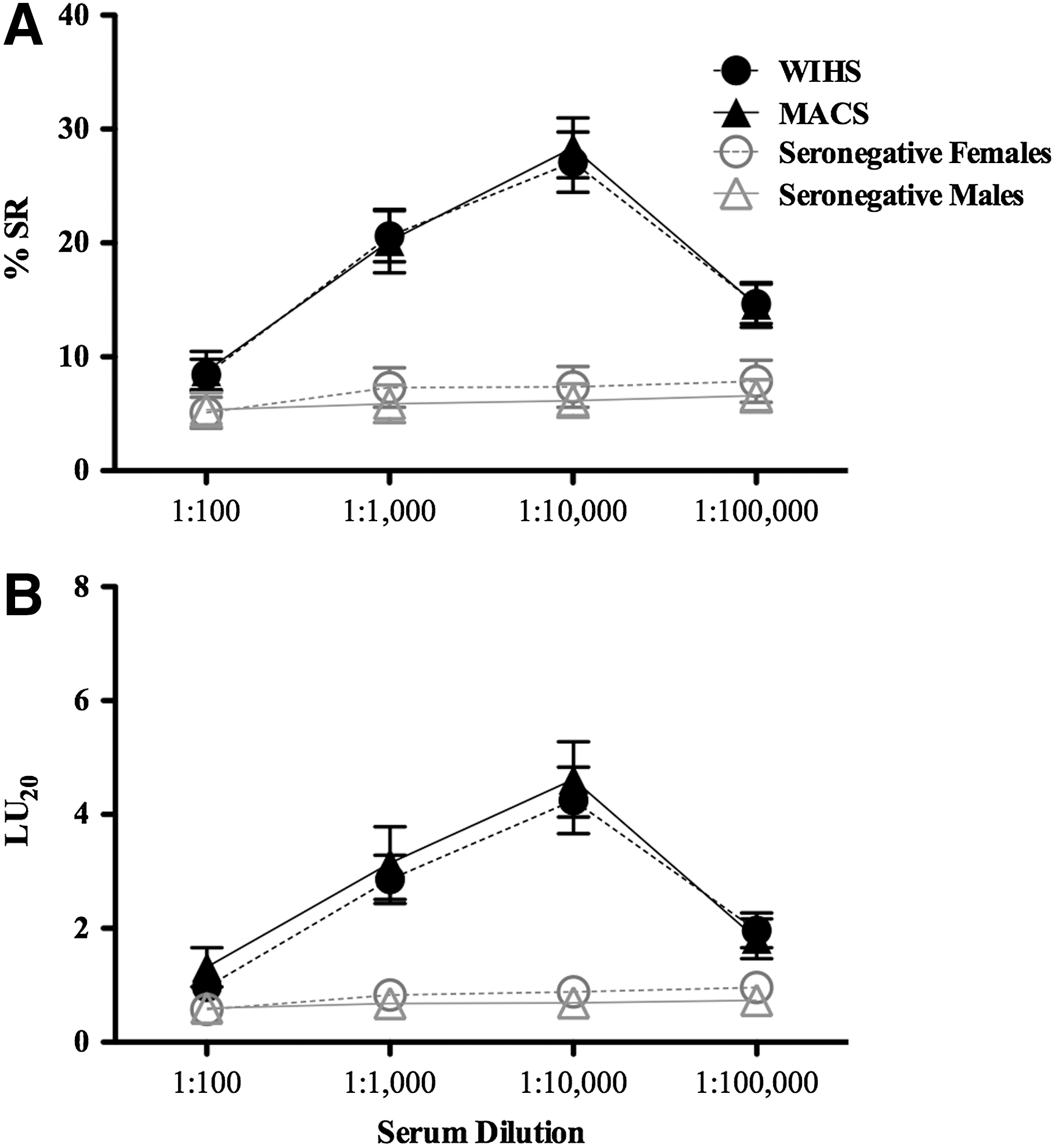
HIV-1-gp120 serum antibody-dependent cell-mediated cytotoxicity (ADCC) antibodies in untreated asymptomatic HIV-1-infected men and women. Fresh ADCC effector cells from uninfected healthy donors were incubated for 3.5 h with 51Cr-labeled HIV-1-gp120 bearing CEM.NKR target cells in the presence of serum (10-fold serial dilutions) from HIV-1-infected men (n=32) from the Multicenter AIDS Cohort Study (MACS), women (n=32) from the Women's Interagency HIV Study (WIHS), and from uninfected donors (n=5 males/5 females) who were not participants from either the MACS or WIHS cohorts. Each effector:target cell ratio (E:T) and serum dilution was assayed in quadruplicate.
