
Abstract
Patients with HIV-1 and human T-lymphotropic virus type 2 (HTLV-2) coinfections often exhibit a clinical course similar to that seen in HIV-1-infected individuals who are long-term nonprogressors. These findings have been attributed in part to the ability of HTLV-2 to activate production of antiviral chemokines and to downregulate the CCR5 coreceptor on lymphocytes. To further investigate these observations, we tested the ability of recombinant Tax1 and Tax2 proteins to suppress HIV-1 viral replication in vitro. R5-tropic HIV-1 (NLAD8)-infected peripheral blood mononuclear cells (PBMCs) were treated daily with recombinant Tax1 and Tax2 proteins (dosage range 1–100 pM). Culture supernatants were collected at intervals from days 1 to 22 postinfection and assayed for levels of HIV-1 p24 antigen by ELISA. Treatment of PBMCs with Tax2 protein resulted in a significant reduction in HIV-1 p24 antigen levels (p<0.05) at days 10, 14, and 18 postinfection compared to HIV-1-infected or mock-treated PBMCs. This was preceded by the detection of increased levels of CC-chemokines MIP-1α/CCL3, MIP-1β/CCL4, and RANTES/CCL5 on days 1–7 of infection. Similar, but less robust inhibition was observed in Tax1-treated PBMCs. These results support the contention that Tax1 and Tax2 play a role in generating antiviral responses against HIV-1 in vivo and in vitro.
Introduction
A
Two retrospective case-controlled cohort studies investigating the clinical outcomes and survival probabilities among persons coinfected with HIV-1 and HTLV-1 or -2 showed that HTLV-2 coinfections resulted in improved survival and delayed progression to AIDS. 3,7 These findings have been attributed in part to the ability of HTLV-2 to activate production of antiviral chemokines and to downregulate the CCR5 coreceptor on lymphocytes. 9,10 Lewis et al. 9 reported that lymphocytes from individuals with HIV-1 and HTLV-2 coinfections spontaneously produce CC-chemokines. Recently, our group reported that Tax2 and Tax1 proteins induced the expression of three CC-chemokines with antiviral function MIP-1α/CCL3, MIP-1β/CCL4, and RANTES/CCL5 from peripheral blood mononuclear cells (PBMCs) and monocytes with the concomitant downregulation of CCR5, the HIV-1 entry coreceptor. 10,11 These results suggested that Tax2 and Tax1 could promote innate immunity in the extracellular environment during HTLV-2 and HTLV-1 infections via CC-chemokine ligands and receptors, and led us to hypothesize that Tax2 and Tax1 may directly or indirectly inhibit HIV-1 replication. In the present study, an in vitro cell culture system was established where HIV-1-infected PBMCs were treated with Tax2 or Tax1 proteins. The in vitro treatment of HIV-1-infected PBMCs with Tax2 or Tax1 resulted in a significant decrease of HIV-1 replication. These findings further implicate Tax1 and Tax2 as disease modifiers among coinfected individuals.
Materials and Methods
HIV-1 stock
HIV-1 stocks were derived from the infectious molecular clone pNL-AD8 12 and used to infect the Hut/CCR5 cell line; the culture supernatants were centrifuged at 1,500 rpm for 10 min, passed through a 0.45-μm filter, and stored at –80°C. Gag p24 concentrations of HIV-1NLAD8 stocks were measured using an enzyme-linked immunosorbent assay (ELISA; anti-p24-coated plates were purchased from the AIDS Vaccine Program, SAIC, Frederick, MD).
Recombinant Tax1 and Tax2 proteins
Recombinant Tax1 and Tax2 proteins were expressed and purified as recently described.
10,11
Briefly, transformed BL21(DE3) strains of Escherichia coli with pET-Tax1 or pET-Tax2 were grown at 18°C under ampicillin selection and induced using isopropyl β-
Peripheral blood mononuclear cells (PBMCs)
Blood samples were collected from three healthy donors after receiving written informed consent, in a protocol approved by the Institutional Review Board for Human Investigation of the Milwaukee Veterans Affairs, Research Service Committee. Donors were confirmed to be HIV-1 and HTLV-1/-2 seronegative by ELISA assay. Whole blood was collected in CPT/Vacutainer BD tubes (BD Biosciences, San Jose, CA), and PBMCs were obtained following the manufacturer's recommendations.
