
Abstract
Definitive treatment of HIV infection remains a critical but elusive goal, with persistence of residual virus even in the face of prolonged administration of suppressive combination antiretroviral treatment (cART) providing a source for recrudescent infection if treatment is stopped. Characterization of the residual virus and devising strategies to target it for eradication are key goals in HIV treatment research. Indian rhesus macaques (In-RM) infected with SIVmac have been widely used in such research. However, it has proven challenging to achieve and sustain clinically relevant levels of suppression (<30 vRNA copies/ml plasma) with cART in such models. As ease of viral suppression by cART is related to pretreatment levels of viral replication, and levels of replication of SIVmac239/251 are lower in Chinese rhesus macaques (Ch-RM) than in In-RM, we evaluated cART administration to SIVmac-infected Ch-RM as a potential model for studies of residual virus and eradication strategies. Four SIVmac239-infected Ch-RM received cART including reverse transcriptase inhibitors PMPA/FTC and integrase inhibitor L-870812 daily for 8 weeks. Plasma viral loads were promptly reduced to <30 copies/ml upon initiation of cART. Cell-associated SIV DNA levels in lymphocytes from the gut were also significantly reduced. Jejunal and colonic CCR5+CD4+ mucosal memory T cells increased significantly; restoration of these cells was associated with reductions in immune activation. In conclusion, cART effectively suppressed viral replication to <30 vRNA copies/ml in SIVmac239-infected Ch-RM, reducing immune activation and restoring mucosal immune cell populations. SIVmac239-infected Ch-RM may be a useful model for studying responses to cART and persistent tissue reservoirs and evaluating candidate eradication strategies to cure HIV infection.
Introduction
P
Chinese rhesus macaques (Ch-RM) are one of the subspecies in the family of Macaca mulatta. The SIV-infected Ch-RM has been successfully used as a model for HIV/AIDS, and the dynamics of SIVmac replication in this model more closely mimic those of HIV-infected patients, as compared to SIVmac infection in Indian-origin macaques. 13 –15 Ch-RM infected with SIVmac generally develop AIDS more slowly than Indian rhesus macaques (In-RM) and other nonhuman primates that are susceptible to pathogenic SIV infection, 13,16 –18 more closely paralleling human HIV-1 infection. As the ease of suppression of viral replication in SIV-infected macaques is typically inversely related to pretreatment viremia levels, we reasoned that the results of administration of cART to SIV-infected Ch-RM might better resemble those of HIV-1 patients on cART, and provide a more tractable model to study viral suppression, immune restoration, and/or immune activation during antiretroviral treatment.
While residual virus has been documented in a variety of different tissues from individuals receiving cART, gut-associated lymphoid tissue (GALT) appears to be a prominent site. 3 Virus is also found in GALT in untreated long-term nonprogressing SIV-infected Ch-RM, 16 which suggests that the objectives of cART should include suppression of viral replication in the gut.
The combination of integrase inhibitors with standard highly active antiretroviral therapy potently suppresses HIV-1 replication. 19 –21 Here we evaluated a combination of two reverse transcriptase inhibitors with an integrase inhibitor monitoring longitudinal virologic and immunologic changes in blood and in the gut mucosa. Immune restoration and activation were compared with SIV-naive, long-term nonprogressors (LTNP), and normal progressors. The results suggest that cART administration to SIVmac-infected Ch-RM may be a useful model for the evaluation of new antiretroviral therapies, assessment of residual virus in the face of suppressive cART, and testing of strategies to eradicate this residual virus.
Materials and Methods
Animals and virus inoculation
Four rhesus macaques of Chinese origin (Macaca mulatta) were used. All animals were housed at the Tulane National Primate Research Center (TNPRC) and maintained in accordance with the standards of the American Association for Accreditation of Laboratory Animal Care and the “Guide for the Care and Use of Laboratory Animals” prepared by the National Research Council. All studies were reviewed and approved by the Tulane Institutional Animal Care and Use Committee (IACUC). All animals were seronegative for SIV, simian D retrovirus, and simian T cell leukemia virus prior to SIV inoculation. All animals were intravenously inoculated with 100 TCID50 of SIVmac239. DT92 and FB04 were infected for over 5 and 4 years, respectively. DV66 and EL50 were infected for over 13 months. All animals were in the chronic phase of infection.
Antiretroviral therapy
Each animal was treated with 20 mg/kg of (R)-9-(2-phosphonylmethoxyypropyl)adenine (PMPA, tenofovir) and 30 mg/kg of β-2′,3′-dideoxy-3′-thia-5-fluorocytidine (FTC, emtricitabine) daily by subcutaneous injection. Each animal was also simultaneously treated with 10 mg/kg of integrase inhibitor L-870812 orally twice a day by mixing the powder in Flavor-X syrups for voluntary consumption. The combination retroviral therapy was initiated at 5 or 4 years for animals DT92 and FB04 and at 13 months postinfection for animals DV66 and EL50. The treatment was continued for 8 weeks thereafter. PMPA and FTC were generously provided by Gilead Sciences, Inc. (Foster City, CA) and L-870812 was generously provided by Merck Research Laboratories (West Point, PA) via Material Transfer Agreements.
