
Abstract
We have designed a therapeutic HIV-1 vaccine concept based on peptides together with the adjuvant CAF01. Peptides represented 15 HLA-supertype-restricted subdominant and conserved CD8 T cell epitopes and three CD4 T-helper cell epitopes. In this phase I clinical trial, safety and immunogenicity were assessed in untreated HIV-1-infected individuals in Guinea-Bissau, West Africa. Twenty-three HIV-1-infected individuals were randomized to receive placebo (n=5) or vaccine (n=18). Safety was appraised by clinical follow-up combined with monitoring of biochemistry, hematology, CD4 T cell counts, and HIV-1 viral loads. T cell immunogenicity was monitored longitudinally by interferon (IFN)-γ ELISpot. New vaccine-specific T cell responses were induced in 6/14 vaccinees for whom ELISpot data were valid. CD4 T cell counts and viral loads were stable. The study shows that therapeutic immunization is feasible and safe in Guinea-Bissau and that it is possible to redirect T cell immunity with CAF01-adjuvanted HIV-1 peptide vaccine during untreated HIV-1 infection in some patients. However, relatively few preexisting and vaccine-induced HIV-1 T cell responses to CD8 T cell epitopes were detected against HIV-1 using IFN-γ ELISpot in this chronically infected African population.
Introduction
T
Therapeutic HIV-1 peptide immunization strategies aim to stimulate T cell responses to new HIV-1 epitopes not normally targeted during infection with the aim of reducing HIV-1 viral load among infected individuals and sustaining their immunological competence. 6,7 Based on evidence arguing for a protective role of subdominant T cell responses against HIV-1 and other viral infections, 8 –11 our group has focused on developing a novel therapeutic HIV-1 vaccine approach exploiting the duality of conserved and infrequently targeted, or subdominant, CD8 T cell epitopes. 12 –18 To cover the high diversity of HIV-1 and human tissue types we designed an experimental vaccine based on subdominant epitopes conserved among HIV-1, irrespective of clade or geographic origin, restricted by common HLA supertypes. 17 HLA class I molecules can be clustered into groups, designated as supertypes, representing sets of molecules that share largely overlapping peptide binding specificities. 19,20 We have previously shown that immunization with autologous monocyte-derived dendritic cells (MDDCs) pulsed with peptides representing seven HLA-A2-restricted, subdominant HIV-1 cytotoxic T lymphocyte (CTL) minimal epitopes and three CD4 T helper epitopes elicited previously undetectable T cell responses in vivo in 12/12 ART-naive HIV-1+ and HLA-A2+ vaccinees. 12,15
In an effort to broaden this principle to a wider range of HLA repertoires and HIV-1 strains, also relevant for Africa, we have added eight additional non-HLA-A2-restricted HIV-1 CTL peptides to the subdominant T cell vaccine concept bringing it to a total of 15 HIV-1-CTL peptides. These T cell epitopes can theoretically bind to the five predominant HLA supertypes worldwide and are conserved among HIV-1 strains in Denmark and Guinea-Bissau.
13,16,17
Three CD4 T cell epitope peptides were codelivered to provide help for CD8 T cell immune induction.
21
In addition, to provide a delivery vehicle feasible for large-scale clinical use, which may enhance the immunogenicity of these 18 epitopes, the peptides were formulated in the new Cationic Adjuvant Formulation (CAF01).
22,23
The CAF01 adjuvant, a synthetic liposomic adjuvant composed of the polar glycolipid TBD [
Materials and Methods
Study design and ethics
Study approval was obtained from the National Committee on Health Ethics, Ministry of Health in Guinea-Bissau (Comité Nacional da Ética na Saúde No. Ref 148/CNES/2010). A consultative statement of approval (J.nr.: 2009-37919) was also obtained by the Developing-Country Committee of the Danish National Committee on Biomedical Research Ethics. Informed consent was obtained from all individuals. HIV-1-seropositive and HIV-2-seronegative adults were recruited from a cohort of individuals being followed at the Centro de Tratamento Ambulatório, Hospital Nacional Simão Mendes, Guinea-Bissau. The inclusion criteria were HIV-1+, HIV-2−; CD4 T cell counts >400 cells/μl; not on ART; male or female between 18 and 50 years of age; normal blood biochemistry and hematology values; and likelihood to complete follow-up and written informed consent after an explanation of the trial was presented orally and in writing. The exclusion criteria were pregnancy, active chronic infectious diseases likely to influence HIV-1 infection, e.g., HIV-2 infection, active tuberculosis or coinfection with hepatitis B or C viruses; clinically significant blood biochemistry or hematology parameters outside the range of normal values; simultaneous treatment with other experimental drugs; active autoimmune diseases; history of severe allergy or anaphylactic shock; significant medical disease as judged by the investigators; and vaccinated with other vaccines within 3 months before the first therapeutic immunization or treatment with immune-modulating medicine within 3 months before the first therapeutic immunization. A total of 20 females and three males from six major ethnic groups inhabiting Guinea-Bissau were enrolled (Table 1). Baseline CD4 T cell counts, biochemistry, and hematology values for these 23 individuals have been described previously. 27 These 23 subjects were enrolled and randomly assigned to one of the two treatment arms: 18 received the peptide-CAF01 vaccine and five received placebo. Fifteen vaccinees and three placebo recipients completed the whole study including the 6-month follow-up.
