
Abstract
Studies in persons of European descent have suggested that mitochondrial DNA (mtDNA) haplogroups influence antiretroviral therapy (ART) toxicity. We explored associations between mtDNA variants and changes in endothelial function and biomarkers among non-Hispanic white, ART-naive subjects starting ART. A5152s was a substudy of A5142, a randomized trial of initial class-sparing ART regimens that included efavirenz or lopinavir/ritonavir with nucleoside reverse transcriptase inhibitors (NRTIs), or both without NRTIs. Brachial artery flow-mediated dilation (FMD) and cardiovascular biomarker assessments were performed at baseline and at weeks 4 and 24. Ten haplogroup-defining mtDNA polymorphisms were determined. FMD and biomarker changes from baseline to week 24 by mtDNA variant were assessed using Wilcoxon rank-sum tests. Thirty-nine non-Hispanic white participants had DNA and 24-week data. The nonsynonymous m.10398A>G mtDNA polymorphism (N=8) was associated with higher median baseline adiponectin (5.0 vs. 4.2 μg/ml; p=0.003), greater absolute (−1.9 vs. −0.2 μg/ml) and relative (−33% vs. −3%) adiponectin decreases (p<0.001 for both), and lower week 24 brachial artery FMD (3.6% vs. 5.4%; p=0.04). Individual mtDNA haplogroups, including haplogroups H (N=13) and U (N=6), were not associated with adiponectin or FMD changes. In this small pilot study, adiponectin and brachial artery FMD on ART differed in non-Hispanic whites with a nonsynonymous mtDNA variant associated with several human diseases. These preliminary findings support the hypothesis that mtDNA variation influences metabolic ART effects. Validation studies in larger populations and in different racial/ethnic groups that include m.10398G carriers are needed.
Introduction
M
Mitochondrial DNA (mtDNA) encodes electron transport chain subunits that are critical for energy production, and mitochondrial function influences vascular health. 10 Combinations of single nucleotide polymorphisms (SNPs) within mtDNA allow for categorization of individuals into haplogroups. 11 Variation in mtDNA sequence has been implicated in aging and neurodegenerative diseases, 12 and more recently in type 2 diabetes mellitus 13 and metabolic syndrome. 14 Our group 15,16 and others 17 –19 have identified mtDNA variants that may influence metabolic ART toxicities in HIV-infected persons. Potential roles for mitochondria in endothelial dysfunction and atherosclerosis have been described, 20 and a recent study reported associations between high-dose statins, mitochondrial dysfunction, and impaired endothelial function. 21
AIDS Clinical Trials Group (ACTG) study A5142 was a randomized trial comparing class-sparing ART regimens for the initial treatment of HIV infection. 22 Metabolic results from A5142 demonstrated less limb fat loss by dual-energy X-ray absorptiometry (DEXA), but greater increases in fasting serum lipids among persons receiving NRTI-sparing ART over 96 weeks. 23 ACTG study A5152s was a substudy of A5142 designed to characterize endothelial function over the first 24 weeks of ART. Using brachial artery flow-mediated dilation (FMD), this study demonstrated impaired endothelial function at baseline, but an improvement of endothelial function at week 24 that was associated with the magnitude of HIV RNA reduction, and was similar across treatment arms. 24 Cardiovascular and metabolic biomarker data were also collected in A5152s. In this pilot study, we explored relationships between FMD, biomarkers, and mtDNA variants in a subgroup of these trial subjects, with the primary hypothesis that variation in mtDNA influences changes in endothelial function and biomarkers relevant for cardiovascular disease in HIV-infected individuals receiving ART.