Lymphocyte isolation from blood and intestinal tissues
Blood and intestinal tissue samples were collected from jejunum and colon before cART, at weeks 2, 4, 6, and 8 during cART. Peripheral blood mononuclear cells (PBMCs) were isolated from fresh EDTA-anticoagulated blood. PBMCs were separated from EDTA-blood by Ficoll density gradient centrifugation. Gut tissues were obtained via endoscopic biopsies. Isolation of lamina propria lymphocytes from jejunum (LPLjej) and colon (LPLcol) tissues has been described elsewhere. 13,16 Briefly, intestinal lymphocytes were isolated using EDTA/collagenase digestion and Percoll density gradient centrifugation. First, biopsies were washed with 5% RPMI for 10 min, then incubated in HBSS containing 5 mM EDTA for 30 min. Cells were then digested with 60 U/ml collagenase (type II, Sigma, St. Louis, MO) for 40 min. To enrich and purify lymphocytes, cells were layered on discontinuous 40%/60% Percoll gradients (Sigma, St. Louis, MO) or 95% Ficoll-paque plus was used and centrifuged for 30 min at 1,000×g, washed, and resuspended in complete RPMI media containing 5% fetal calf serum (FCS) for further use. Due to the limited amount of cells obtained from biopsies, cells were used only for flow cytometry. Cells were unavailable for viral DNA quantification in each animal at certain time points during therapy.
Antibodies, immunofluorescent staining, and flow cytometry
PBMCs, LPLjej, and LPLcol were stained simultaneously for T cell immunophenotyping, proliferation, and activation with the following fluorescently conjugated monoclonal antibodies from BD: CD3–Pacific Blue (SP34), CD8–PE-Texas Red (MHCD0817), CD4–FITC (L200), CD95–PE-Cy5 (DX2), CD28–APC (28.2), CCR5–PE (3A9), and HLA-DR-PE-Cy7 (L243). CD8-Qdot655 (M-T807) was obtained from the NIH NPRR courtesy of Dr. K. Reimann (Harvard University, Cambridge, MA). Cells were stained for 30 min at 4°C in the dark and washed twice with phosphate-buffered saline (PBS) containing 2% FCS; cells were fixed with BD Stabilizing Fixative Buffer. Samples were analyzed using a FACSAria flow cytometer. Cells were gated through CD3+ T lymphocytes, and at least 20,000 events (CD3+ T cells) were collected per sample.
Quantification of plasma viral RNA and total cellular SIV DNA
SIV plasma viral loads (pVL) were analyzed by standard real-time PCR (qRT-PCR) 22 for the longitudinal monitoring after SIV infection but before cART and an ultrasensitive quantification qRT-PCR was used to confirm pVL before cART, and used thereafter during the entire course of therapy. 22 The limit of detection was 30 copies vRNA/ml. For cell-associated SIV DNA quantification, total cellular genomic DNA from PBMCs, and lymphocytes isolated from jejunum and colon were extracted using the DNeasy Blood & Tissue Kit (Qiagen Inc., Valencia, CA). A real-time TaqMan PCR assay was used as previously described. 16 Briefly, approximately 100 ng of total DNA was used and mixed with PCR master mix in a total of 50 μl and subjected to 40 cycles of quantitative real-time PCR analyses. Fluorescence signal was detected with an ABI Prism 7900HT Fast Real-Time PCR System. Data were captured and analyzed with Sequence Detector Software (Applied Biosystems, Carlsbad, CA). Viral copy number per 1×106 cells was determined and calculated by plotting C T values obtained from samples against a standard curve generated with in vitro transcribed RNA representing known viral copy numbers and was calculated with RNase P TaqMan Copy Number Reference Assay for cell number count (Life Technologies Corporation). The limit of detection was 200 copies/106 cells equivalent.
Statistical analyses
Data were analyzed by repeated-measures ANOVA followed by Bonferroni's posttest for comparison between the different time points during cART. A nonparametric statistical analyses Mann–Whitney test was used for comparison between different compartments. A paired t test was used to compare levels of SIV-targeted T cells (memory CD4+CCR5+ T cells) between jejunum and colon. The Spearman correlation was used to assess the relationship of levels of SIV-target cells with CD4+ central or CD4+ effect memory T cells in jejunum or colon, as well as this population between colon and jejunum. GraphPad Prism 4.0 statistical software was used to analyze data and statistical significance was set two-sided at p<0.05.
Results
Dynamics of plasma viral loads before and during combination antiretroviral therapy
We studied cART administration in four animals chronically infected with SIVmac239 (Fig. 1). Animals DV66 and EV50 had been infected for 52 weeks before cART. DV66 had plasma viremia between 103 and 104 copies/ml and EL50 had between 105 and 106 copies/ml plasma before initiation of cART. DT92 and FB04 had been infected over 5 and 4 years, respectively. Both had peak viremia 2 weeks post-SIV infection, but had achieved control of viremia to <1,000 copies/ml after 4 weeks of infection. DT92 had sustained plasma viral load around the level of 103 copies/ml for 8 weeks prior to initiation of cART, whereas FB04 maintained levels below 125 copies/ml but had a blip of 180 copies/ml 4 weeks prior to cART. We included these two animals to examine changes in viral levels in tissues such as the GALT, one of the major reservoirs of HIV/SIV, and also to examine immune restoration and activation during cART. After initiation of cART, all four animals demonstrated rapid decreased viremia within 2 weeks, and three animals were suppressed to less than 30 copies/ml within 7 days of therapy. Only DT92 had a blip (160 copies/ml) at 42 days of therapy, and all other animals maintained less than 30 copies/ml throughout the 60 days of cART. Animal EL50 was euthanized at day 19 of cART due to colitis. Even though this animal had a high pVL before therapy, it suppressed the pVL to less than 30 copies/ml within 14 days of cART.