HIV infection and treatment with Tax
PBMCs (1×106/ml) were cultured overnight in complete RPMI-1640 [RPMIc, 10% fetal bovine serum (FBS), 1% penicillin-streptomycin-glutamine, 1% sodium pyruvate] medium containing 5 μg/ml phytohemagglutinin (PHA) (Sigma). The activated cells were centrifuged, washed, suspended in HIV-1NLAD8 virus stock (0.8–1.2 ng/ml of HIV-1 p24 equivalent virus), and incubated on a rocker platform for 2 h at 37°C to allow virus adsorption. The cells were washed thoroughly in RPMIc to remove unadsorbed virus, resuspended in RPMIIL-2 (RPMI-10% IL-2, 1% penicillin-streptomycin-glutamine, 1% sodium pyruvate), and aliquoted into 24-well plates, followed by the addition of recombinant Tax1, Tax2, or ebe without the Tax vectors as a control. In some experiments, the recombinant proteins were added immediately following infection with HIV-1, while in other experiments (Tax2 only) the recombinant protein was also administered 48 h prior to and at 48 h following HIV-1 infection. In both experiments, the recombinant proteins were then subsequently added daily during the entire incubation period. Recombinant Tax proteins (Tax2 and Tax1) were added in a range of 1–100 pM and each condition was tested in triplicate. Uninfected and untreated PHA-activated PBMCs were also included as controls. Cell culture supernatants were harvested at days 1, 3, 5, 7, 10, 14, 18, and 22 postinfection and virus replication was assessed for levels of HIV-1 p24 antigen by ELISA (AIDS Vaccine Program, SAIC, Frederick, MD).
Assay for determination of CC-chemokines in culture supernatant fluids
Supernatants harvested for p24 assays were reserved and subsequently assayed for MIP-1α, MIP-1β, and RANTES expression by specific ELISA using DuoSet ELISA development kits (R&D Systems, Minneapolis, MN) according to the manufacturer's specifications. Serial dilutions of known concentrations of the respective CC-chemokine were used in each assay to generate a standard concentration curve. Absorbance values at 490 nm were used to quantify the levels of chemokines in the culture supernatants from the standard concentration curve and CC-chemokine protein levels were expressed in picograms per milliliter (pg/ml).
Statistical analysis of data
Two-way repeated-measures (RM) ANOVA (within factor of time and between factor of treatments) was performed to assess the presence of differences between HIV-1NLAD8-infected PBMCs and Tax2-, Tax1-, ebe-treated, and untreated samples at each concentration at each day of sampling. All ANOVAs with significant main effects (p<0.05) were followed by Bonferroni posttests to determine between which variables the difference existed. Data were analyzed in GraphPad Prism version 6.00 (GraphPad Software, San Diego, CA).
Results
Effect of Tax1 and Tax2 in the inhibition of HIV-1 viral replication in PBMCs
PHA-activated PBMCs from three HIV-1/HTLV-negative donors were infected with 0.8 pg/ml HIV-1NLAD8 virus stock; the cells were washed, plated for triplicate conditions, and immediately treated with a concentration range (1–100 pM) of Tax2, Tax1, or ebe as a mock control. The presence of HIV-1 could be determined in the control R5-infected PBMC group by day 7 when cell-free supernatants were measured for p24 viral antigen by ELISA. Interestingly, treatment with Tax2 protein resulted in a significant reduction (p<0.05) in HIV-1 replication from 10 to 18 days after the initial HIV-1NLAD8 infection. Tax1 at 1 and 100 pM also inhibited HIV replication at days 10 and 14 (p<0.05) (Fig. 1A–C). No significant difference in HIV-1 p24 levels was determined with the background ebe compared to R5-infected controls. Although Tax2 and Tax1 inhibited HIV-1 replication, neither treatment resulted in a total suppression of HIV-1 expression (Fig. 1).