Materials and Methods
Study subjects
This analysis included subjects from ACTG study A5142 (NCT #00050895), a multicenter, randomized, open-label trial that enrolled HIV-1-infected, ART-naive volunteers in the United States with plasma HIV-1 RNA≥2,000 copies/ml. 22 Subjects were randomized to one of three class-sparing ART regimens: lopinavir/ritonavir plus efavirenz (NRTI-sparing) or two NRTIs plus either lopinavir/ritonavir [non-NRTI (NNRTI)-sparing] or efavirenz (protease inhibitor-sparing). As described elsewhere, 22,23 investigator-selected NRTIs (zidovudine, stavudine, or tenofovir) were given with lamivudine if the subject was randomized to the NRTI-containing arms; randomization was stratified by NRTI choice. A subgroup of A5142 subjects from six U.S. sites also coenrolled in substudy A5152s (NCT #00050908) 24 and underwent additional cardiovascular risk assessments at baseline and 4 and 24 weeks after starting A5142 ART regimens. Exclusions for A5152s included known cardiovascular disease, diabetes mellitus, or current use of lipid-lowering medications, antioxidant vitamin supplementation, or hormones at greater than replacement doses. Pharmacologic treatment of diabetes mellitus, dyslipidemia, and changes in doses of angiotensin-converting enzyme inhibitors were not permitted on the study. Race/ethnicity categories were by self-report.
This pilot analysis focused on “white, non-Hispanics” as data on European mtDNA haplogroups from A5142 were available from our prior study (Fig. 1). 16 Protocols A5142, A5152s, and A5128 (the ACTG Human DNA Repository) were approved by institutional review boards at each study site, and subjects provided written informed consent. The Vanderbilt Committee for the Protection of Human Subjects and the ACTG approved the use of deidentified genetic and clinical data.
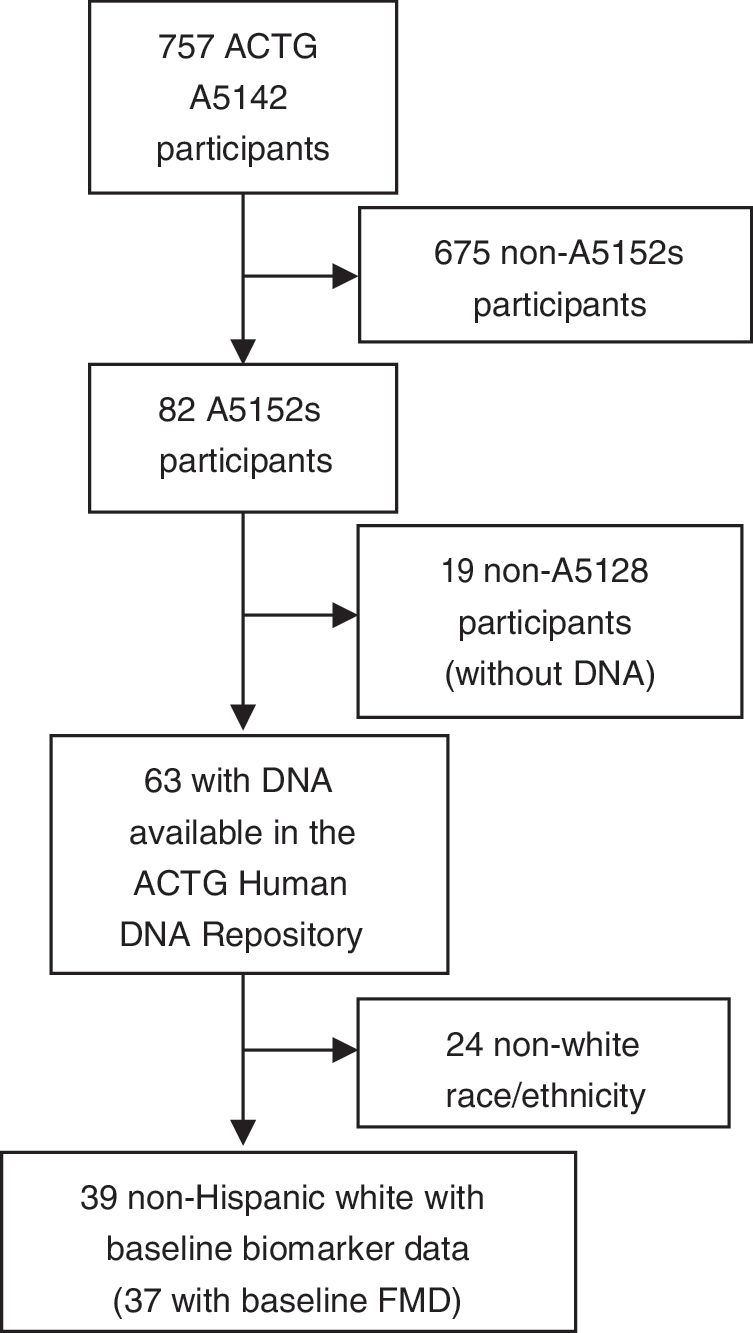
Flow diagram of study participants included in the primary ACTG study (A5142), substudy (A5152s), and the analyses presented here. ACTG, AIDS Clinical Trials Group; FMD, (brachial artery) flow-mediated dilation.