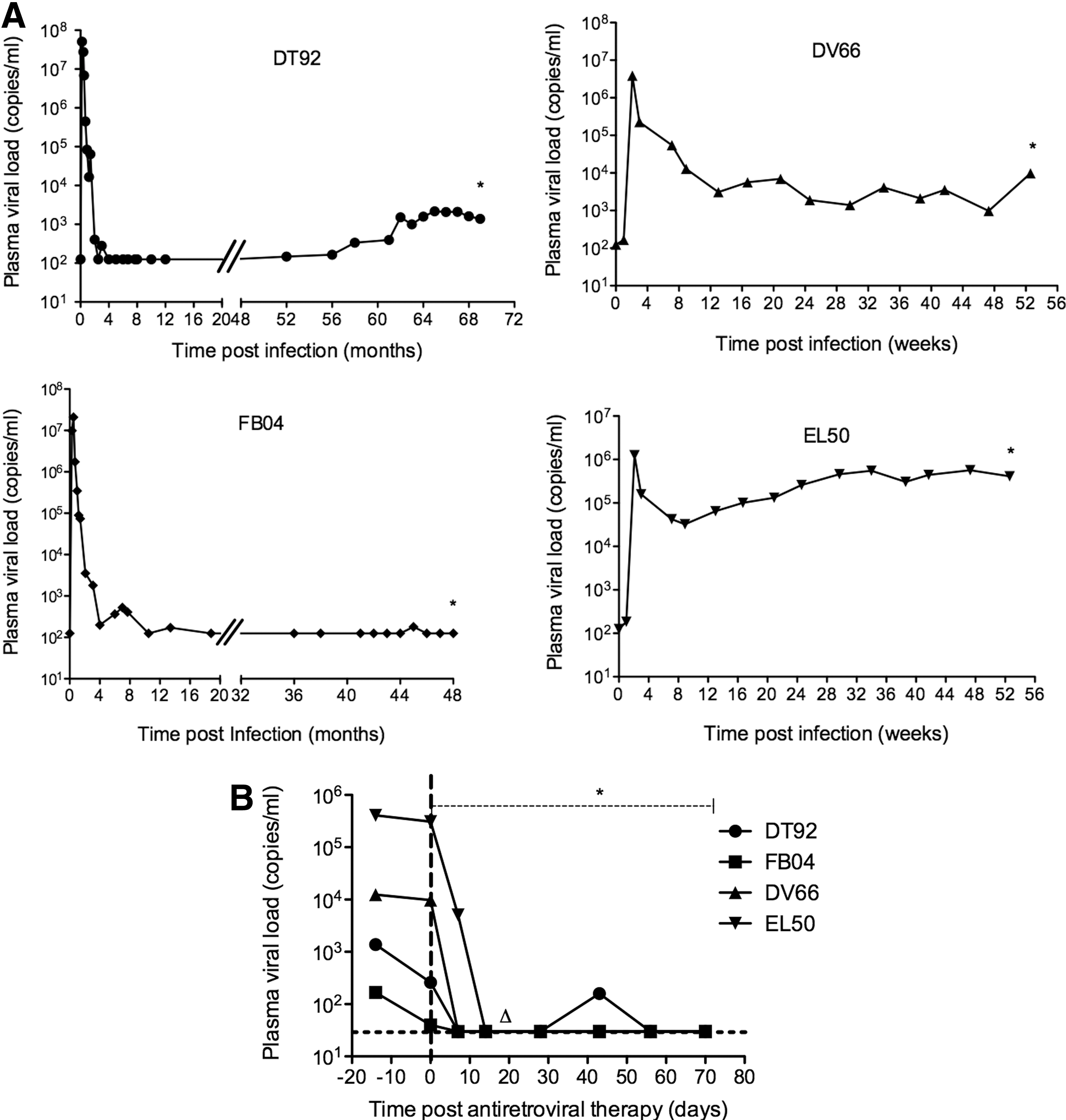
Plasma viral load measured longitudinally for each animal before
Total cell-associated SIV DNA load in different tissues throughout the course of combination antiretroviral therapy
To monitor dynamic changes in levels of cell-associated SIV DNA in peripheral blood, jejunum, and colon, viral DNA was quantified by real-time PCR before cART and every 2 weeks during cART (Fig. 2) with the exception of DT92 for which few cells were available from colon at 2 weeks of cART and from jejunum at 6 and 8 weeks of cART. FB04 had few cells available from jejunum biopsy at 2, 6, and 8 weeks of cART or colon biopsy at 8 weeks of cART, and DV66 had few cells from colon sample at 8 weeks of cART. EL50 had intestinal data for pre-cART and 2 weeks of cART but no other time points.
