
Abstract
Increasing evidence has suggested that HIV infection severely damages the Vγ2Vδ2 (Vδ2) T cells that play an important role in the first-line host response to infectious disease. However, little is known about Vδ2 T cell-mediated antibody-dependent cell-mediated cytotoxicity (ADCC) in HIV disease. We found that although the CD16+ Vδ2 T cell subset hardly participated in phosphoantigen responses dominated by the CD16− Vδ2 T cell subset, the potency of the ADCC function of Vδ2 T cells was correlated with the frequency of the CD16+ subset. Thus, two distinct and complementary Vδ2 T cell subsets discriminated by CD16 were characterized to explore the respective impacts of HIV-1 infection on them. HIV-1 disease progression was not only associated with the phosphoantigen responsiveness of the CD16− Vδ2 subset, but also with the ability of the CD16+ Vδ2 subset to kill antibody-coated target cells. Furthermore, both of the two Vδ2 functional subsets could be partially restored in HIV-infected patients with antiretroviral therapy. Notably, in the context of an overall HIV-mediated Vδ2 T cell depletion, despite the decline of phosphoantigen-responsive CD16− Vδ2 cells, CD16+ Vδ2 cell-mediated ADCC was not compromised but exhibited a functional switch with dramatic promotion of degranulation in the early phase of HIV infection and chronic infection with slower disease progression. Our study reveals functional characterizations of the two Vδ2 T cell subsets with different activation pathways during HIV-1 infection and provides a rational direction for activating the CD16+ Vδ2 T cells capable of mediating ADCC as a means to control HIV-1 disease.
Introduction
H
Antibody-dependent cell-mediated cytotoxicity (ADCC), which relies on specific antibodies and Fc receptor-bearing effector cells for a proper antiviral response, plays an important role in controlling HIV infection. Previous studies have documented compromised ADCC responses in progressive HIV-1 infection from the perspective of HIV-specific antibodies. 16 –18 Furthermore, the RV144 Thai trial demonstrated that nonneutralizing antibodies elicited by the vaccination might protect against HIV acquisition, potentially preventing infection through the ADCC mechanism. 19 Effector cells, including natural killer (NK) cells, Vδ2 T cells, and monocytes, are able to recognize the antibodies bound to infected cells through a low-affinity Fc receptor for IgG, called FcγRIIIa (CD16). Recently, impaired ADCC function of NK cells was observed in HIV-infected individuals, 20 which indicates that in addition to antibodies, the capacity of effector cells to respond to target cells should also be studied when evaluating ADCC activity. Similar to NK cells, Vδ2 T cells also express CD16 that can be used for ADCC, but little is known about the Vδ2 T cells with respect to their activity as ADCC effectors during HIV-1 disease progression.
It has been reported that memory Vδ2 T cells can be divided into two subsets with unique effector functions based on the expression of CD16 and these subsets represent different pathways of maturation for circulating Vδ2 T cells. 21 Thus, Vδ2 T cells comprise a number of distinct effector subsets and likely have complex activities during HIV infection. We propose that one of these activities is ADCC, which is mediated by a unique Vδ2 T cell subset with CD16 expression. Given the divergent activation pathways, we wanted to study in detail the two Vδ2 T cell subsets discriminated by CD16 from uninfected controls, HIV-1-positive subjects without treatment at different stages of infection and HIV-1-positive subjects receiving highly active antiretroviral therapy (HAART). We examined the magnitudes of IPP-induced activation in the two subsets across different cohorts. We also focused on the Vδ2 T cell-mediated ADCC in terms of degranulation and cytokine production, and measured CD16 expression on Vδ2 T cells to see if the baseline frequency of the Fc receptor correlated with their ADCC function. These studies offered us a comprehensive view of Vδ2 T cell activity during HIV infection with an in-depth analysis of these two complementary effector subsets.