Cells/μl blood.
Log10 (RNA copies/ml plasma).
The indicated CD4 T cell counts and viral loads are those observed in samples taken during enrollment on day 0, the day of the first immunization.
Vaccine and immunization
The 15 HIV-1 subdominant CD8 T cell peptides plus two HIV-1 CD4 T helper peptides and one non-HIV, universal CD4 T helper PADRE peptide 28 contained in the vaccine have been described in detail previously with regard to their selection rationale, HLA binding, immunogenicity and antigenicity during HIV-1 infection, GMP production, and preclinical toxicity. 12,15,17,25,29 The peptides are listed in Table 2.
The freeze-dried peptides and CAF01 liquid adjuvant GMP batches were transported via temperature-monitored cold-chains at below −15°C and 2–8°C, respectively, and stored accordingly. The vaccine was reconstituted by the addition of 2.0 ml CAF01 solution to 7.2 mg of the peptide mix (containing 400 μg/peptide). Then 1.25 ml of the peptide/CAF01 vaccine was injected intramuscularly on weeks 0, 2, 4, and 8. Saline (1.25 ml) was used as placebo since it is considered unethical to use adjuvant alone, given that it has no known benefit but may generate adverse reactions. The predetermined randomization list was blinded for the participant (single blinded) and for the scientists analyzing the data.
Safety and reactogenicity evaluation
After each immunization, participants were observed for 30–60 min to monitor any immediate adverse events (AEs). Participants were contacted by mobile phone 24 h postimmunization to ascertain vaccine tolerability. Solicited local and general AEs were recorded at six visits (on weeks 2, 4, 8, 10–15, 20–24, and 32–36).
Blood samples were drawn at each of the seven visits (on weeks 0, 2, 4, 8, 10–15, 20–24, and 32–36) to monitor changes in CD4 T cell counts, HIV-1-viral load, and blood biochemistry and hematology at each of six visits (on week 0, 2, 4, 8, 10–15, and 32–36). CD4 T cell counts were determined by standard flow cytometry using a CyFlow SL Blue (Partec GmbH, Görlitz, Germany). Viral load in plasma was quantified by the COBAS Ampliprep/COBAS TaqManHIV-1 Test, version 2.0 system (Roche Diagnostics, Copenhagen, Denmark), which has been shown to accurately quantify various HIV-1 subtypes, including the predominant CRF02_AG, A3, and C circulating in Guinea-Bissau. 30 These analyses were performed in Denmark. For biochemistry analyses, we tested sera using a Reflotron Plus Clinical Chemistry Analyzer with Roche rapid tests for creatinine, bilirubin, and aspartate aminotransferase (AST/GOT), according to the manufacturer's instructions. For hematology analyses, whole blood was collected in Vacuette, Greiner bio-one, K3EDTA tubes and the blood was analyzed using a Medonic CA530 Oden Hematology Analyzer (Clinical Diagnostic Solutions, Plantation, FL) and Medonic reagent sets according to the manufacturer's instructions. Twelve biochemistry and hematology parameters were measured using methods and ranges reported previously in detail. 27 Due to temporary problems at the hematology laboratory, we were not able to measure hemoglobin, leukocytes, differential counts, and platelets between October 6 and December 10, 2010, which was the time period during which most participants came for their third and/or fourth vaccination. Instead, the participants were evaluated clinically for any signs of anemia or thrombocytopenia.
HLA genotyping
Genomic DNA was purified using the QIAamp DNA Mini Kit (Qiagen) and HLA genotyped at Genome Diagnostics BV.
Immunogenicity evaluation
To test the reactivity of T cells against each of the 18 peptides contained in the vaccine, we used the interferon (IFN)-γ ELISpot assay at four time points, weeks 0, 10–15, 20–24, and 32–36. Blood samples were collected in BD Vacutainer cell preparation tubes (CPT) containing sodium citrate buffer (Becton Dickinson, Franklin Lakes, NJ). Peripheral blood mononuclear cells (PBMCs) were isolated and used immediately. The alkaline phosphatase-conjugated version of the Human IFN-γ ELISpotPRO kit (Mabtech, Stockholm, Sweden) was used according to the manufacturer's instructions. PBMCs (1.25×105) were added per well and the final concentration of each individually tested peptide was 8 μg/ml. Phytohemagglutinin A (PHA) (0.21 μg/ml) (Remel c/o Thermo Fisher Scientific, Roskilde, Denmark) and 0.2% dimethyl sulfoxide (DMSO, Sigma-Aldrich, St. Louis, MO) were used as positive and negative controls, respectively. Plates were shipped to Denmark and spots were counted at the Statens Serum Institut using an AID ELISpot reader (Autoimmun Diagnostika, Strassberg, Germany). ELISpot positivity was defined as >4 times the background and significantly different from that of control wells, according to the DFR(eq) statistical test. 31,32
Assessment of vaccine feasibility
The feasibility of conducting vaccine studies in HIV-1-positive individuals in Guinea-Bissau was appraised by counting the number of participants completing an 8-month clinical trial period. Participants had an additional consultation and physical examination 3 months after the clinical trial period ended and received a detailed description of the laboratory procedures and an explanation of the trial results and were able to ask any questions about the explained procedures. All interviews were conducted in Kiriol, the most commonly spoken language in Guinea-Bissau.