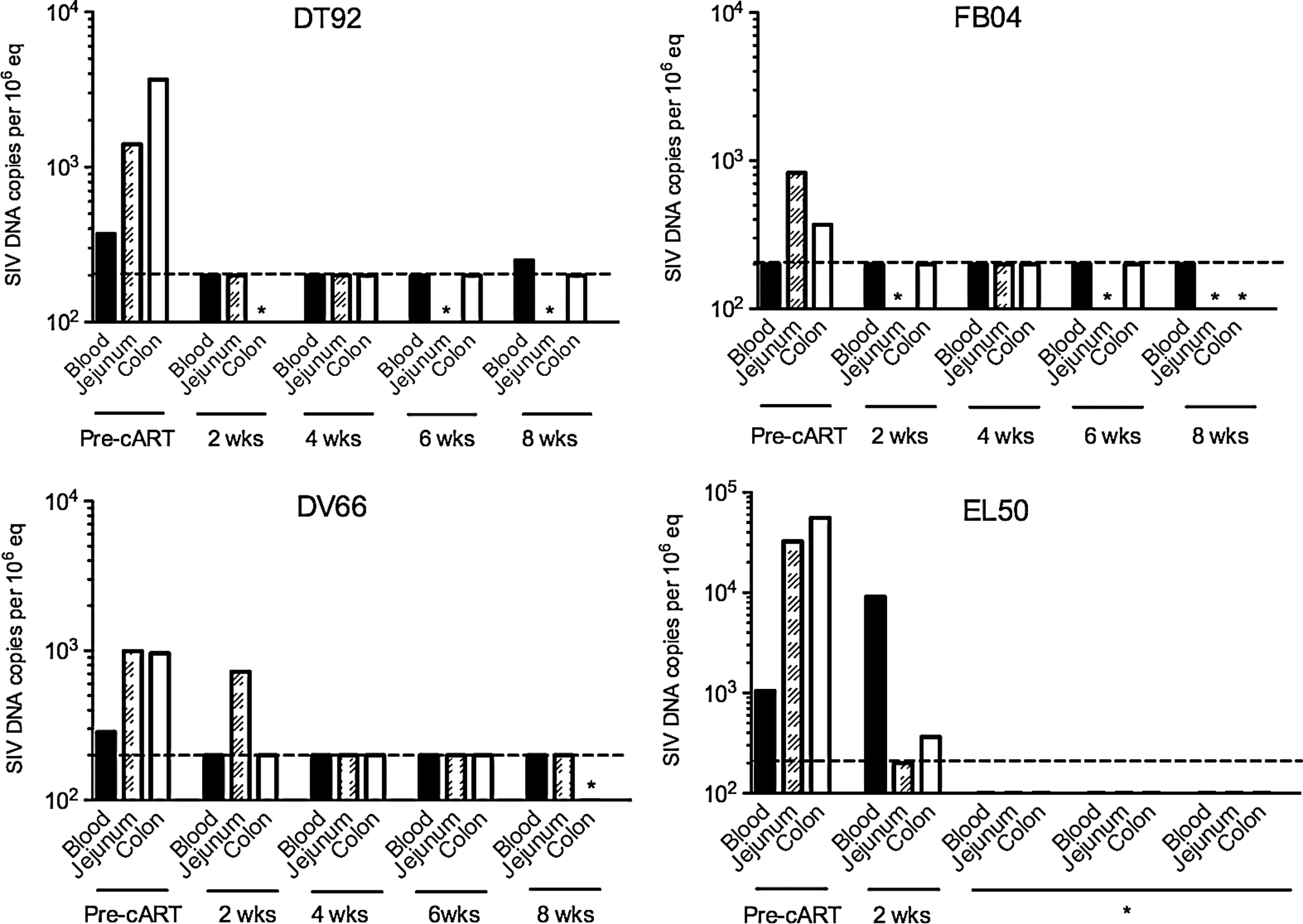
Levels of cell-associated simian immunodeficiency virus (SIV) DNA in peripheral blood mononuclear cells (PBMCs) from peripheral blood and lamina propria lymphocytes from the jejunum and colon in individual animals before cART and at 2, 4, 6, and 8 weeks of cART. *Data were not available. Dashed line indicates the limit of detection at 200 copies/106 cells equivalent.
Animals had variable levels of SIV DNA in jejunum and colon. Except for FB04, all other animals had detectable SIV DNA in PBMCs as well (limitation of SIV DNA detection was 200 copies/106 cells). DT92 had 250 copies/106 cells in PBMCs at 8 weeks of cART, DV66 had 722 copies/106 cells in jejunum at 2 weeks of cART, and EL50 had 9,090 copies/106 cells in PBMCs and 364 copies/106 cells in colon. SIV DNA levels were below 200 copies/106 cells in tissues at all other time points such as 4 weeks and 6 weeks of cART. Combined, these results suggest that cART was effective in reducing SIV DNA load not only in peripheral blood but also in the gut during the 8 weeks of cART.