Materials and Methods
Subjects
A total of 91 subjects were recruited for the study, including 21 healthy HIV-1-negative controls, 18 treatment-naive early HIV-1-infected subjects, 42 treatment-naive chronic HIV-1-infected subjects, and 10 HIV-1-infected subjects receiving HAART (Table 1). All the treatment-naive HIV-1-infected subjects were from a cohort of HIV-1-positive men who have sex with men (MSM) infected for different lengths of time. The MSM cohort was recruited at Chaoyang CDC in China. The estimated time of HIV infection was interpolated as the midpoint between the negative and HIV-1 seroconversion. Early HIV-1 infection was defined as infection within 6 months (2–6 months). Chronic HIV-1-infected subjects had been infected for at least 1 year (12–36 months); they were further divided into three subgroups based on CD4 T cell counts. The cohort of treated HIV-1-infected subjects was recruited at Dehui CDC in China and had received HAART for at least 1 year (12–72 months) with prolonged limited viral loads <100 copies/ml and with a CD4 T cell count >300 cells/μl. Ethical approval was granted by the Institutional Review Board of the National Center for AIDS/STD Control and Prevention, Chinese Center for Disease Control and Prevention. All patients gave informed consent for participation in the study.
Data are presented as median (range); NA, not applicable.
Cell processing
Peripheral blood mononuclear cells (PBMCs) were isolated from EDTA-treated venous blood by Ficoll Hypaque centrifugation (Sigma) within 6 h of sample collection. After cell counts and a viability check, PBMCs were resuspended at a density of 1×106 cells/ml in R-10 medium [RPMI-1640 medium supplemented with 10% fetal bovine serum (FBS), 2 mmol
Phenotypic analysis of Vδ2 T cells
Cell surface staining was performed using the following monoclonal antibodies: CD3 (PerCP conjugate, Clone SK7, BD Biosciences), Vδ2 (PE conjugate, CloneB6, BD Biosciences), and CD16 (PB conjugate, Clone CB16, eBioscience). PBMCs (1×106) were used for each staining. After staining, cells were washed with phosphate-buffered saline (PBS), fixed with 2% paraformaldehyde (PFA) solution, and collected on BD FACS Aria. Data were analyzed by FlowJo software (TreeStar Inc.).
Vδ2 T cell stimulation by IPP
PBMCs were cultured at 1.5×106 cells/well in R-10 medium in 96-well round-bottomed plates and stimulated with medium alone or isopentenyl pyrophosphate (Sigma), respectively. CD107a (APC conjugate, Clone H4A3, BD Biosciences), Golgi-Stop (BD Biosciences), and Brefeldin A (Sigma) were added to each well. Following an 11-h incubation at 37°C with 5% CO2, PBMCs were collected, washed with PBS, and stained with monoclonal antibodies CD3, Vδ2, and CD16. Cells were then permeabilized using Caltag Fix & Perm (Invitrogen) and intracellular cytokine staining was carried out for interferon (IFN)-γ (Alexa700 conjugate, Clone B27, BD Biosciences). After staining, cells were washed with PBS and fixed in 2% PFA solution. Labeled cells were acquired on BD FACS Aria and analyzed by FlowJo software.
Vδ2 T cell-mediated ADCC assay
A single cell line p815 (mouse leukemic cell line) coated with rabbit antibodies specific to p815 cells (Accurate Chemical & Scientific Corporation) was used as target cells. Target cells were prepared by culturing p815 cells with p815 antibodies in R-10 medium for 1 h at 37°C with 5% CO2 prior to ADCC assay. PBMCs were used as effector cells and stimulated with R-10 medium, uncoated p815 cells, or antibody-coated (Ab-coated) p815 cells, respectively. CD107a, Golgi-Stop, and Brefeldin A were added to each well and samples were incubated for 11 h at 37°C with 5% CO2. After incubation, cells underwent surface staining with monoclonal antibodies CD3, Vδ2, and CD16 and intracellular staining for IFN-γ. After staining, cells were washed with PBS and fixed in 2% PFA solution. Labeled cells were acquired on BD FACS Aria and analyzed by FlowJo software.