Results
Safety
No signs of allergic reactions were observed in any of the participants. When the participants were interviewed over the phone 24 h after vaccination, none complained of any general swelling, aching, itching, or visible signs of irritation at the injection site. This was also corroborated clinically within 2 weeks after each immunization. One vaccinee experienced tiredness and headache grade 1 after the fourth vaccination that resolved within 3 days. No other AEs were reported. Two male subjects receiving placebo (saline) did not complete the study: one 34-year-old male (P12) was discontinued after his second immunization because of elevated liver enzymes, i.e., high bilirubin and aspartate aminotransferase (GOT/AST) at grade 4, according to the AIDS Toxicity Grading Scales. The reason for the elevated liver enzymes was not identified. The other (P19) withdrew after the fourth vaccination. Three female study subjects receiving vaccine did not complete the study: the first (P6) was lost to follow-up after the first immunization due to insufficient address information, the second (P22) stopped before the fourth vaccination visit due to an unwillingness to provide blood samples, and the third (P11) represented a serious AE deemed to be unrelated to the vaccine. This participant was a 28-year-old woman diagnosed with HIV-1 in August 2010. She received the first three vaccinations, without experiencing any side effects. She had stable CD4 T cell counts (>400/μl) and viral load. Hematology and biochemistry parameters were within normal ranges, apart from a slightly low hemoglobin (Hb) value of 9.1 g/dl, which was not considered to be clinically important, as African females are known to have lower Hb values. 27,33,34 At her second visit, she was on treatment with sulfamethoxazole/trimethoprim and vitamin B for stomatitis. When she came for her last vaccination, she had had diarrhea, fever, cough, and oral candidiasis for the past 2 weeks. The symptoms were only moderate and were considered unrelated to the vaccine and it was decided to postpone the vaccination. The patient was referred for chest x-ray and sputum examination for tuberculosis, which she did not attend, and treatment with metronidazole, albendazole, and loperamide was started. A new appointment was scheduled; however, the participant decided to see the local doctor in her village instead and died on November 21, 2010 due to severe gastroenteritis and pneumonia (clinical judgment). The death was deemed unrelated to the vaccine. Eighteen participants completed all four injections and the 6-month follow-up.
HIV-1 viral load and CD4 T cell counts
We monitored the baseline and change in viral load during immunization and up to 6 months of follow-up (Fig. 1, Table 1). Among the 18 participants completing the 6-month follow-up no significant changes in viral load were seen. The minor fluctuations observed in viral load during the immunization regimen were <0.5 log. Overall, CD4 T cell counts in both groups were stable (Fig. 1). Thus, there was no observable detrimental effect of the vaccine.