Dynamics of CD4+, CD8+ T cells and CD4/CD8 ratios in blood during combination antiretroviral therapy
Pretreatment percentages of CD4+ T cells were within the range of 35–48% in all animals. Upon initiation of treatment, levels increased by day 7 to 53–78% and levels were sustained above 40% thereafter through the first month of therapy. By day 56 of cART, the CD4 cells were within 55–62% in DT92, FB04, and DV66. Although animal EL50 responded to cART and increased percentages of CD4+ T cells to 62% by day 14 of cART (Fig. 3A), the absolute CD4+ counts were low (less than 200 cells/μl) prior to receiving cART, reflecting severe lymphopenia. This animal was euthanized at day 14 of cART because of severe colitis, which may not have been related to this study. The other three animals slightly increased and maintained CD4+ T cell counts during the period of cART. The ratios of CD4/CD8 were in the range of 0.5–0.8 before initiation of cART, but the ratios increased to almost 1.0 by the end of therapy (Fig. 3B).

Dynamics of CD4+ T cells (%) in peripheral blood during combination antiretroviral therapy
Restoration of CD4+ T cells in the gut during combination antiretroviral therapy
To assess the effect of cART on restoration of CD4+ T cells in the gut, we examined the dynamics of changes in total CD4+ T cells and CD4+ T cell subsets in lamina propria lymphocytes of the jejunum and colon. The percentages of CD4+ T cells in the gut markedly increased after 2 weeks of treatment, and levels were maintained higher than pretreatment for each animal (Fig. 4A). Interestingly, percentages of CD4+ T cells were always higher in the colon than that in jejunum for each animal. Percentages of CD4+ T cells increased from 4.3±2.4 at pre-cART to 17.6±6.2 at the end of cART in the jejunum and from 20±8.5 at pre-cART to 33.8±7.7 at the end of cART in the colon. Throughout the period of cART, there was a steady increase with significantly higher levels in the colon than that in jejunum (Fig. 4B). Further analysis of CD4+ T cell subsets showed that percentages of CD4+CD95+CD28+ T cells (central memory T cells, TCM) in the jejunum and colon were comparable, and these cells increased during treatment (Fig. 4C). In contrast, percentages of CD4+CD95+CD28− T cell subsets (effector memory T cells, TEM) decreased 7 days posttherapy, and remained at low levels in most animals. One exception was animal DV66 who had remarkably higher levels than others even before treatment, but nevertheless still had a similar trend as the others (Fig. 4D).