Brachial artery flow-mediated dilation
Subjects underwent FMD at baseline and weeks 4 and 24 on the same day as phlebotomy using recommended techniques, 25 and were required to be fasting and to not have used tobacco products for 8 h prior to the study. 24 Trained sonographers performed FMD assessments using standardized equipment at each site. Steps were taken to minimize inter- and intrasonographer variability, and measurements were performed by a single reader at a core reading center blinded to subject information and treatment arm. 24 In A5152s subjects who underwent repeat scans within 16 days of the week 24 scan (N=20), the difference in FMD was 0.26% (−0.43 to +0.72%, p=0.498). Blinded rereading of 57 scans revealed correlations of 0.97–0.99 (p<0.001) and small median differences of −0.14 to +0.09% at each week. 24
Cardiovascular and inflammatory biomarkers
Fasting blood samples were collected at baseline and weeks 4 and 24. Biomarker quantification (including adiponectin) was performed using Luminex bead array (Linco Research, St. Charles, MO) or ELISA (ALPCO Immunoassays, Salem, NH) at Northwestern University (Chicago, IL). Routine lipid, glucose, lipoprotein (a), and high-sensitivity C-reactive protein (hsCRP) quantification were performed at the ACTG Central Metabolic Laboratory (Quest Diagnostics, Baltimore, MD). Homeostasis model assessment-insulin resistance (HOMA-IR) was determined using [fasting insulin (μIU/ml)×fasting glucose (mg/dl)]/405.
DNA sequencing and mitochondrial haplogroup determination
DNA for these analyses was isolated from whole blood using PUREGENE (Gentra Systems Inc., Minneapolis, MN) under protocol A5128 (the ACTG Human DNA Repository). Genotyping was performed with the ABI PRISM 7900HT Sequence Detection System (Applied Biosystems Inc., Foster City, CA) using the 5′ nuclease allelic discrimination Taqman assay. Based on reported haplogrouping methods, 26,27 we characterized 10 mtDNA SNPs using probes and primers described previously. 15 In addition to haplogroup analyses described below, we compared biomarker and FMD outcomes by mtDNA variant (m.) 10398A>G (rs2853826) status. This SNP was chosen because it is one of the SNPs used to define haplogroups, is nonsynonymous (causing a threonine to alanine amino acid change in the NADH dehydrogenase subunit of Complex I), and has been associated with human diseases including metabolic syndrome. 28 Genotypic data were analyzed using ABI Sequence Detection System version 2.0 software and genotype calls were confirmed by visual inspection of plots.
Statistical analysis
Simple proportions are used to describe demographic and genetic data. Medians and interquartile ranges (IQR) are presented for continuous data. Fisher's exact or Pearson Chi-squared tests and Wilcoxon rank-sum test were used for comparisons of categorical and continuous covariates by haplogroup or m.10398A>G, respectively. Stata SE version 10 (Stata Corp., College Station, TX) was used for statistical analyses. We did not formally correct for multiple comparisons in this pilot study, but did explore corrected associations post hoc.