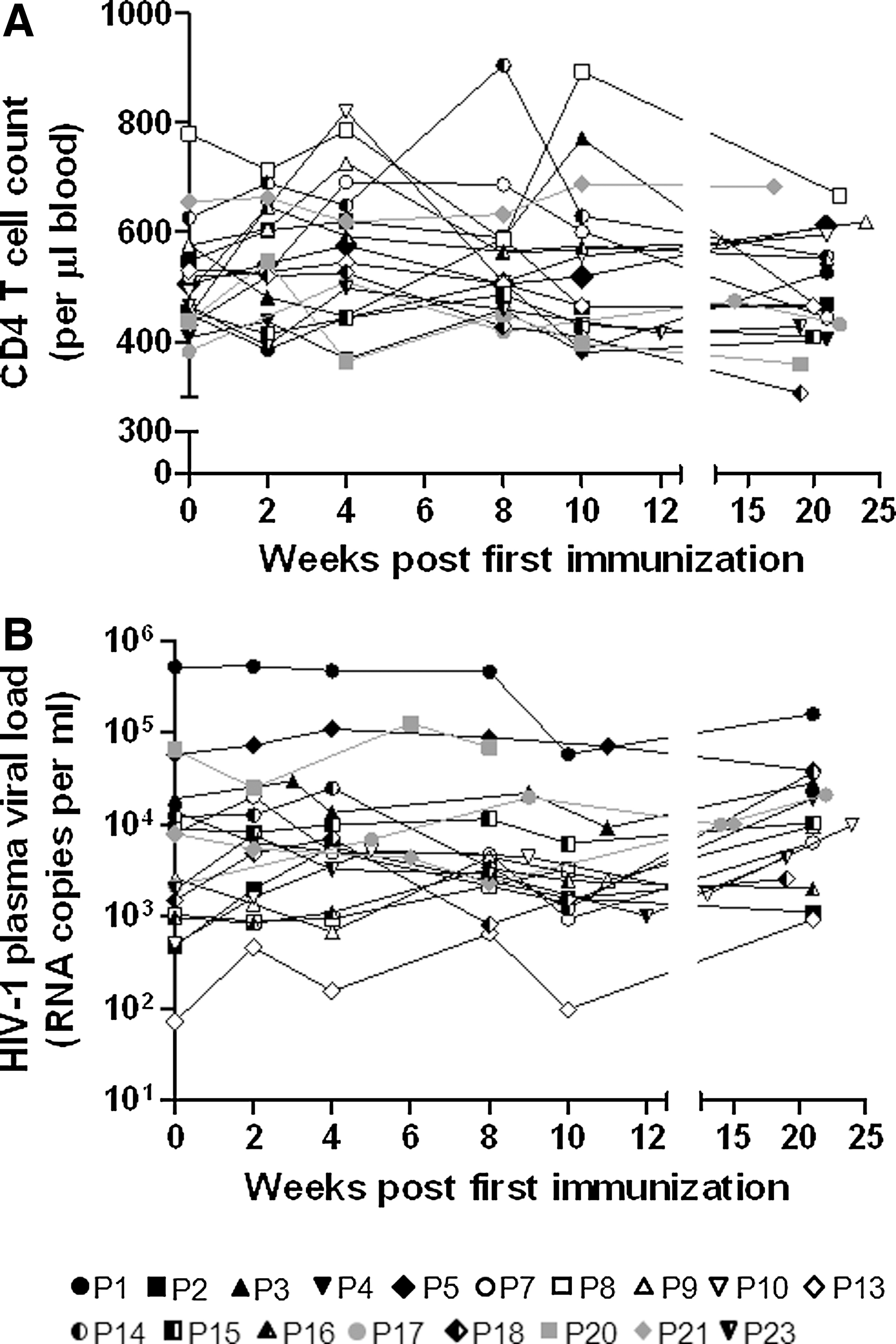
Longitudinal changes in
Immunogenicity
As expected, baseline IFN-γ ELISpot responses to the HIV-1 epitope peptides were sporadic (Fig. 2). The low level of baseline T cell reactivity to the peptides was not due to the limitations of the level of detection in the IFN-γ ELISpot assay, as PBMCs from all individuals were shown to react potently with the positive control (PHA) as well as positively with a reference pool of clade A HIV-1 Gag peptides, 32 thereby confirming that the peptides indeed contain subdominant and infrequently targeted T cell epitopes. 17,29 The baseline IFN-γ ELISpot responses ranged from 42 to 2,648 (median 312) SFU per million PBMCs. We were unable to detect any boosting of these responses by vaccination. Of the 10 preexisting T cell responses observed to individual peptides among 14 evaluated vaccinees, seven were conserved throughout the immunization regimen. HIV-1 clade-A-specific T cell responses (positive control) were also preserved. One individual in the vaccine group and one individual in the placebo group had consistently high background ELISpot responses when their PBMCs from four separate bleeds were tested; as such, no immunogenicity data could be gathered from them (Fig. 2). All HIV-infected individuals had HLA alleles belonging to at least one of the five predominant HLA supertypes (HLA-A1, -A2, -A3, -B7, -B44) predominating in Guinea-Bissau and Denmark 16 and chosen for the vaccine design 17 (Fig. 2). The maximal theoretical CD8 T cell epitope reactivity matching the HLA supertypes of these participants ranged from 1 to 12 vaccine CD8 T cell epitopes per individual. Eight of the 13 T cell responses to CD8 T cell epitope peptides (preexisting and induced) matched the HLA type of the individual, whereas five responses (four preexisting and one vaccine induced) were directed to epitopes outside the predicted HLA supertype restriction (Fig. 2).