Cell counts of CD4+ T cells and Vδ2 T cells
The cell counts of CD4+ T cells and CD3+ T cells were measured in EDTA-treated venous blood with FACS Calibur TruCount tube (Becton Dickinson), staining by multicolor antibodies that included CD3-FITC, CD4-PE, CD8-APC, and CD45-PerCP (Becton Dickinson). Labeled cells were acquired on FACS Calibur. Data were analyzed by MultiSETTM software (BD Bioscience). Cell counts of Vδ2 T cells or functional Vδ2 T cells were determined based on their percentage among CD3+ T cells and CD3+ T cell counts. 11
Viral load quantification
Plasma viral RNA was quantified in EDTA-treated venous blood by fluorescent real-time polymerase chain reaction (PCR). Plasma viral loads were analyzed by the Amplicor ultrasensitive assay (Hoffmann-La Roche) with a detection limit of 50 viral RNA copies/ml of plasma.
Statistical analysis
All the statistical and graphic analyses were performed using GraphPad Prism software (GraphPad Software Inc.) and Sigma Plot software (SPSS Inc.). Statistical comparisons between different groups were made using one-way ANOVA. p values of pairwise comparisons were adjusted for multiple comparisons using Dunn's method. Spearman rank order correlation was used to analyze the correlation between variables. p values less than 0.05 were considered to be significant.
Results
Two different patterns of activation of Vδ2 T cells discriminated by CD16 expression
Vδ2 T cells can be specifically and efficiently activated by phosphoantigens, such as IPP, in a TCR-dependent manner. 8 FcγRIIIa (CD16), a low-affinity receptor that binds only to clustered IgG bound to cell surface, is found on Vδ2 T cells and can mediate ADCC activity. 22 In this study, we measured both the TCR-dependent IPP response and CD16-dependent ADCC function of Vδ2 T cells. The method for measuring NK cell-mediated ADCC 20 was applied to Vδ2 T cells in this study. We used a single target cell line p815 coated with p815-specific antibodies as Fc target cells (Fc targets) to induce the ADCC activity of Vδ2 T cells. Similar to NK cells, we found an obvious decline of CD16 expression on Vδ2 T cells following stimulation with Ab-coated p815 cells, but not in the presence of medium, IPP stimuli, or p815 cells alone (Fig. 1a).

Response of Vδ2 T cells to isopentenyl pyrophosphate (IPP) and Fc targets stimulation. Levels of CD16 expression in Vδ2 T cells (upper panel) as well as CD107a and IFN-γ (lower panel) were measured in the assay
CD107a, a degranulation marker, is a necessary indicator for cellular cytotoxicity assays. 23 In this study, CD107a and IFN-γ were both used to evaluate the cytotoxicity and cytokine-producing capacity of Vδ2 T cells, respectively. After treating with IPP or Ab-coated p815 cells, a significant expression of CD107a and IFN-γ was detected on Vδ2 T cells, while medium alone and uncoated p815 cells did not induce any response (Fig. 1a). In particular, IPP induced degranulation and IFN-γ production mostly by the CD16− Vδ2 T cell subset (% median for CD16− subset in IPP-responsive Vδ2 T cells, 98.82% of CD107a+ Vδ2 T cells and 99.40% of IFN-γ+ Vδ2 T cells), and rarely by CD16+ Vδ2 T cells (Fig. 1a), and the IPP response dominated by CD16− Vδ2 T cells was observed across cohorts independent of disease progression (Fig. 2b and c).