Results
Of 82 A5152s subjects, 63 had DNA available (Table 1 and Fig. 1). This analysis included the 39 subjects self-described as non-Hispanic white and with baseline biomarker data; they had a median age of 36 years, were predominantly male (92%), and had a median CD4 count of 273 cells/mm3. These factors did not differ statistically between this subgroup and the non-Hispanic white A5142 subjects included in prior mtDNA analyses 16 (data not shown). Median body mass index (BMI) was 25 kg/m2 and median fasting high-density lipoprotein (HDL) and low-density lipoprotein (LDL) cholesterol concentrations at baseline were 31 and 88 mg/dl, respectively. Similar numbers of subjects were randomized to protease inhibitor-sparing (N=11), NRTI-sparing (N=13), or NNRTI-sparing (N=15) ART. Of those who received NRTIs, six received stavudine and 10 each received zidovudine and tenofovir. Individual mtDNA haplogroup sample sizes were small (Table 1). Most subjects (33%) belonged to European haplogroup H. Of those remaining, six (15%) belonged to haplogroup U, and haplogroups J, T, and X included four persons (10%) each.
All m.10398 comparisons are nonsignificant by Wilcoxon rank-sum or Fisher's exact/chi2 tests (all p>0.15).
m.10398 status could not be determined in one subject.
ART, antiretroviral therapy; HDL, high-density lipoprotein; HOMA-IR, homeostasis model assessment-insulin resistance; LDL, low-density lipoprotein; mtDNA, mitochondrial DNA; NRTI, nucleoside reverse transcriptase inhibitor; NNRTI, non-NRTI; XR, extended release.
Overall median (IQR) baseline FMD for 37 subjects was 3.3% (1.9, 4.7). After 4 and 24 weeks of ART, median FMD increased to 4.8% (3.0, 6.4) and 5.1% (3.0, 6.7), respectively (Table 2). This represented median percentage increases of 43% and 36%, respectively. No haplogroup demonstrated significant differences in FMD responses over 4 or 24 weeks of ART (Supplementary Table S1; Supplementary Data are available online at
FMD data were not available at each time point in all subjects.
Wilcoxon rank-sum test.
m.10398 status could not be determined in one subject.
FMD, flow-mediated dilation.
Baseline adiponectin levels were higher among subjects with m.10398G (p=0.003; Table 2). At week 24, however, subjects with m.10398G tended to have lower adiponectin levels (p=0.08), and had experienced significantly greater absolute and relative decreases in adiponectin (p=0.0005 for both; Table 2). These associations remain statistically significant after conservative Bonferroni adjustment for multiple comparisons [p=0.05/(6 mtDNA variants×7 biomarkers×2 time points)=0.0006]. Of note, there were 13 subjects (43%) with an increase in adiponectin from baseline to week 24; none carried the m.10398G allele (p=0.03). Although the small sample size precluded comprehensive multivariate adjustment, in individual single-covariate linear regression models, relationships between m.10398G and 24-week decrease in adiponectin remained significant with adjustment for baseline age, BMI, DEXA measures, randomized study arm, thymidine analog NRTI (zidovudine or stavudine) use, baseline or 24-week HOMA-IR (all p≤0.001), and baseline adiponectin (p=0.04). To further assess the impact of ART on these associations, we compared 24-week adiponectin changes by the m.10398 allele in various treatment groups. Across the randomized class-sparing arms and for those who received a thymidine analog NRTI (stavudine or zidovudine), median 24-week adiponectin changes ranged from −5% to +6% in the m.10398A group, compared with −27% to −55% among the m.10398G group. Wilcoxon p-values ranged from 0.005 to 0.1 for these comparisons.
Regarding other biomarkers, there were marginally statistically significant associations (uncorrected) among persons with haplogroup H (see Supplementary Table S1). These included lower baseline and 24-week asymmetrical dimethyl arginine (ADMA) levels (p=0.02 for both), and lower baseline lipoprotein (a) (p=0.03) levels. Four persons belonging to haplogroup T had a greater percentage increase in hsCRP at week 4 than non-T individuals (p=0.04), and six persons belonging to haplogroup U had significant increases in HOMA-IR at week 24 versus non-U individuals (p=0.04). None of these associations remained statistically significant after adjustment for multiple comparisons.