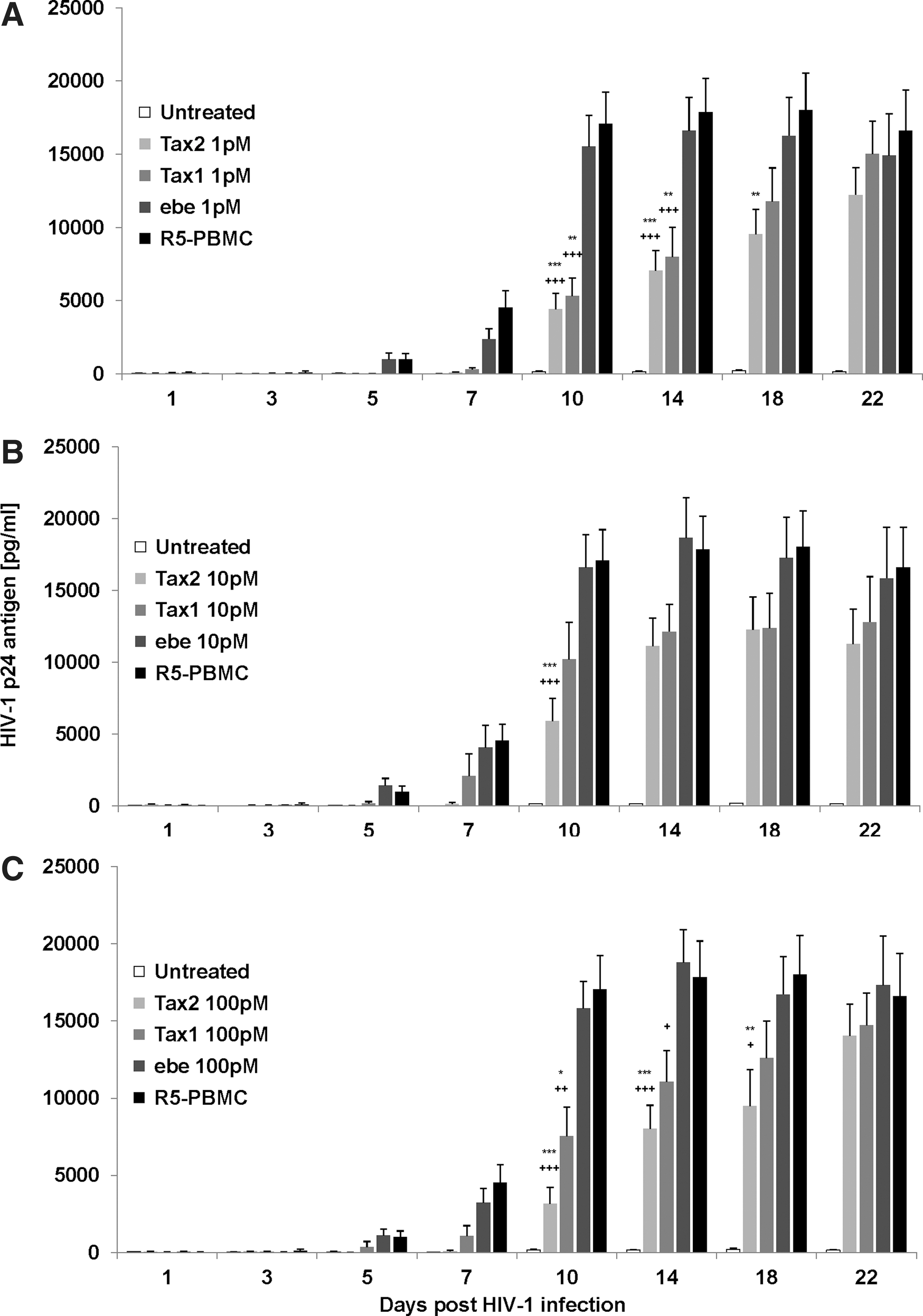
Suppression of M-tropic HIV-1NLAD8 replication in activated infected peripheral blood mononuclear cells (PBMCs) when treated with Tax2 or Tax1 proteins at different concentrations:
Expression of CC-chemokines in culture supernatant fluids
Supernatant fluids were subsequently assayed for levels of CC-chemokines. Addition of PHA resulted in spontaneous generation of CC-chemokines in both Tax-treated and mock-treated controls. Nevertheless, the daily addition of Tax proteins (Tax1 and Tax2) in groups R5-PBMC-Tax2 and R5-PBMC-Tax1 induced statistically significant higher levels of CC-chemokines MIP-1α, MIP-1β, and RANTES compared to R5-PBMC and R5-PBMC-ebe controls at days 1 through 7 (Fig. 2A–C). At days 10 to 22 after R5 infection, PBMCs treated daily with Tax proteins do not show any significant production of CC-chemokines compared to both the P5-PBMC and R5-PBMC-ebe control groups.