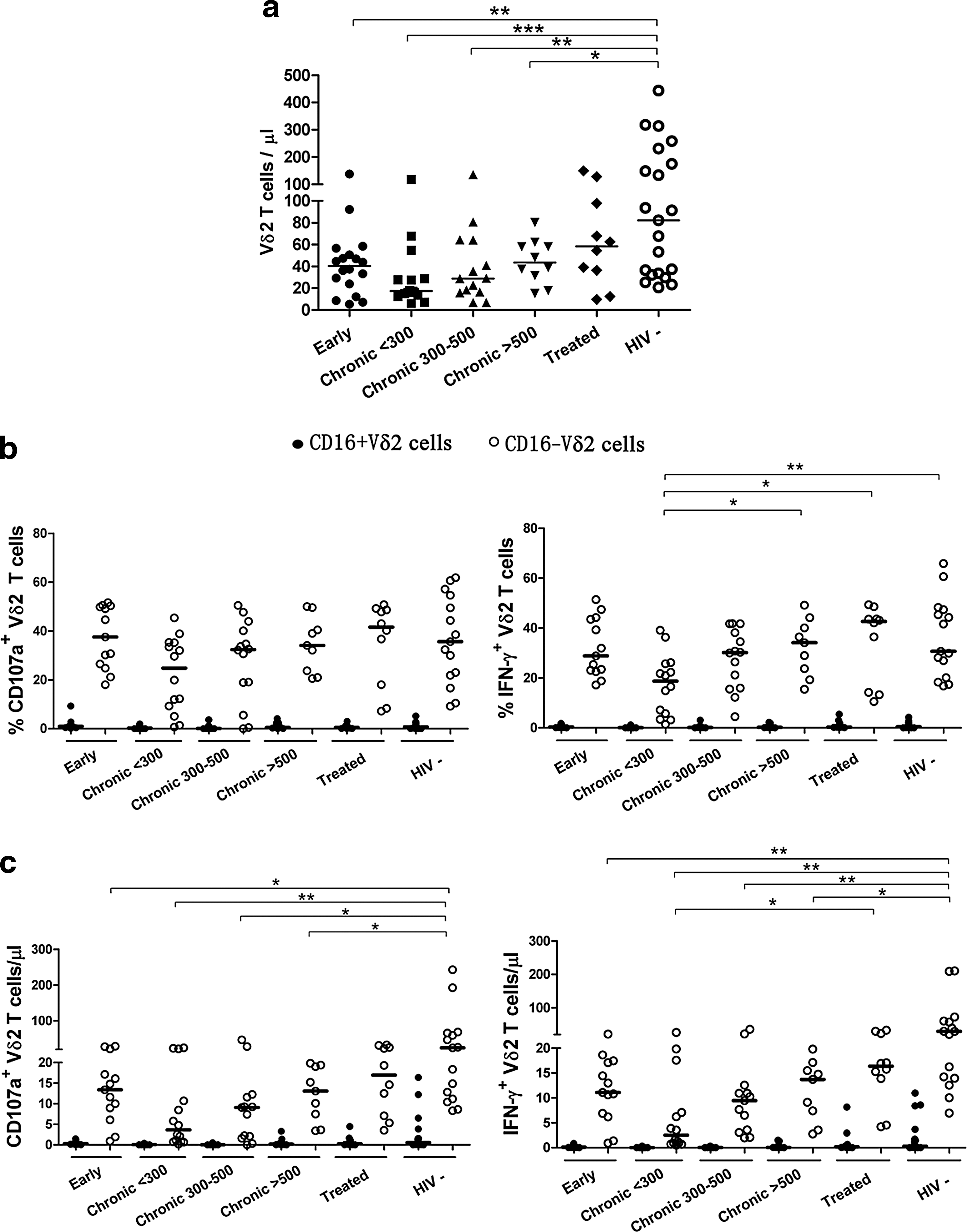
IPP-induced responses of Vδ2 T cells based on CD16 expression during different stages of HIV-1 infection. Dot plots show the differences in Vδ2 T cell counts
Meanwhile, stimulation with Fc targets resulted in a profound activation of Vδ2 T cells as seen by increased degranulation and IFN-γ production, implicating ADCC as a key effector function of Vδ2 T cells. Given that CD16 is the only Fc gamma receptor reported on Vδ2 T cells, we expected that CD16+ Vδ2 T cells, and not CD16− Vδ2 T cells, were the ones exerting ADCC. Subsequent regression analysis of the cohorts revealed a strong correlation between the CD16+ Vδ2 T cells and Fc target responders, CD107a+ Vδ2 T cells and IFN-γ+ Vδ2 T cells, in terms of either frequencies or cell counts (Fig. 1b). These data indicated that an increased baseline frequency of CD16+ Vδ2 T cells allowed them to better respond to Fc targets. This is consistent with the report showing that CD16 expression closely correlates with ADCC function in NK cells. 20 Thus, we defined and characterized Vδ2 T cells on the basis of CD16 expression to study the respective impact of HIV-1 infection on the immune response of two functional Vδ2 T cell subsets with distinct antigen recognition pathways.