Discussion
This pilot study of ART-naive clinical trial subjects identified associations between a nonsynonymous mtDNA SNP and adiponectin changes consistent with metabolic dysfunction after 24 weeks of ART. To our knowledge, this represents the first reported association between an mtDNA variant and FMD, and the second published report of an association between mtDNA and adiponectin in HIV-infected persons. 17 The m.10398A>G variant that was associated with a decreased adiponectin on ART results in a threonine to alanine amino acid change in the NADH dehydrogenase subunit 3 of Complex I, and alters in vitro mitochondrial measures (e.g., mitochondrial matrix pH and calcium concentration). 29 It has also been studied in human diseases and aging, 30,31 but risk associations have been inconsistent. In a Chinese population, this variant was associated with an increased risk for metabolic syndrome. 28 Interestingly, baseline adiponectin was higher in persons with the m.10398G allele. This apparent paradoxical association might suggest a compensatory response to chronic HIV infection that precedes a greater decrease on ART. Alternately, an effect of ART on adiponectin may have been most pronounced in those persons with the highest baseline levels. These interpretations are speculative and require further study.
Adiponectin is an adipose tissue-derived peptide hormone that mediates energy balance and metabolism, and has antioxidant properties. In HIV-negative populations, low adiponectin is associated with cardiovascular risk factors 32,33 and disease. 34 Recent in vitro data suggest adiponectin synthesis is dependent on mitochondrial function. 35 Adipokines are dysregulated in HIV-infected persons. 36 –38 Lean HIV-positive, ART-treated men had adiponectin levels similar to obese, insulin-resistant HIV-negative men, and those with extremity fat loss had even lower levels. 36 Baseline trunk and limb fat by DEXA did not differ by m.10398 allele in our analyses (Table 1). DEXA data from A5142 were not available at week 24, but 48-week limb fat changes did not differ by m.10398, and 24-week adiponectin levels/changes were not correlated with 48-week trunk or limb fat changes (data not shown), suggesting that adiponectin changes were not merely markers of adipose changes in this population.
A recent cross-sectional analysis of Spanish HIV/HCV coinfected subjects reported an association between European mtDNA haplogroups and serum adiponectin. 17 In that study, subjects belonging to the JT clade (including persons with both m.10398A and G alleles) had significantly lower adiponectin levels while on ART than those belonging to the HV clade (including only the m.10398A allele). 17 Those subjects had>5 years median ART exposure, thus results cannot be directly compared with a clinical trial population followed during the first 24 weeks of ART. However, if the 24-week adiponectin decrease among ACTG m.10398G allele subjects persisted, the findings of these two studies may be consistent. Our analysis included eight individuals belonging to the JT clade. Absolute and relative 24-week adiponectin decreases tended to be greater in these individuals than in those belonging to the HV clade (p=0.05; data not shown), consistent with the m.10398 analyses and with the prior report. 17
The small sample size of our study precluded comprehensive haplogroup analyses, limited our capacity to adjust for potential confounders, and may have led to missed associations. The m.10398G allele is present in European haplogroups I, J, and K, thus a careful analysis in a larger population might better determine whether m.10398G is a marker for other variant(s) shared across multiple haplogroups or is confined to a single haplogroup. Diabetes was an exclusion from A5152s, and development of overt diabetes during the first 24 weeks of ART was not observed. Fasting insulin levels and HOMA-IR did not change significantly over 24 weeks of ART, and did not differ by m.10398 allele (data not shown). Due to the small sample size, we were also unable to fully assess for interactions between ART and mtDNA variants in robust adjusted models. However, in the primary A5152s analysis, randomized ART arm was not associated with 24-week changes in adiponectin or FMD, 24 and neither randomized ART nor NRTI differed significantly by m.10398 allele in the subgroup analyzed here (Table 1). Associations between 24 week adiponectin change and m.10398G persisted across multiple ART subgroups and with single-covariate adjusted models.