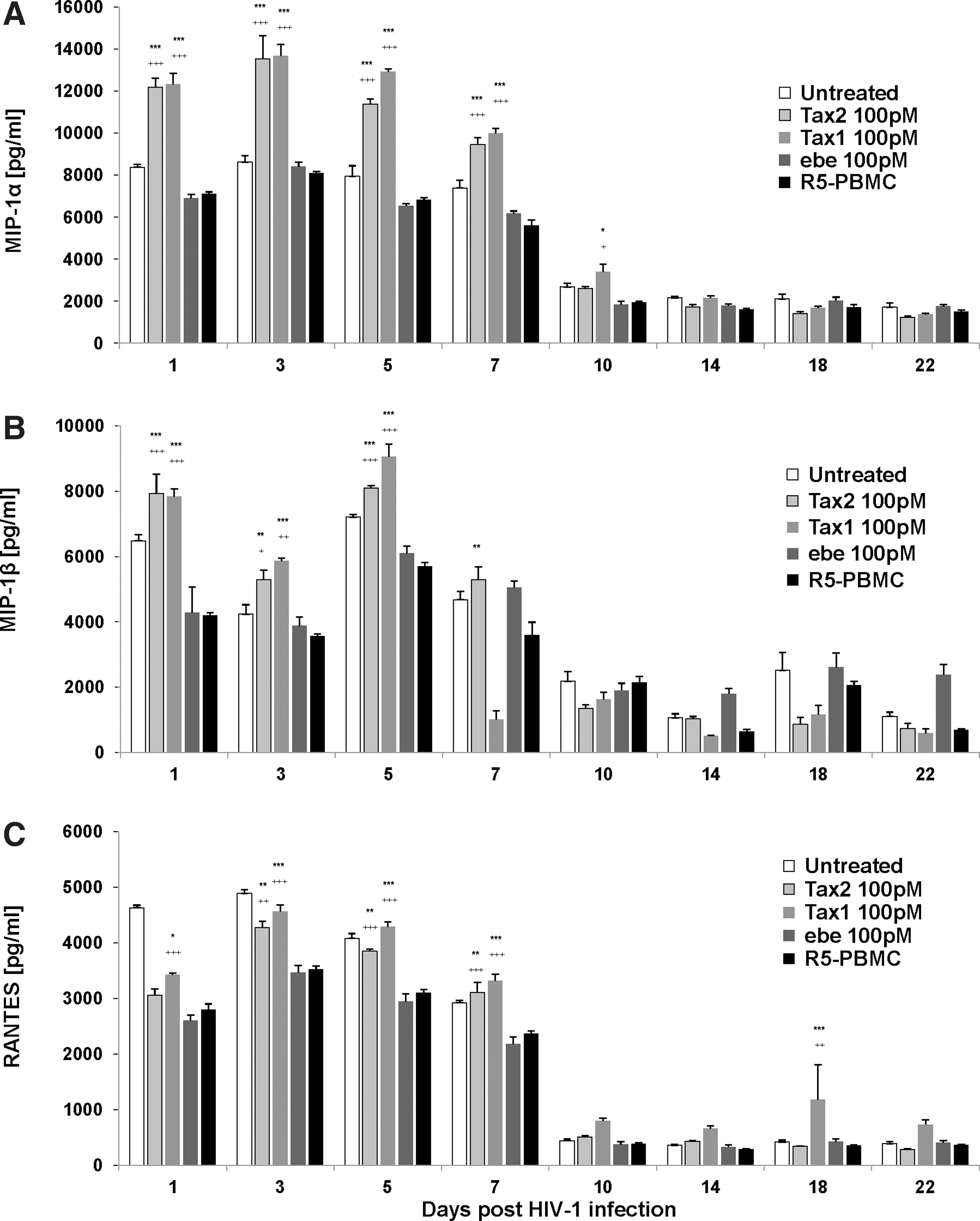
Higher levels of CC-chemokines:
Testing the ability of Tax2 protein to suppress HIV-1 viral replication in PBMCs when protein was added prior to, during, and after infection with HIV-1
To evaluate the potential HIV-suppressive effect of Tax2 at different times before or after HIV-1NLAD8/R5 infection, different groups were set up as follows: (A) Tax2 was added to PBMCs 48 h before infection; (B) Tax2 was added at once after viral infection; and (C) Tax2 was added to the PBMCs 48 h post-R5 infection. Figure 3 shows the level of HIV-1 p24 antigen in untreated or Tax2-treated PBMC culture supernatant assayed by ELISA at different times postinfection. The initial addition of 1 pM, 10 pM, or 100 pM of Tax2 starting 48 h previous to infection resulted in a significant inhibition of HIV-1 p24 levels (p<0.05) at days 7 to 14 compared to the untreated, R5-infected PBMCs (Fig. 3A). When the addition of Tax2 protein started immediately after HIV-1NLAD8 infection, significant inhibition of HIV-1 p24 levels could still be seen (p<0.05), starting at day 7 and continuing up to day 18 with all concentrations of Tax2 tested (Fig. 3B). In contrast, when the initial addition of Tax2 was performed 2 days after R5 infection, significant HIV-1 p24 levels were determined only at days 7 and 14 for 1 pM and at day 7 for 10 pM (p<0.05) (Fig. 3C).
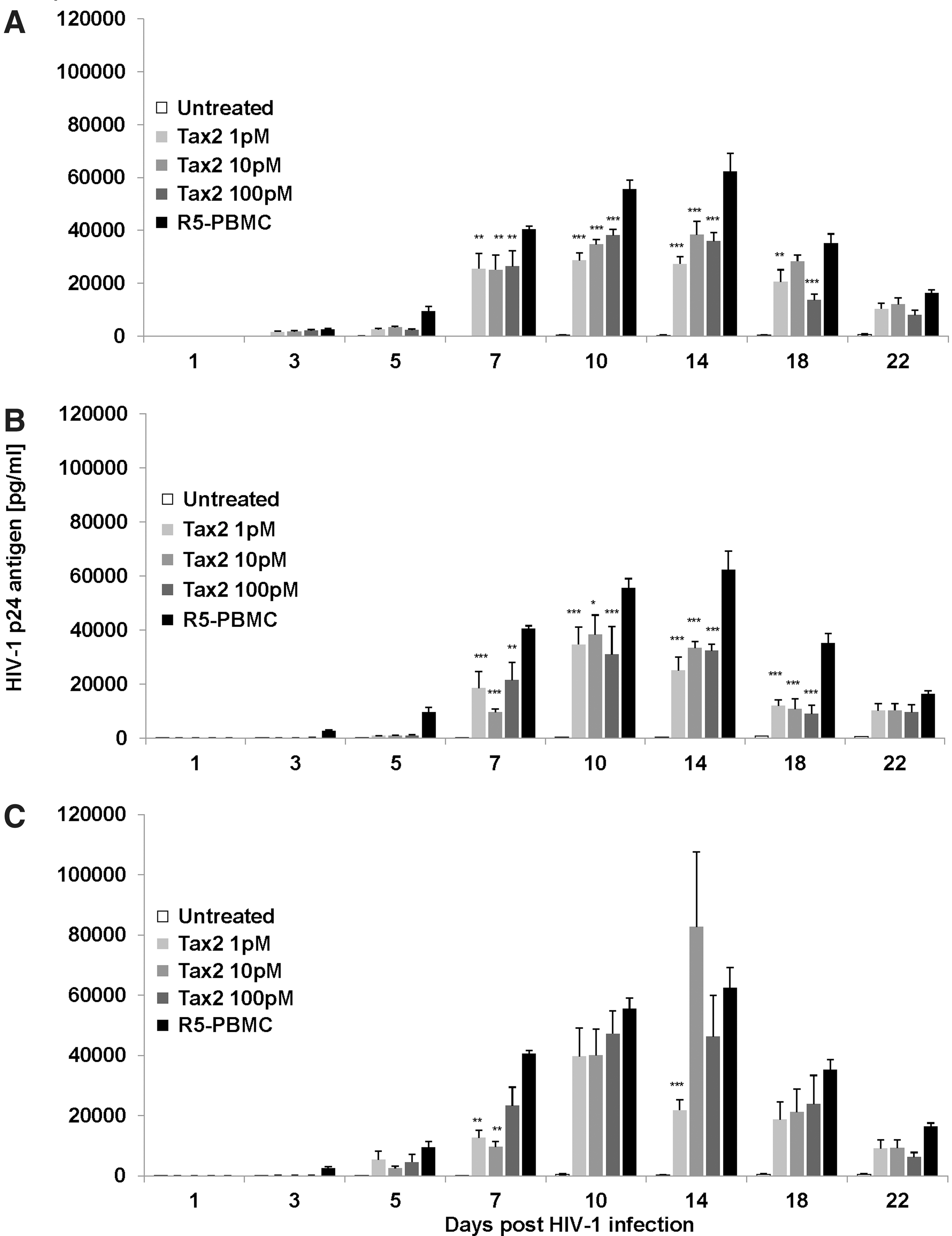
Tax2 protein inhibits M-tropic HIV-1NLAD8 replication in activated infected PBMCs when Tax2 started to be added before and immediately after R5 infection.
Discussion
In these experiments it was shown that recombinant HTLV-1 and HTLV-2 Tax proteins were able to suppress HIV-1 viral replication in PBMCs. Tax2 was more potent in this ability than Tax1, and the inhibitory effects of Tax2 were most pronounced when Tax2 was added prior to infection with HIV-1. Although the inhibition of HIV-1 p24 replication was not complete, these findings provide an indication that Tax proteins restrain the production of HIV-1 virus, possibly through induction of innate antiviral immune responses. Tax1 has been clearly implicated as an