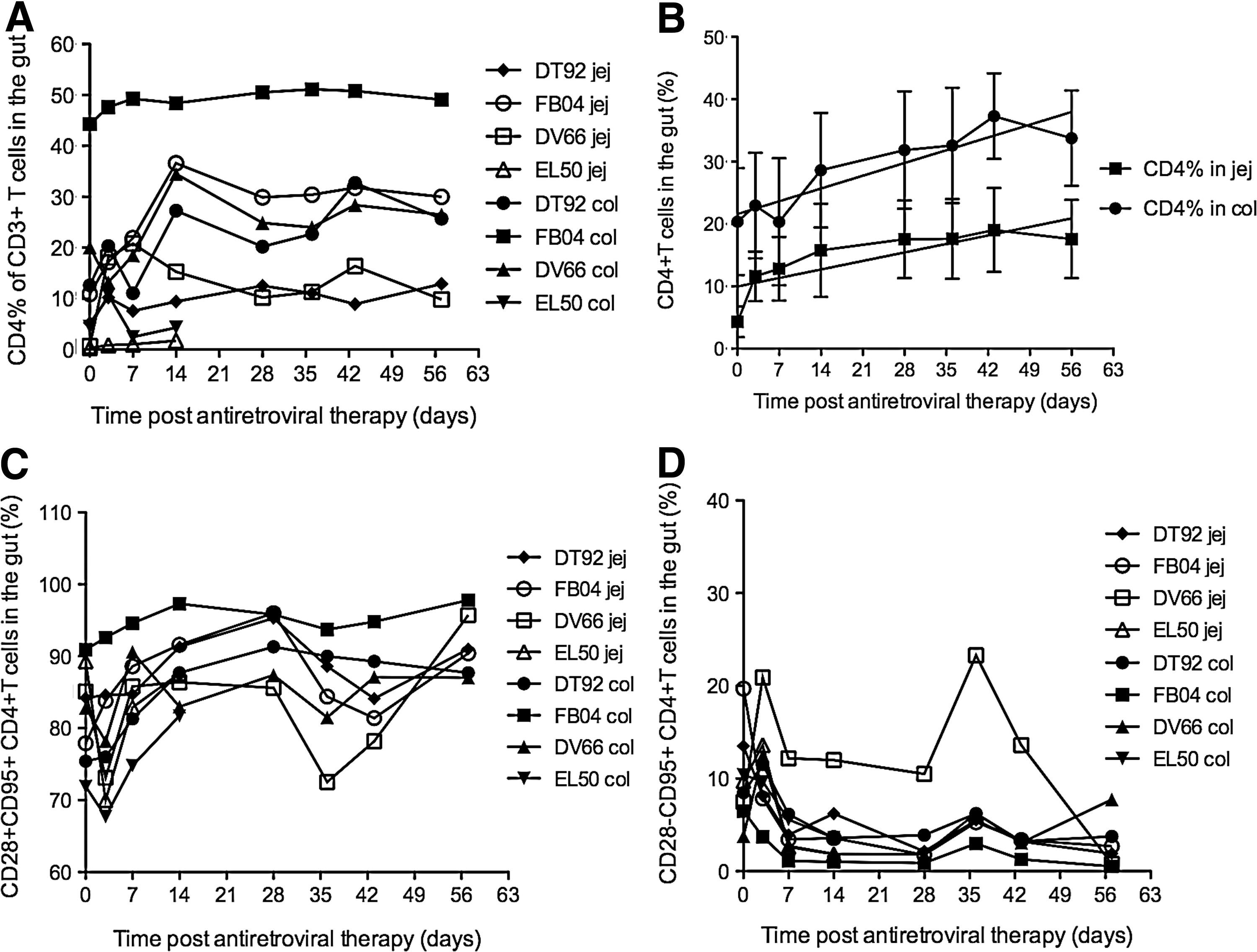
Dynamics of CD4+ T cells (%) in the jejunum and colon in each animal
Dynamics of memory CD4+CCR5+ T cells (SIV-target cells) in the gut during combination antiretroviral therapy
We previously reported that early restoration of memory CD4+CCR5+ T cells in the gut distinguishes long-term nonprogressors and normal progressors in SIV-infected Ch-RM. 17 To further investigate which population of CD4+ T cells changed during cART, we examined changes in memory CD4+CCR5+ T cells using a previously described index, 16 which reflects the proportion of total CD4+ T cells in the CD3+ T cell pool that are memory (CD95+) and coexpress CCR5. 16 These cells were also designated as SIV-target cells due to their expression of the HIV/SIV coreceptor CCR5. We observed that with cART all animals had a marked increase in this population in both the jejunum and colon (Fig. 5A). There was a strong positive correlation (p<0.001) between levels of target cells in the jejunum and colon, with a higher level in the colon (Fig. 5B). This result was consistent with our previous findings in LTNP. 16
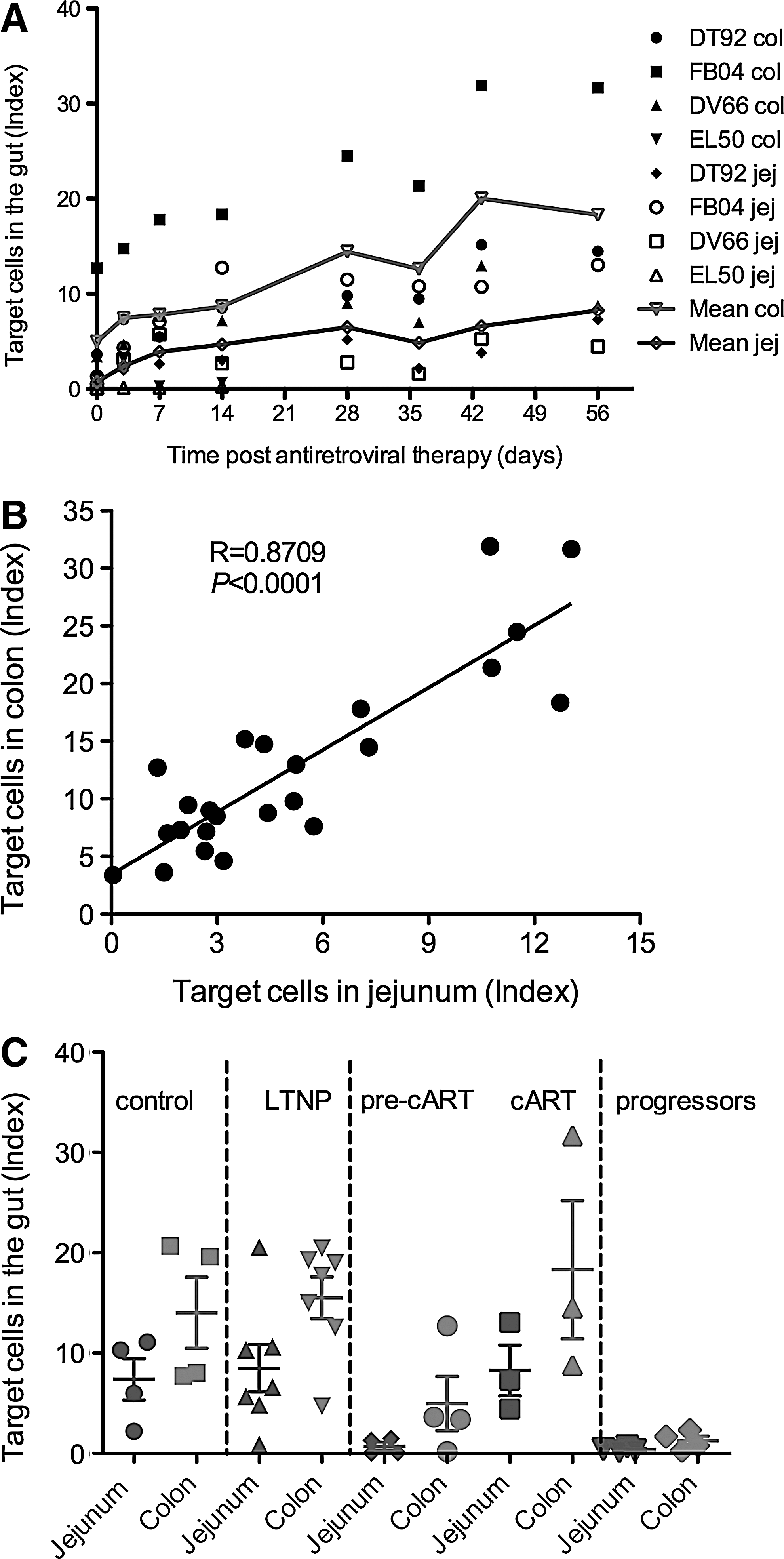
Longitudinal follow-up of changes of SIV-target cells (memory CD4+CCR5+ T cells) in the colon and jejunum during ART. The index reflects the proportion of total CD4+ T cells of the T cells pool (CD3+ T cells) that are CCR5+ and memory (CD95+) phenotype; this index has been described elsewhere.16 Lines (in red and blue) represent the mean index of SIV-target T cells in the colon and jejunum, respectively
To compare the restoration of target cells in cART animals with SIV-naive controls, LTNP, and progressors that was previously reported, 16 we found that in both compartments of the jejunum and colon, the cART macaques had levels similar to the SIV-naive controls and LTNP and also increased compared to the levels before cART (Fig. 5C), whereas progressors had significantly lower levels of these cell populations. These results suggest that animals receiving treatment with combination antiretroviral regimen were able to restore target cells to levels observed for LTNP and healthy SIV-uninfected macaques.
Association of SIV-target cells with CD4+ TCM and TEM in the gut