Compromised IPP-induced response of Vδ2 T cells dominated by the CD16− subset in HIV-1 infection
Although a decreased cell number and impaired phosphoantigen response of Vδ2 T cells in HIV disease have been reported, 11,13 the detailed impact of HIV infection on CD16+ and CD16− Vδ2 T cell subsets is not yet clear. In our cohorts, untreated subjects at different stages of chronic HIV-1 infection were divided into three subgroups based on CD4 T cell counts (cells/μl): CD4<300, 300≤CD4 ≤500, and CD4 >500. During early HIV-1 infection, when a rapid decline of CD4 T cells was observed in comparison with healthy controls (p<0.001), Vδ2 T cell counts had been reduced significantly and the loss of Vδ2 T cells was also observed in untreated chronic HIV-1 infection (p<0.05; Fig. 2a). In terms of IPP responders, dramatic declines of CD107a+ CD16− Vδ2 T cells and IFN-γ+ CD16− Vδ2 T cells (p<0.05 for both) were observed in early HIV infection compared to healthy controls (Fig. 2c). In addition, there was an overall loss of IPP-responsive CD107a+ and IFN-γ+ CD16− Vδ2 T cells in chronic untreated HIV-infected individuals when compared to healthy subjects (p<0.05 for all). The chronic untreated patients with CD4<300 had a decreased frequency of IFN-γ+ CD16− Vδ2 T cells in response to IPP compared to patients with higher CD4 T cell counts (p<0.05; Fig. 2b). Furthermore, we observed an elevated percentage and cell count of IPP-responsive IFN-γ+ CD16− Vδ2 T cells from treated individuals in comparison to untreated individuals with CD4 <300 (p<0.05 for both).
Consistent with one previous study
11
, IPP-responsive Vδ2 T cells were positively correlated with CD4 T cell counts (r=0.369, p=0.027 for CD107a+ Vδ2 T cells and r=0.431, p=0.009 for IFN-γ+ Vδ2 T cells) and inversely correlated with viral loads (r=−0.401, p=0.035 for CD107a+ Vδ2 T cells and r=−0.460, p=0.014 for IFN-γ+ Vδ2 T cells) in chronic HIV infection (Supplementary Fig. S1a; Supplementary Data are available online at
Defective ADCC function of Vδ2 T cells during chronic HIV-1 infection
CD16, which may serve as a predictor for FcR-mediated activation of Vδ2 T cells, was examined during the course of HIV-1 infection. As shown in Fig. 3a, there was no difference in the overall proportions of CD16+ Vδ2 T cells. However, there were notable differences in the CD16+ Vδ2 T cell counts between the two chronic subgroups with CD4 ≤500 and healthy controls (p<0.05 for both), while the CD4 >500 subgroup maintained the CD16+ Vδ2 T cell level (quantity median, 11.78 cells/μl; range, 3.92–13.43 cells/μl) compared to healthy controls (quantity median, 11.94 cells/μl; range, 4.09–39.51 cells/μl). Given the strong correlation between CD16 expression and Vδ2 T cell-mediated ADCC, the decline of CD16+ Vδ2 T cells could result in the Vδ2 T cells being incapable of exerting ADCC in chronic HIV-1 infection with fast disease progression.