oncoprotein that targets multiple host cell transcriptional pathways. While Tax2 has a marked inability to induce leukemogenesis, it can potentially function as an immunomodulatory protein during HTLV-2 infection. The differential functional characteristics of Tax1 and Tax2 most likely explain the differences in disease conditions associated with HTLV-1 and HTLV-2. 8 In clinical scenarios where overexpression of Tax1 and Tax2 occurs, increased transactivation of cellular genes might result in immune dysregulation in affected individuals. This has been clearly shown to be the case in HTLV-1-infected individuals with HTLV-1-associated myelopathy, in whom very high levels of HTLV-1 tax/rex mRNA can be detected. 13
HIV-1 and HTLV-2 coinfections have been linked in some cases to a “long-term nonprogressor” phenotype. 7 Recently we have shown that Tax2 has the potential to modify innate host immune responses through increasing lymphocyte viability, activating monocytes and PBMC production of CC-chemokines (CCL3, CCL4, and CCL5), and downregulating the CCR5 receptor. 10,11 It is plausible that Tax2 could modify innate host responses during HIV-1/HTLV-2 coinfection, potentially by hampering the fusion and entry of the most commonly transmitted HIV-1 strains. In fact, in a study investigating whether HTLV-1 infection could modulate HIV-1 infection of surrounding cells, primary CD4+ T cells were treated with cell-free supernatants from HTLV-1-infected MT-2 cell cultures. The primary CD4+ T cells became resistant to macrophage-tropic HIV-1 and the CC-chemokines with antiviral function (CCL3, CCL4, and CCL5) were identified as the major suppressive factors. 14
HIV-1 proviral DNA integration has been reported to be detectable by 24 h after HIV-1 infection and reaches a plateau by 48 h. 15,16 Although the results herein may suggest that Tax2 inhibits virus replication mostly by inhibiting M-tropic virus entry in the early stage of virus replication, Tax2 (1 pM) was able to restrain virus replication (at days 7 and 14) when initially added 48 h after HIV-1NLAD8/R5 infection (Fig. 2C). Tax2 may also exert inhibitory effects during