To investigate potential associations between SIV-target cells and CD4+ TCM or TEM cells, we performed a correlation analysis. In the jejunum, while the correlation of target cells with TCM was not significant (Fig. 6A), a significant inverse correlation with TEM (p<0.0101) was observed (Fig. 6B). In the colon, target cells had a strong positive correlation with TCM cells (p<0.0001) (Fig. 6C); in contrast, these cells had a significant inverse correlation with TEM cells (p<0.0002) (Fig. 6D) suggesting that these target cells are most likely within the population of CD4+ TCM T cells.

Association between SIV-target cells and CD4+CD95+CD28+ T cells and between SIV-target cells and effector CD4+ T cells in the jejunum
Immune activation in gut CD4+ T cells and CD8+ T cells during combination antiretroviral therapy
Chronic HIV infection is associated with chronic immune activation, which persists even in subjects on fully suppressive cART at levels above those seen in uninfected subjects. 23,24 To investigate whether there was any reduction of immune activation in the gut during cART, we examined activation of CD4+ and CD8+ T cells throughout cART by comparing their coexpression of HLA-DR. In the jejunum, CD4+ T cell activation decreased significantly after 14 days of cART and was sustained at a lower level (<5% of HLA-DR+CD4+ T cells) through the initial 35 days of cART. However, HLA-DR expression increased at day 42 of cART and was maintained at comparable levels through day 56 of cART (Fig. 7A). In the colon, activation of CD4+ T cells decreased after 14 days of cART and was maintained at low levels except at day 56 of cART.

CD4+ and CD8+ T cell activation determined by HLA-DR expression in the gut
In the jejunum, activation of CD8+ T cells was low before cART, remained low throughout day 42 of cART, and increased to 8% at day 56 of cART. In the colon, activation of CD8+ T cells was significantly higher than in the jejunum and had a peak of 17% at day 3 of cART but decreased thereafter until the end of cART. Both the jejunum and colon showed similar trends between day 7 and day 42 of cART. Activation of CD8+ T cells was also decreased or maintained below 10% by the end of cART except in the colon of DT92 (Fig. 6B). While no correlations were observed between activation of CD4+ or CD8+ T cells with target cells in the jejunum (Fig. 7C and D), inverse correlations were shown in the colon with activation of both CD4+ (p<0.0042) and CD8+ T cells (p<0.02) (Fig. 7E and F). In comparison with SIV-naive, LTNP, and progressors, activation levels in the cART group were comparable to LTNP and SIV-naive groups in both the jejunum and colon, and slightly decreased compared to pre-cART.