CD16+ Vδ2 T cell-mediated ADCC responses during different stages of HIV-1 infection. The dot plots depict changes in frequencies and cell counts of CD16+ Vδ2 T cells
In the Vδ2 T cell-mediated ADCC assay, although there was a sharp reduction of Vδ2 T cells after HIV-1 infection, a modestly higher percentage as well as nearly equal number of antibody-dependent cytotoxic Vδ2 T cells were found in early-infected subjects (frequency median, 7.83%; range, 3.26–15.36%) (quantity median, 2.09 cells/μl; range, 0.76–7.49 cells/μl) as compared to healthy controls (frequency median, 3.57%; range, 0.28–10.17%) (quantity median, 1.41 cells/μl; range, 0.10–7.34 cells/μl) (Fig. 3b and c). In chronic HIV-1 infection, Vδ2 T cell-mediated ADCC was compromised most severely in the chronic subgroup with CD4 <300 in all respects when compared to both the chronic subgroup with CD4 >500 and healthy controls (p<0.05). Notably, CD107a+ Vδ2 T cells responding to Fc targets were not compromised in the CD4>500 subgroup. The median percentage of CD107a+ Vδ2 T cells in the CD4>500 subgroup (frequency median, 11.39%; range, 4.56–15.42%) was over 3-fold higher than in healthy controls, and the median quantity of CD107a+ Vδ2 T cells in the CD4>500 subgroup (quantity median, 3.39 cells/μl; range, 1.66–7.18 cells/μl) was slightly higher than in healthy controls. Higher cell counts of degranulated and cytokine-producing Vδ2 T cells responding to Fc targets were observed in chronic treated individuals when compared with chronic untreated individuals with CD4<300 (p<0.05).
The ADCC functional Vδ2 T cells exhibited a positive correlation with CD4 T cell counts (r=0.518, p<0.001 for CD107a+ Vδ2 T cells and r=0.624, p<0.001 for IFN-γ+ Vδ2 T cells) and an inverse correlation with viral loads (r=−0.378, p=0.043 for CD107a+ Vδ2 T cells and r=−0.461, p=0.012 for IFN-γ+ Vδ2 T cells) in chronic subjects (Supplementary Fig. S1b), which was consistent with the varying levels of ADCC activities at different stages of chronic HIV-1 infection. In accordance with the IPP-activated Vδ2 T cells, the individuals with CD4<300 during early HIV-1 infection had much higher frequencies and cell counts, both with respect to degranulated and cytokine-producing Vδ2 T cells, than the chronic subgroup with CD4<300 (p<0.05 for all; Supplementary Fig. S2b). No relationships were observed between the ADCC functional Vδ2 T cells and CD4 T cell counts or viral load during early HIV-1 infection (Supplementary Fig. S1b).
Functional profile of Vδ2 T cells in response to IPP and Fc targets
We next analyzed the functional repertoires of Vδ2 T cells activated by IPP or Fc targets in both HIV-infected and healthy subjects (Fig. 4). Following stimulation with IPP, the dual-functional cells with expression of CD107a and IFN-γ dominated the functional proportion independent of HIV-1 infection. Significant expansion of the monoexpression of CD107a was found in Vδ2 T cells responding to IPP in early HIV-infected subjects compared to healthy controls (p<0.001).

Different functional profiles of IPP-responsive and ADCC-mediating Vδ2 T cells in HIV infection. The pies depict the two functional profiles of Vδ2 T cells in response to IPP and Fc targets, respectively, among individuals at different stages of HIV-1 disease. Vδ2 T cells were divided into three subpopulations based on their production of CD107a and IFN-γ individually or in combination (nonresponsive cells were excluded for the analyses). Median values for individuals from different populations are shown.
Different from the profile of the IPP-responsive Vδ2 T cells, the functional division between the degranulated and cytokine-secreting subset was much clearer in ADCC activity. Strikingly, the Vδ2 T cells from the HIV-infected and HIV-uninfected groups displayed an almost reverse ratio of the two monofunctional subgroups in response to Fc targets. Significant differences in the frequencies of monoexpression of CD107a on Vδ2 T cells were found between healthy controls and the two untreated HIV-infected subgroups, including the early HIV-infected subgroup (p<0.001) and the chronic subgroup with CD4>500 (p<0.05). The great degranulation capacity of Vδ2 T cells after HIV infection indicates that CD16-dependent ADCC activity in Vδ2 T cells might contribute to the killing of HIV-infected cells.