advanced levels of HIV-1 virus replication. Others have reported that HIV-1/HTLV-2 coinfection resulted in low/absent STAT activation in vivo that paralleled a diminished secretion of interferon (IFN)-γ after ex vivo cultivation, highlighting an interference exerted by HTLV-2 on HIV-1-induced STAT activation and supporting the hypothesis that HTLV-2 may also interfere with HIV-1 infection at multiple levels. 17
There are some limitations to this report. We were surprised that we did not observe a dose dependence attributable to Tax2-mediated inhibition of viral replication. In our earlier report 10 we observed similar levels of CC-chemokine expression at all doses of Tax2 employed (1–100 pM); this suggested that the cell signaling pathway resulting in Tax2-mediated chemokine induction is probably saturated. This could reasonably explain the lack of dose dependence observed in the current experiments. Second, only one HIV-1 viral strain was tested, an R5 tropic strain of HIV-1 (HIV-1NLAD8/R5). We elected to study our hypothesis using this particular strain as a proof-of-concept, as it seemed most likely that CCR5 receptor downregulation would be the possible mechanism by which HIV/HTLV-2 dual infections would result in a slow-progressor phenotype. Other laboratory strains of strictly R5 tropic virus should ultimately be tested as well to confirm this hypothesis. Additionally, it would be interesting to know if Tax2 could modulate replication of strains of HIV-1 that infect PBMCs independent of CCR5, such as syncytia-forming CXCR4-tropic strains of HIV-1. 18 We previously tested the ability of Tax2 to affect replication of an X4 tropic strain of HIV-1 (HTLV-IIIb), and were unable to see any significant effects of Tax2 on levels of HIV-p24 expression in X4-infected PBMC cultures (data not shown). Nonetheless, we think it is possible that Tax2 could modulate replication of X4 tropic strains of HIV-1 in PBMCs in vivo through an indirect mechanism, possibly through activation of natural killer cells by CC-chemokines. Further work is underway to explore this possibility.