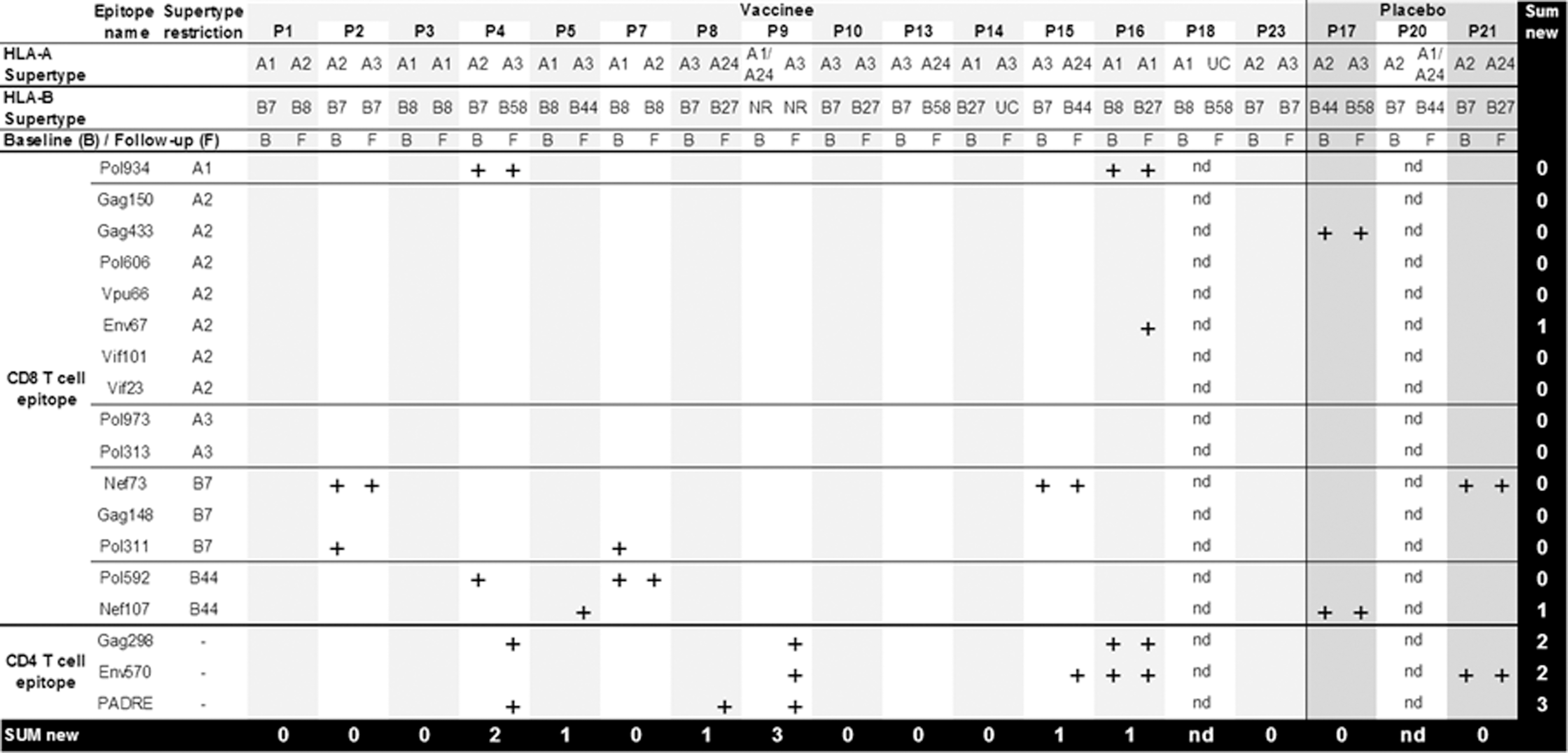
Interferon (IFN)-γ ELISpot responses to individual HIV-1 peptides. Plus signs (+) denote a cumulative response (detected at any time point) measured by IFN-γ ELISpot. Plus signs denote positive responses observed before immunization (B, baseline) and along the course of immunization (F, follow-up). nd, not determined because of high background. UC, unclassified. NR, not resolved.
Induction of nine new T cell responses was observed among vaccinees, compared to zero new responses in individuals receiving placebo. Cumulative responses (i.e., detected at any time point) are shown in Fig. 2. Out of these nine new responses, six were detected at the first evaluation (2 weeks after the last immunization), two appeared at the second evaluation (3 months after the last immunization), and one appeared at the third evaluation (6 months after the last immunization). Seven of the induced T cell responses were directed to CD4 T cell epitopes and two responses were directed to CD8 T cell epitopes. Overall, the vaccine induced new T cell responses in six of the 14 vaccinees from whom ELISpot data were valid and interpretable (Fig. 2). The vaccine-induced responses detected by IFN-γ ELISpot ranged from 36 to 1,140 (median 128) SFU per million PBMCs. Most of the vaccine-induced responses were transient, i.e., they dropped below the detection level for the assay by the following evaluation time point. Only two new responses were detected at two time points and one response was sustained over all three time points (Fig. 3). Among the six IFN-γ ELISpot responders in this cohort, the potency of the HIV-1 T cell responses was greatest and most consistent in vaccinee P15 (Fig. 3).