Antibody-Dependent Cell-Mediated Cytotoxicity Serum Antibody Titers of Men and Women from the Multicenter AIDS Cohort Study and Women's Interagency HIV Study
Study participants from the Multicenter AIDS Cohort Study (MACS) and Women's Interagency HIV Study (WIHS) cohorts (n=32/32) were evaluated for antibody-dependent cell-mediated cytotoxicity (ADCC) against HIV-1 gp120-coated CEM.NKR targets in a standard 51Cr-release assay. ADCC antibody titers were determined based on percent specific release values. The number and percentage of observed ADCC antibody titers in men and women are depicted. ADCC antibody titers of 100 and of 1,000 were not observed among subjects from either cohort.
Association of ADCC antibody titer with CD4+ T cell number and HIV-1 viral load
For our study groups, we selected infected individuals who were asymptomatic and not on drug treatment. This eliminated variables that could be responsible for differences in ADCC activity between men and women, but it also limited our ability to characterize differences early after infection or later in disease progression because it provided a group of study participants who had a narrow range of CD4+ T cell numbers and viral loads. As expected, Pearson's correlations between ADCC antibody titers and CD4+ T cell number were not statistically significant when samples were matched for age, race, and CD4+ T cell number in either sex-based group [(r=0.03, p=0.87), (r=−0.09, p=0.61), men and women, respectively]. Nor did ADCC antibody titer show a relationship to HIV-1 viral load in men or women, respectively [(r=−0.21, p=0.25), (r=0.11, p=0.56)]. When we stratified our population by CD4+ T cell number (400–500 cells/μl, 500–600 cells/μl, 600–700 cells/μl, and >700 cells/μl), we did not find an association between CD4+ T cell number and ADCC antibody titer for men or women (Fig. 2). Two-way ANOVA measures between the sexes at two different viral load categories (0–4,000 copies/ml and >4,000 copies/ml) also revealed no correlation of titer with protection and no difference in this correlation between men and women (p>0.5 in all tests). Since only one man and one woman had no functional ADCC activity it was not possible to determine whether the presence of ADCC activity against HIV correlated with the number of CD4+ T cells or viral load.