Discussion
Here we have demonstrated that irrespective of HIV-1 infection status, CD16− and CD16+ Vδ2 T cell subsets performed different functions in response to various stimuli, suggesting the existence of distinct activation pathways. This result is consistent with the functions of Vδ2 T effector memory subsets discriminated by CD16 expression, including Vδ2 TEMh cells, which lack CD16 expression and respond to TCR-dependent phosphoantigen stimulation, and Vδ2 TEMRA cells, which mediate ADCC function via CD16. 21 Previous studies have investigated Vδ2 T cell activation in response to phosphoantigen and revealed a close relationship between cell-surface TCR and phosphoantigen recognition. 8 –10
Of importance, our data show that the baseline frequency of CD16+ Vδ2 T cells correlated with the capacity of the Vδ2 T cell pool to respond to Fc targets. Moreover, when comparing the different patient cohorts, a significant decline in CD16+ Vδ2 T cells was observed specifically in subjects with faster disease progression compared to healthy controls, while this decline was not observed in patients with better controlled HIV infection. In addition, this preserved frequency of CD16+ Vδ2 T cells in better controlled HIV+ patients was associated with a higher capacity of the Vδ2 T cell population to exert ADCC. This suggests that CD16+ Vδ2 cells (possibly of the TEMRA phenotype) could be significantly contributing to the control of HIV infection via their strong capacity to exert ADCC. However, the observed decline of CD16+ Vδ2 T cells in HIV infection begs the question of what is the mechanism of this depletion.
One possibility includes CD16 downregulation of otherwise CD16+ Vδ2 T cells by cell surface proteolytic cleavage following activation, as has been described in NK cells, 20,24,25 and is inferred from our in vitro Vδ2 T cell-mediated ADCC assay (although the likelihood that Fc targets might block the detection of CD16 could not be excluded); another possibility is the progressive destruction of CD16+ Vδ2 T cells by HIV virus-mediated cytopathic effects, analogous to the pronounced damages to the TCR γ-2 chain repertoire seen in HIV viremic patients. 26 The mechanism of the CD16+ Vδ2 T cell decline needs further study, and may help to guide research aimed at improving the efficacy of Vδ2 T cell-mediated ADCC activity.
Consistent with previous studies, 13,27 we found the Vδ2 T cells declined at least as rapidly as CD4 T cells in early HIV infection, accompanied by the decreased IPP-responsive cells in the CD16− Vδ2 T cell subset. However, with a strong capacity for degranulation, the CD16+ Vδ2 T cells from the early-infected group maintained cytotoxic Vδ2 T cells as the potentially efficient ADCC effectors. The shift in ADCC functional repertoire with dominant degranulation, which irreversibly continued throughout HIV-1 infection, might be the result of CD16+ Vδ2 T cell effectors in response to HIV through ADCC.
Recently, the HIV envelope-mediated, CCR5/α4β7-dependent killing mechanism has been revealed by Pauza's group to explain the depletion of Vδ2 T cells. They also observed the highest level of HIV envelope-induced cell death in Vδ2 T cells when exposed to phosphoantigen, which suggests that phosphoantigen activation might be the reason for the specific loss of the Vγ2Jγ1.2 Vδ2 T cell subpopulation. 27 This finding may partially explain the different status of the two functional Vδ2 T cells in early HIV-1 infection: the preservation of the functional Vδ2 T cells responding to Fc targets, but not to IPP, in early HIV-1infection was probably due to the barely phosphoantigen-activated CD16+ Vδ2 T cell subset.