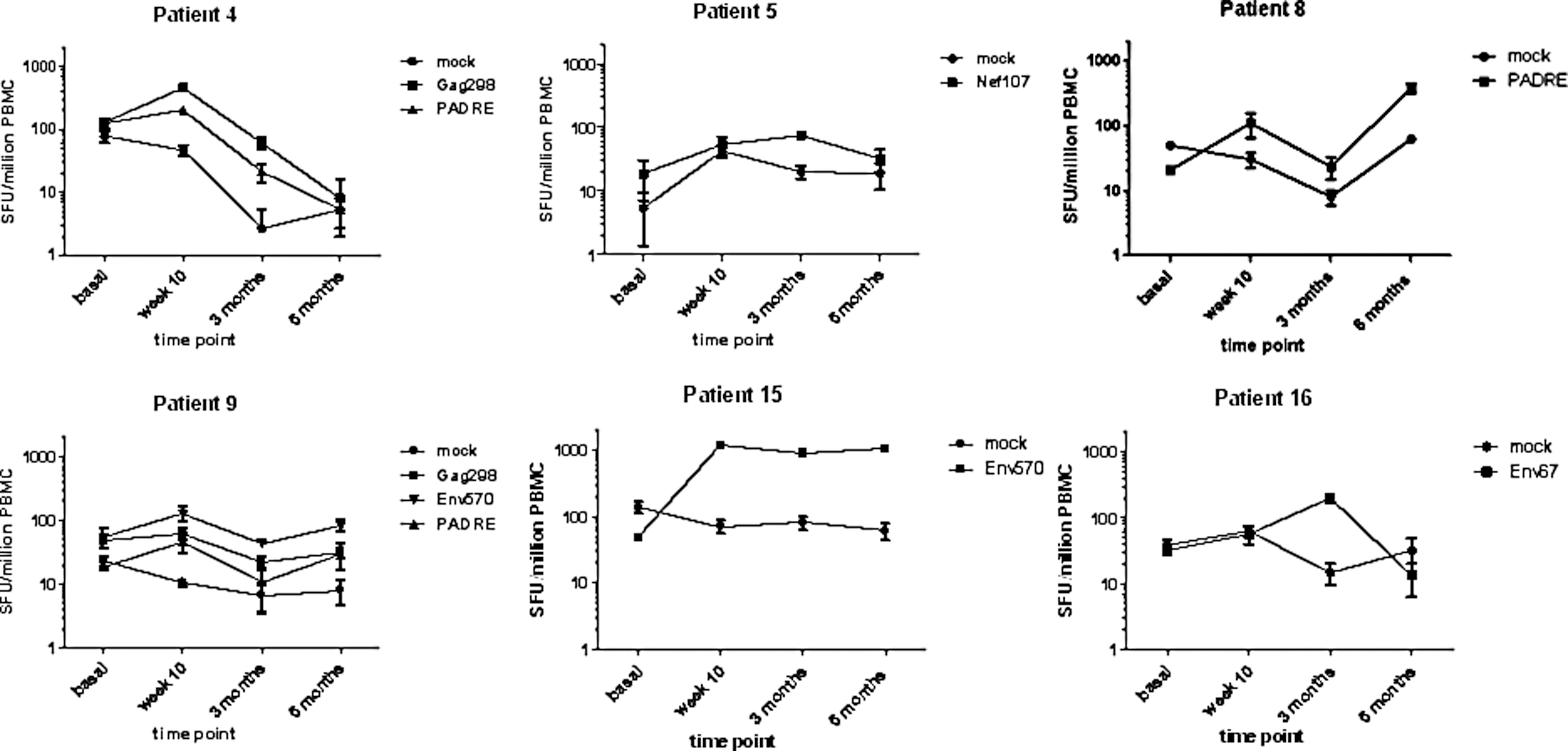
Longitudinal IFN-γ ELISpot responses in six vaccine responders. Spot-forming unit (SFU) counts for each individual in response to the indicated stimuli are presented per million peripheral blood mononuclear cells (PBMCs) at four time points along the therapeutic immunization regimen. Mock denotes background responses to 0.2% dimethyl sulfoxide (DMSO). Each point represents the mean of three responses observed for each donor. Error bars denote the standard error of the mean.
Discussion
In a cohort of treatment-naive HIV-1-infected West African individuals, we report on the feasibility, safety, and immunogenicity of a therapeutic immunization with minimal T cell epitope peptides in CAF01. We used a therapeutic HIV-1 vaccination concept attempting to redirect T cell immunity to infrequently targeted, subdominant epitopes conserved among HIV-1, irrespective of clade or geographic origin. Moreover, the epitopes were restricted by five common HLA supertypes in order to overcome differences in HLA distributions in different geographic regions.