An additional concern is that the PHA used in these experiments has pleotropic effects on PBMCs. For example, PHA can induce expression of CD4+ receptors on CD8+ T cells, which has been suggested as a potential mechanism for HIV-1 infection of CD8+ T cells in advanced HIV/AIDS. 19 Additionally, we have previously shown that Tax2 (and to a lesser degree Tax1) promoted increased viability in fresh, unstimulated PBMCs. 10 If anything, this would provide an increased population of susceptible T cells in which HIV could replicate. With that said, we used two different controls (non-Tax treated, and mock treated, using bacterial lysate without the expression vector) to control for the pleiotropic effects of PHA in all experiments. In each donor tested, we found similar inhibitory effects of Tax2 when compared to controls. Additionally, in all conditions, PHA treatment resulted in nonspecific release of CC-chemokines (Fig. 2), although levels of the chemokines were significantly higher in Tax-treated specimens.
Interestingly, our findings follow a paradigm observed with other viral copathogens infecting persons with HIV-1. Studies showed that coinfection with GBV-C slows the progression of HIV-1 disease and coinfected individuals can survive longer than those without GBV-C. 20 African variants of GBV-C have been shown to replicate in PBMCs and to inhibit the replication of HIV in vitro via the mediation of induced CC-chemokines and the downregulation of CCR5. 21,22 Another group has reported in vivo and in vitro evidence of HIV-1 replication inhibition by measles virus (MV) coinfection concurrent with intense immune activation including higher plasma levels of RANTES in HIV-infected children with measles. 23,24
In conclusion, the HTLV Tax2 protein appears to downregulate HIV-1 viral replication in vitro, and implicates Tax2 as a potential immunomodulatory agent capable of influencing clinical outcomes in HIV-1-infected individuals.
Footnotes
Acknowledgments
This work was supported by funds received from VA Merit Review Grant BX000488-01 and the Department of Medicine of the Medical College of Wisconsin.
Author Disclosure Statement
No competing financial interests exist.