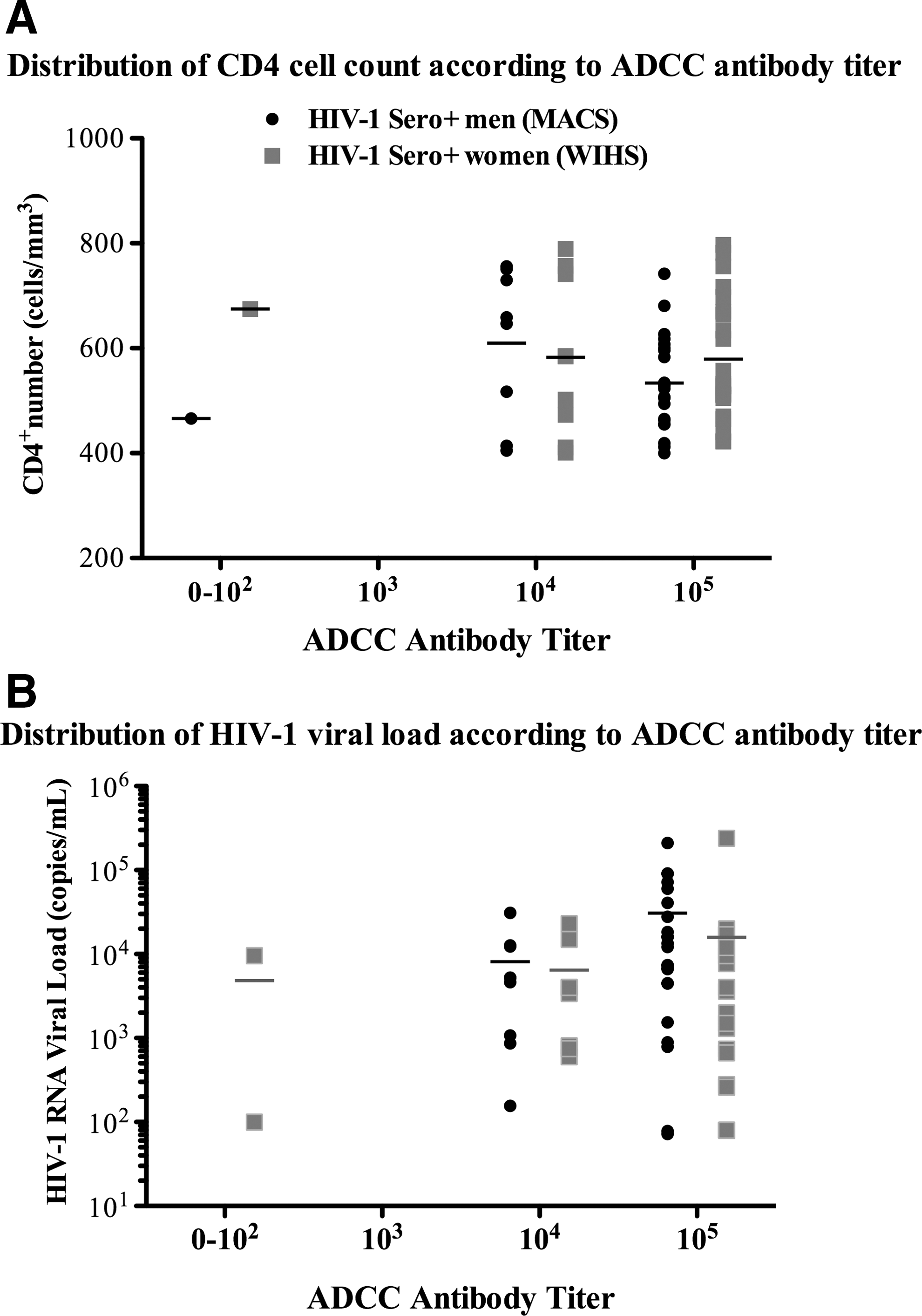
Distribution of HIV-1 immune markers according to detected antibody-dependent cell-mediated cytotoxicity (ADCC) titers in men and women. ADCC antibody titers were determined from percent specific release (% SR) values in a standard 51Cr-release ADCC assay using effector cells from healthy HIV-1-negative donors, gp120 coupled CEM.NKR targets, and 10-fold serial dilutions of serum as the source of antibodies. Serum samples from HIV-1+ men (n=32) from the Multicenter AIDS Cohort Study (MACS) and HIV-1+ women (n=32) from the Women's Interagency HIV Study (WIHS), all asymptomatic with no antiretroviral treatment (ART) were compared. Scatter plots and mean are depicted.