The vaccine has previously been tested for safety in a Danish population of asymptomatic, treatment-naive, HIV-1-infected male individuals 18 before being tested in Guinea-Bissau, West Africa. The aim was to evaluate the universal design, i.e., to test the immunogenicity in a distant geographic region, where the HLA distributions 17 as well as the circulating HIV-1 subtypes are quite different. 35 The vaccine was safe and well tolerated also in this cohort of treatment-naive HIV-1-infected individuals, predominantly women, living in Guinea-Bissau. It was noteworthy that only one solicited AE was observed among the vaccinees in Bissau as compared to, e.g., soreness at the injection site in 25 out of the total 44 (4 times 11) injections in the 11 Danish study participants. 18 It is conceivable that differences in culture and/or gender between male participants in Denmark and female participants in Guinea-Bissau could account for this difference in perception and reporting of solicited AEs. Because CD4 T cells were sustained over 7–9 months in the 15 HIV-1-infected vaccinees, the data presented here, together with the Danish study, 18 provide the first observation that immunization with HIV-1 peptides and CAF01 is safe in an asymptomatic HIV-1-infected population with high CD4 T cell counts.
The vaccine induced new T cell vaccine responses in 6/14 vaccinees in Guinea-Bissau as evaluated by IFN-γ ELISpot. These data differ from those reported in the prior study conducted in Denmark, where 10/10 vaccinees developed new T cell responses to 12 of the 15 CD8 T cell epitopes and to all three CD4 T cell epitopes. 18 However, immunogenicity in the latter study was evaluated using the IFN-γ ELISpot test plus intracellular cytokine staining (ICS) followed by flow cytometry for IFN-γ, interleukin (IL)-2, tumor necrosis factor (TNF)-α, macrophage inflammatory protein (MIP)-1β, and CD107a, which has the potential to detect a broader range of responses compared to the IFN-γ ELISpot used in this African study. In fact, in the Danish study the vaccine-induced CD8 T cell responses were dominated by the expression of one or two functional markers, particularly MIP-1β and/or CD107a. Comparing only the IFN-γ ELISpot data, 7 of 10 Danish vaccinees showed new vaccine responses in the ELISpot assays 18 versus 6 of 14 in Guinea-Bissau (not statistically significantly different, Fisher's exact test). Thus, the peptide vaccine was immunogenic also in Guinea-Bissau albeit somewhat lower. A limited set of IFN-γ ELISpot and ICS experiments measuring IFN-γ, IL-2, and TNF-α was conducted in parallel at one time point during the second evaluation (week 20–24) in Guinea-Bissau. Six additional T cell responses were seen by ICS; however, as we did not have baseline ICS data in Guinea-Bissau we cannot determine whether these responses were vaccine induced or preexisting (data not shown). Looking at only IFN-γ ELISpot reactions directed to the CD8 T cell epitope peptides, the number of responders (2 of 14) in Guinea-Bissau was, however, significantly fewer than in Denmark (7 of 10) (p=0.010, Fisher's exact test). Three of the vaccine responses appeared in a delayed fashion, i.e., detected at the second or third evaluation. We can only speculate that these delayed responses are a result of the properties of the adjuvant CAF01. DDA-based liposomes are known to have depot-forming properties, e.g., a slow, low-level release of antigen over prolonged time. 22 –24
Consistent with this, the degree of preexisting T cell immunity to HIV-1, as measured by IFN-γ ELISpot to the CD8 epitopes, was lower in the study participants (both vaccine and placebo groups) in Guinea-Bissau, in whom 11 ELISpot reactions were detected out of the pool of the 240 possible reactions tested (15 CD8 T cell epitopes times 16 participants) as compared to the 17 ELISpot reactions of the pool of the 165 possible reactions tested (15 epitopes times 11 participants) in Denmark 18 (p=0.029 Fisher's exact test). Although our data are limited, they suggest a lower degree of specific HIV-1 CD8 T cell immune reactivity among the tested population of West African HIV-1-infected females compared to the HIV-1-infected males in Denmark, although all study participants, in Denmark as well as in Guinea-Bissau, had HLA alleles belonging to at least one of the five HLA supertypes, HLA-A1, -A2, -A3, -B7, -B44, and chosen for the vaccine design. 17 The maximal theoretical CD8 T cell epitope reactivity matching the HLA supertypes of these participants ranged from 1 to 12 vaccine CD8 T cell epitopes per individual. Interestingly, we and others have found a far wider promiscuity of epitope cross-reactivity than originally suggested by the HLA supertype concept, 8,18 –20 which was also observed in this study. This HLA promiscuity suggests a potential wider use of epitope-based vaccines. Nevertheless, binding affinity for the specific alleles has not been determined and such differences may contribute to the difference in specific HIV-1 CD8 T cell immunity between the two populations. Moreover, differences in factors such as gender, disease burden or general health status, regulatory T cell population, and other inflammatory inhibitory mechanisms could influence CD8 T cell reactivity. 36 –38 Differences in HIV vaccine responses induced in vaccine recipients from African and U.S. sites have also been observed in the HVTN073 study (G. Gray, abstract X20109, the Keystone Symposium “HIV Vaccines,” Keystone, Colorado, February 20–15, 2013). This is an issue that will require further parallel studies to resolve.