A strong and consistent relationship between viral load and the level of either CD4+ or CD8+ T cell activation has been found during early HIV infection. 28 As opposed to conventional T cells bearing αβ TCR, neither the IPP-induced nor the Fc target-induced activation of Vδ2 T cells from the early-infected group was related to the viral load or the CD4 T cell counts. Individuals who progressed rapidly after HIV-1 infection with CD4<300 in the early phase still maintained a functional Vδ2 T cell pool, in contrast to chronically infected subjects with CD4<300, who had a severe impairment of IPP responses and ADCC function. This indirectly suggests that the prolonged exposure to HIV-1 in the chronic phase may have a great impact on the overall defects of Vδ2 T cells, which is consistent with the study examining the ongoing damage to the Vδ2 T cell population caused by persistent chronic HIV-1 infection, in terms of both cell count and function. 26
Contrary to individuals in the early stage of HIV-1 infection, the capacity of Vδ2 T cells to respond to both IPP and to Fc targets had a close relationship with disease progression in chronic infection. Vδ2 T cells from chronically HIV-infected subjects with fast disease progression—as seen by low CD4 T cell counts—were compromised in their ability to respond to IPP and exert ADCC. Of importance, in the subjects with slower disease progression, despite the ongoing depletion of IPP-responsive Vδ2 T cells, a high quality of Vδ2 T cell cytotoxic response to Ab-coated targets was observed, strongly suggesting that CD16+ Vδ2 T cells might contribute to the control of HIV-1 disease progression as potent ADCC effector cells.
Studies involving HIV-infected subjects receiving HAART have shown that suppression of viral load is closely related to partial recovery of the Vδ2 T cell receptor repertoire. 29,30 However, there is scarce evidence for the Vδ2 T cell-mediated ADCC response in treated HIV-1 patients. Our study shows that treatment not only partially restored IPP-responsive Vδ2 T cells, but also significantly increased ADCC functional Vδ2 T cells in a treated cohort versus untreated HIV-infected subjects with a low CD4 T cell count. Recently, zoledronate, together with interleukin-2, has been found to amplify the CD16+ Vδ2 T cell subset and could expand Vδ2 T cells from HIV-infected individuals to become cytotoxic effectors for ADCC in vitro. 31 Here we demonstrate a restoration of Vδ2 T cell-mediated ADCC in treated patients with low viral load and immune recovery of CD4 T cells, which supports the new direction of research aimed at bolstering ADCC to improve HIV-1 treatment.
Altogether, we identified two unique Vδ2 T cell subsets discriminated by CD16 and demonstrate that HIV-1 disease progression is not only related to the magnitude of the IPP-activated response dominated by the CD16− Vδ2 T cell subset, but is also related to the capacity of CD16+ Vδ2 T cells to mediate ADCC. Although the p815 assay does not directly evaluate the capacity of HIV-specific antibodies to mediate ADCC, it provides an indirect measure of the potential capacity of the Vδ2 T cells as the ADCC effectors to clear the HIV-infected cells. Notably, HIV-1 infection has different impacts on the functional activity of the two Vδ2 T cell subsets: HIV-induced depletion of Vδ2 T cells eliminates phosphoantigen-stimulated CD16−Vδ2 T cells but not the cytotoxic, Fc target-responsive CD16+Vδ2 T cells, a phenomenon that is seen in patients with early and slow-progressed chronic HIV-1 infection but not in individuals with fast HIV-1 disease progression. As a member of the ADCC effectors, the Vδ2 T cell pool has alterations in its overall functional profile with significantly increased cytotoxicity in HIV-1 infection. These data highlight the CD16+ Vδ2 T cell subset as one with preserved ADCC activity throughout HIV-1 disease progression despite the generally damaged Vδ2 T cell pool, and warrants the selective generation of CD16+ Vδ2 T cells capable of mediating ADCC as a promising strategy for HIV vaccine and therapy research.
Footnotes
Acknowledgments
The authors thank David Pauza for his generous advice and Wilfredo F. Garcia Beltran and Christian Korner for their editorial assistance.
This work was supported by National Major Projects for Infectious Diseases Control and Prevention (2012ZX10001008, 2012ZX10004501-001-006), National Natural Science Foundation of China (81020108030, 81261120379, 31100126), SKLID Development Grant (2012SKLID103, 2011SKLID207).
Prior abstract publication: X. He et al.: Two independent functions of Vγ2Vδ2T cells discriminated by CD16 during HIV-1 infection. Retrovirology 2012;9(Suppl 2).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.