ADCC cross-clade reactivity in HIV-1-infected men and women
Cross-clade ADCC activity was defined as ADCC activity demonstrated by an individual who is infected with one clade of HIV-1, against targets coated with HIV-1 gp120 of another clade. We wanted to determine whether participants of this study, who were infected in the United States and other countries where clade B is the predominant clade of HIV, 21 –23 had any cross-reactivity against a different clade of HIV, clade A/E, and if this immune function was influenced by the sex of the donor. We, therefore, evaluated cross-clade reactivity in all the HIV-1-positive serum samples of the cohorts of this study (m/f=32/32) against gp120 MN (clade B) and gp120 CM (clade A/E)-coated targets. Combined male and female serum samples from clade B HIV-1-infected individuals (n=64) had significantly high ADCC responses against both clades of gp120 when compared to the seronegative controls (Student's t-test, p<0.0001 for % SR in both clades; p=0.03 and p=0.0096 in LU20 for clade B and clade A/E, respectively) (Fig. 3A). ADCC activity against clade A/E was slightly less than clade B activity (Student's t-test, p=0.01 and p=0.001, % SR and LU20, respectively) (Fig. 3A). When comparing groups of men and women, cross-clade reactivity in HIV-1-seropositive men was not significantly different from cross-clade reactivity in HIV-1-seropositive women, although this activity was somewhat lower in the female cohort (ANOVA, p=0.057 and p=0.5, % SR and LU20, respectively) (Fig. 3B). As expected, control seronegative sera (m/f=5/5) did not have ADCC activity against either gp120 clade.
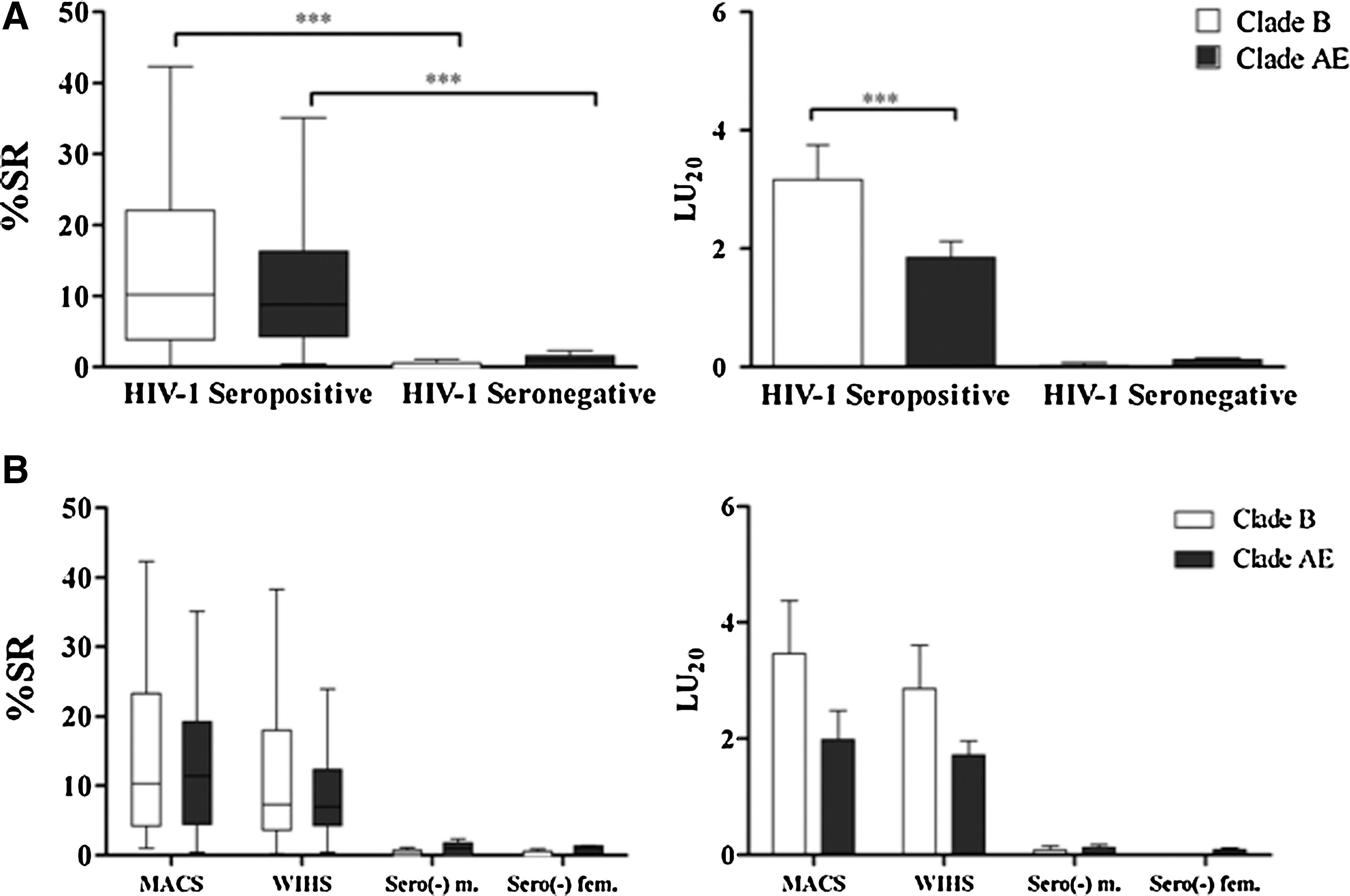