The prospect of using HIV-1 peptides as immunotherapeutic agents has gained more credibility, 4,6 but a major problem has always been the intrinsically poor immunogenicity of peptide antigens when administered without adjuvant and the disparity between synthetic peptides' in vitro antigenicity and their in vivo immunogenicity. 39 We have previously been able to display good immunity when we immunized HIV-1-infected individuals with autologous MDDCs pulsed with peptides. 15 However, the use of MDDCs is not feasible in Guinea-Bissau. To address these obstacles, we have formulated HIV-1 peptides in the novel adjuvant system CAF01. 25 These are the first parallel studies to test a therapeutic HIV vaccine in Europe and Africa using the same subdominant HIV-1 epitope peptides with CAF01 as a new adjuvant delivery system for immunotherapy.
It is noteworthy that seven out of the nine new responses observed in this African study were directed toward CD4 T cell epitope peptides. In contrast, a more balanced CD4 and CD8 T cell response was seen in Denmark where eight out of the 18 new IFN-γ ELISpot T cell responses were directed to CD4 epitopes as confirmed by ICS. 18 This could be due to the ELISpot method preferentially identifying CD4 responses, a more impaired CD8 T cell function, or a relatively low CD8 T cell immunogenicity of HIV-1 and the vaccine in the African participants. One possible way to increase CD8 T cell immunogenicity 23,24 may be to reformulate the vaccine into a different adjuvant system, e.g., adding poly(I:C) to CAF01, which then preferentially stimulates CD8 T cell immunity (K. Korsholm et al., unpublished observations) or including chemokines such as granulocyte monocyte colony-stimulating factor (GM-CSF) to target antigen presentation via dendritic cells. 40 –43 Again, those approaches are dependent on immune function being preserved to a sufficient extent during chronic HIV-1 infection.
Although the study was not powered to demonstrate efficacy, the lack of an effect on viral load after vaccination was disappointing. It may be due to the low level of T cell responses generated after vaccination, which may not be sufficient to inhibit viral replication. Also in the Danish study, where more responses were induced, there were no significant effects on viral load. 18 It is possible that the patient virus strains targeted did not contain the target sequence conserved before or during the study (escape mutations). As all but one of the T cell responses were transient, improvements in the vaccine's immunogenicity may be needed to reach a greater therapeutic potential in untreated HIV-1-infected African individuals. However, immunogenicity alone (as assessed by IFN-γ production) may not be sufficient to confer protective T cell immunity against HIV-1. 44 Alternatively, the lack of observed effect may reflect the immune status of the vaccinees: the therapeutic T cell vaccine could be investigated in more immune-competent HIV-1-infected individuals, e.g., during ART. It is well known that immune function during chronic untreated HIV-1 infection is progressively disrupted, 45,46 which poses an intrinsic problem for therapeutic vaccinations. 44 Such immune dysfunction may be partly reversible 47 and ART has been shown to partially restore the impaired and skewed T cell immune function in HIV-1 infections. 47,48 This is supported by the improved antiviral effects on HIV/SIV achieved by therapeutic vaccinations during ART. 49 –52
Important lessons can be learned from this African study. First, the therapeutic immunizations were well tolerated, as evidenced by both clinical observations as well as longitudinal measurements of viral load and CD4 T cell counts. Second, while performing a clinical phase I HIV vaccine study we have built the capacity to perform clinical phase I HIV-1 vaccine studies in Guinea-Bissau. Finally, we have demonstrated that new immunity can be induced to selected HIV-1 T cell epitopes in two geographically distinct and ethnically diverse untreated HIV-1-infected populations, e.g., in Northern Europe and West Africa. It also appears that the immunogenicity of the vaccine, as well as preexisting HIV-1 CD8 T cell immunity, seemed lower in this West African HIV-1-positive untreated study group as compared to the similar vaccinated Danish European study group. 18 If confirmed, this may have implications for the design of HIV-1 and other vaccines for certain African versus European populations.
Footnotes
Acknowledgments
We would like to thank all the Bissau Guinean volunteers for participating in this study; Peter Aaby for support and guidance through the Bandim Health Project; Zacarias de Silva, Laboratorio Nacional de Saude Publica (LNSP), Bissau for expert laboratory guidance and support; Inés Oliveira Souto for guidance at the Centro de Tratamento Ambulatório, Hospital Nacional Simão Mendes, Bissau; Susanne Thelle, Lene Pedersen, Birgit Knudsen, Solvej Jensen, Irene Jensen, and Bente Østergaard for technical assistance; Gitte Kronborg for providing expert clinical advice and guidance in HIV patient management; Mark Doherty for critical reading of the manuscript; and Peter Hayes for kindly providing the Clade A Gag peptides. This work was supported by grants from the Danish International Development Agency (DANIDA) and the European & Developing Countries Clinical Trials Partnership (EDCTP) and the Danish AIDS Foundation.
Author Disclosure Statement
No competing financial interests exist.