Discussion
We treated four Ch-RM with chronic SIVmac239 infection with a cART regimen consisting of PMPA/FTC/L-812. The four animals had varying levels of pVL prior to antiretroviral therapy. We included animals with both low pVL and higher pVL (1) to assess decay of tissue reservoirs (i.e., the GALT), as it is known that even when pVLs are undetectable in peripheral blood, replicating virus is consistently found in tissues such as GALT, 3,16,25,26 and (2) to evaluate immune restoration and activation, because even in LTNP, gut immune CD4+ T cells are not fully recovered to baseline levels. 17 Effective cART could help restore CD4+ T cells in tissues and reduce immune activation. cART was very effective in suppressing viral replication in these SIV-infected Ch-RM. Plasma viremia decreased to <30 copies/ml within 7–14 days of cART, even when pre-cART pVL was 104–106 copies/ml. In reports of the implementation of cART in SIV-infected In-RM and pigtailed macaques it can take months to reduce viremia to <100 copies/ml and low level spikes of measurable viremia “blips” may persist in some animals. 4,11,12, 27 Generally, Ch-RM have lower pVL than other NHPs after SIV infection, which is closer to those of HIV-1 patients. 13,14,18 Accumulating data indicate that the levels of viral replication, reflected by pVL prior to initiation of antiretroviral therapy, influence the facility of viral suppression with cART.
Depletion of gut CD4+ T cells is a hallmark of HIV/SIV pathogenesis and restoration of gut CD4+ T cells, especially restoration of CD4+ TCM, which are extremely important for maintenance of memory T cell homeostasis, is regarded as a therapeutic benefit as sustained depletion or dysfunction of this population eventually leads to the onset of AIDS. 28 –30 Combination ART can improve restoration of this population of CD4+ T cells in SIV infection as observed in the present study, as well as by others in other systems. 10 In some studies, TCM were defined slightly differently than here, as either CD28+CCR7+CCR5−, 31 CD95+CD62L+, 23 or CD28+CD95+. 32 We used the latter immunophenotyping, which may include the transitional effector memory cells, but this population is in general quite small. The correlation of TCM with CD4+CCR5+ SIV-target cells indicates that this population of immunologically critical cells represents a major compartment for persisting virus, complicating eradication strategies. 33,34
It is known that HIV-1 latency is rare with one infected CD4+ resting cell in a million CD4+ resting cells. 35 However, it is unclear whether restoration of total numbers of TCM cells proportionally increases the number of productively or latently infected TCM cells. It has been reported that even lymph nodes are enriched for memory CD4+ T cells when compared to the blood, and the frequencies of memory CD4+ T cells harboring proviral DNA were still comparable in the lymph nodes, blood, and gut, 33 indicating a lack of positive association between total TCM CD4+ T cells and latently infected TCM CD4+ T cells. It is also unknown whether there is a threshold for the size of the viral reservoir in cells that does not permit viral replication rebound (functional cure), although it is argued that theoretically one infected cell is all that may be required for viral replication to resume. Recent results demonstrate that naive T cells are dispensable for memory CD4+ T cell homeostasis, and CD4+ memory T cells are capable of self-renewal, 31 indicating that viral reservoirs may transit between TCM and TEM only, but virus is mainly derived from TCM as observed in this study and by others. 33 Further investigations are needed to elucidate whether virally infected TCM increase when the restoration of total CD4+CCR5+ memory T cells is improved by cART, which could provide insight into strategies to specifically eliminate infected TCM but preserve uninfected immunologically critical TCM cells.
We also found that the percentage of CD4+ T cells in the colon was higher than that in jejunum in all animals receiving cART, consistent with our previous findings in LTNPs. 16 Testing colon or rectum CD4+ restoration may serve as a useful parameter to assess efficacy of cART in general.
It is known that HIV-1 patients on effective cART have continuous chronic immune activation and inflammation that account for higher morbidity and mortality than HIV-uninfected people. 36 However, the causes contributing to the chronic immune activation are yet to be elucidated, although multiple factors such as residual HIV viremia, microbial translocation, and dysfunction of immunoregulatory responses may play important roles. 37 It is interesting that there were inverse correlations between immune activation and SIV-target cell restoration, suggesting that reduction of immune activation benefits the reconstitution of T cells (Fig. 6E and 6F). Of note, this phenomenon was observed only in the colon and not in jejunum. Considering the differences in anatomy and histology of these two sites, this is not completely unexpected, although caution is advisable in generalizing from the small number of animals studied here. Previously, we observed a positive correlation of SIV-target cells in the jejunum and colon in long-term nonprogressing Ch-RM 16 and here in Ch-RM on cART with a higher baseline level of target cells in the colon. It remains to be determined whether levels of virus in the colon can be significantly reduced, along with the decrease in immune activation by cART.
Taken together, and although the numbers of animals in this study are small, our results indicate that SIVmac239-infected Ch-RM may have some advantages compared to other currently used NHPs as a model for studying the virus that remains in the face of suppressive cART, and evaluating strategies for targeting this virus to achieve a functional cure or eradication.
Footnotes
Acknowledgments
We thank C. Lanclos and J. Bruhn of the immunology flow cytometry core laboratory, M. Duplantis and L. Li for tissue sample collection, and L. Doyle-Meyers and the animal care staff of the Department of Veterinary Medicine for their technical assistance. Research was supported by NIAID R01 AI093307-01A1 (BL), R01 AI084793 (RSV), a Tulane Research Enhancement grant (BL), and the National Center for Research Resources, and the Office of Research Infrastructure Programs (ORIP) of the National Institutes of Health through grant OD011104-51 and in part with federal funds from NCI/NIH contract HHSN261200800001E. The funders had no role in the study design, data collection and analysis, decision to publish, or preparation of the article.
Author Disclosure Statement
No competing financial interests exist.