Cross-clade gp120 antibody-dependent cell-mediated cytotoxicity (ADCC) reactivity in men and women. Serum samples from HIV-1+ men (n=32) from the Multicenter AIDS Cohort Study (MACS) and from HIV-1+ women (n=32) from the Women's Interagency HIV Study (WIHS), all asymptomatic and under no antiretroviral (ART) treatment, were incubated at their ADCC optimal dilution for 3.5 h with 51Cr-labeled CEM.NKR target cells bearing either HIV-1-gp120MN (clade B) or HIV-1-gp120CM (clade A/E) in the presence of effector cells from healthy donors. Serum samples from HIV-1− healthy donors, not participants from the MACS or WIHS cohorts (n=5 males/5 females), were used as controls.
Discussion
Earlier studies came to conflicting conclusions about the differences between men and women with respect to HIV-1 infection and progression to AIDS. 25 –28 However, differences between HIV-1-specific ADCC antibodies in men and women have not previously been systematically compared. It is important to clarify these findings so that we will know that a vaccine that stimulates the production of ADCC antibodies will protect men and women equally. In the past, several studies have assessed the significance of ADCC antibodies in host defense against HIV-1-infected cells; 14,29 –31 however, the purpose of these studies was to determine whether there are differences between men and women in HIV-1 gp120-specific antibody titers responsible for ADCC activity. To accomplish this, we obtained serum samples from asymptomatic HIV-1-infected individuals matched for CD4+ cell number, HIV-1 viral load, age, and ethnicity and evaluated their ADCC activity (Table 1). Our results suggest that in matched populations of asymptomatic HIV-1-positive men and women with relatively high CD4+ T cell numbers, there were no differences in the concentration or the functional activity of HIV-1 gp120-specific antibodies that mediate ADCC, assessed either by % SR or LU20 (Fig. 1).