In summary, these pilot data support a previously published association between an mtDNA SNP and adiponectin levels 17 and suggest a potentially novel association with poorer FMD on ART. The m.10398G allele was associated with adverse adiponectin and FMD phenotypes in this population. These findings suggest a potential role for mtDNA variation in metabolic/cardiovascular complications in HIV-infected, ART-treated persons. Although not definitive, these data also suggest a possible mechanistic link between mtDNA variation and metabolic derangements through relatively short-term (24 week) effects on adiponectin. Analyses in larger and more ethnically diverse populations are needed to define whether the m.10398G allele and/or other variants are associated with longer-term cardiovascular measures, biomarkers, or outcomes in these populations.
Footnotes
Acknowledgments
The authors gratefully acknowledge the study volunteers who participated in A5142, A5152s, and the ACTG Human DNA Repository protocol A5128. We additionally acknowledge A5142 study co-chairs Richard Haubrich, MD (University of California-San Diego, San Diego, CA) and Sharon Riddler, MD (University of Pittsburgh, Pittsburgh, PA), staff members at A5152s study sites, and other members of the A5152s study team: Lauren Kamarow, MS (Harvard School of Public Health, Boston, MA); Robert A. Parker, ScD (Harvard School of Public Health, Boston, MA); Judith S. Currier, MD (University of California-Los Angeles, Los Angeles, CA); Carl J. Fichtenbaum, MD (University of Cincinnati Medical Center, Cincinnati, OH); Carol K.C. Mitchell, PhD (University of Wisconsin School of Medicine and Public Health, Madison, WI); Robert L. Murphy, MD (Northwestern University Feinberg School of Medicine, Chicago, IL); and Kathleen Squires, MD (University of Southern California, Los Angeles, CA).
The project described was supported by Award Number U01AI068636 from the National Institute of Allergy and Infectious Diseases and was supported by the National Institute of Mental Health (NIMH) and the National Institute of Dental and Craniofacial Research (NIDCR). The ACTG Human DNA Repository is also supported by CTSA award No. UL1TR000445 from the National Center for Advancing Translational Sciences. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institute of Allergy and Infectious Diseases, the National Center for Advancing Translational Sciences, or the National Institutes of Health.
A5142 (NCT #00050895) was also supported by Grants AI 068636 (AIDS Clinical Trials Group Central Grant), AI 068634, AI 069471, AI 27661, AI 069439, AI 25859, AI 069477, AI 069513, AI 069452, AI 27673, AI 069470, AI 069474, AI 069411, AI 069423, AI 069494, AI 069484, AI 069472, AI 38858, AI 069501, AI 32783, AI 069450, AI 32782, AI 069465, AI 069424, AI 38858, AI 069447, AI 069495, AI 069502, AI 069556, AI 069432, AI 46370, AI 069532, AI 46381, AI 46376, AI 34853, AI 069434, AI 060354, AI 064086, AI 36214, AI 069419, AI 069418, AI 50410, AI 45008, AI 25915, RR 00075, RR 00032, RR 00044, RR 00046, RR 02635, RR 00051, RR 00052, RR 00096, RR 00047, RR 00039, RR 16176, and DA 12121 from the National Institute of Allergy and Infectious Disease, National Institutes of Health. The collaborating pharmaceutical companies provided lopinavir–ritonavir (Abbott), efavirenz and stavudine (Bristol-Myers Squibb), and tenofovir DF (Gilead).
Other support included NIH Grants AI 64086, AI 36214, AI 69432, AI 60484, MD 007601, MD 000173, GM 103341, and AI 77505.
These data were presented in part at the 6th International Conference on HIV Pathogenesis, Treatment, and Prevention, July 17–20, 2011, Rome, Italy (Abstract #A-361-0075-03110).
Author Disclosure Statement
T.H. has served as principal investigator of research grants to Vanderbilt University from Merck & Co. J.H.S. has served on Data and Safety Monitoring Committees for clinical trials of lipid-lowering medications sponsored by Abbott and Lilly, and for a diabetes medication sponsored by Takeda. M.P.D. has received research funding from Gilead Sciences and Glaxo Smith Kline. D.W.H. has been supported by research grants to Vanderbilt University from Boehringer-Ingelheim, Gilead Sciences, and Merck & Co. The other authors report no disclosures.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.