In previous studies we demonstrated that ADCC antibody titer correlates positively with CD4
One man and one woman had no functional ADCC activity even though they had serum antibodies against HIV. The presence of antibodies directed toward HIV in serum does not necessarily mean that these antibodies will have functional ADCC activity, since not all HIV-specific antibodies or even all neutralizing antibodies have ADCC activity. 33 This is not the first time that we have observed or reported seeing HIV-positive individuals who do not have serum ADCC activity. In our studies of paired serum and CVL samples in DATRI 009, we observed that a small percentage of HIV-positive women had ADCC antibodies in their genital fluids but not in their serum. 15
We also evaluated these 64 HIV-1 clade B-infected asymptomatic individuals for ADCC activity against target cells bound with either clade B (homologous) or clade A/E (heterologous) HIV-1 gp120 (Fig. 3A). The studies presented in Fig. 3 evaluated each sample at the serum dilution that had the best activity in our initial experiments. Men and women had comparable ADCC antibody responses to homologous clade B HIV-1. As we anticipated, clade B HIV-1-infected individuals had notably higher ADCC activity against clade B, HIV-1 gp120 MN, than they had against clade A/E, HIV-1 gp120 CM; it is noteworthy that the activity was reduced by only 16%. These results suggest that HIV-1 gp120-specific antibodies in the serum of infected individuals in this study had cross-reactive ADCC antibodies at asymptomatic stages of infection. These data are consistent with a previous report of cross-clade ADCC activity in HIV-infected individuals 34 and extend this observation, showing that sex does not have an effect on cross-clade reactivity.
To mediate ADCC, the ADCC antibodies must bind to epitopes of the virus that are expressed on the surface of infected cells and have an Fc region that will bind to the Fc receptor of ADCC effector cells. Recent studies in primates demonstrate that ADCC antibodies arise early after infection, 35 while neutralizing antibodies, which are generally clade specific, arise later in HIV-1 disease progression. Macaque studies indicate that ADCC antibodies have a broader range of activity than neutralizing antibodies. 36
Our studies confirm the cross-clade reactivity capabilities of ADCC antibodies and extend this observation to ADCC antibodies in humans. Furthermore, our results show evidence that in both men and women, serum antibodies from clade B HIV-1-infected individuals have cross-reactivity to clade A/E HIV-1. An HIV-1 vaccine should have broad cross-clade reactivity; consequently, it is important to understand the relative likelihood of generating cross-clade reactivity with new HIV-1 vaccine candidates and to identify what contributions they make to a protective immune response. These findings demonstrate that men and women behave similarly in response to natural infection and suggest that they would respond to vaccination with similar HIV-specific ADCC activity. If ADCC antibodies to HIV are considered as a target of vaccine development, this should be confirmed in the context of a vaccination. To our knowledge, this study is the first to compare ADCC and cross-clade ADCC reactivity to HIV-1 in men and women and to show that a vaccine that stimulates the production of protective ADCC antibodies would likely protect men and women equally.
Footnotes
Acknowledgments
Mariana M. Mata was the primary author of this article, analyzed data, and contributed to experimental work. Joyce R. Iwema contributed to data analysis and presentation and experimental work. Shanna Dell and Leslie Neems contributed to the experimental work. Beth D. Jamieson and John Phair contributed samples from the MACS and edited the manuscript. Mardge H. Cohen and Kathryn Anastos contributed samples from the WIHS and edited the manuscript. Linda L. Baum conceived and supervised all aspects of this project including the writing. Thanks to Fareeha Mahmood and Shohreh Raeisi for general laboratory assistance. Thanks to Ryan T. Sowell for critical reading of the manuscript. The Multicenter AIDS Cohort Study (MACS) and the Women's Interagency HIV Study (WIHS) provided the samples studied in this article. The WIHS is funded by the National Institute of Allergy and Infectious Diseases (UO1-AI-35004, UO1-AI-31834, UO1-AI-34994, UO1-AI-34989, UO1-AI-34993, and UO1-AI-42590) and by the Eunice Kennedy Shriver National Institute of Child Health and Human Development (UO1-HD-32632). The MACS is funded by the National Institute of Allergy and Infectious Diseases, with additional supplemental funding from the National Cancer Institute [UO1-AI-35042, UL1-RR025005 (GCRC), UO1-AI-35043, UO1-AI-35039, UO1-AI-35040, UO1-AI-35041]. Additional support was provided by PO1 AI082971 and the Chicago Developmental Center for AIDS Research (D-CFAR) P30 AI082151. The contents of this publication are solely the responsibility of the authors and do not necessarily represent the official views of the National Institutes of Health.
Author Disclosure Statement
No competing financial interests exist.