
Abstract
The mechanisms underlying B cell activation that persists during antiretroviral therapy (ART) are unknown. Toll-like receptor (TLR) signaling is a critical mediator of innate cell activation and though B cells express TLRs, few studies have investigated a role for TLR signaling in B cell activation during HIV infection. We addressed this question by assessing the activated phenotype and TLR expression/responsiveness of B cells from ART-treated HIV-infected subjects (HIVART +). We evaluated activation markers implicated in B cell-mediated T cell trans infection during HIV pathogenesis. We found no significant difference in TLR expression between B cells of HIVART + and HIV− subjects. However, B cells of HIVART + subjects exhibited heightened endogenous expression levels of IL-6 (p=0.0051), T cell cognate ligands CD40 (p=0.0475), CD54 (p=0.0229), and phosphorylated p38 (p<0.0001), a marker of TLR signaling. In vitro, B cells of HIVART + individuals were less responsive to TLR stimulation compared to B cells of HIV− subjects. The activated phenotype of in vitro TLR-stimulated B cells of HIV− subjects was similar to ex vivo B cells from HIVART + individuals. TLR2 stimulation was a potent mediator of B cell activation, whereas B cells were least responsive to TLR4 stimulation. Compared to HIV− subjects, the serum level of lipoteichoic acid (TLR2 ligand) in HIVART + subjects was significantly higher (p=0.0207), correlating positively with viral load (p=0.0127, r=0.6453). Our data suggest that during HIV infection TLR-activated B cells may exert a pathogenic role and B cells from HIVART + subjects respond to in vitro TLR stimulation, yet exhibit a TLR tolerant phenotype suggesting prior in vivo TLR stimulation.
Introduction
C
During HIV infection, B cell hyperactivation is characterized by elevated expression of activation/costimulatory markers, spontaneous cytokine expression, hypergammaglobulinemia, and B cell malignancies. 7 The mechanisms underlying B cell hyperactivation in particular and immune activation in general are poorly understood and likely multifactorial. 1 Nonetheless, during HIV infection, Toll-like receptor (TLR) signaling is identified as a key component of immune activation. 8 –12 TLRs are members of the family of pattern recognition receptors (PRR) expressed on immune cells. Their primary function is to activate innate immunity in response to foreign antigens including bacterial, fungal, and viral products and antigens. 13 The systemic prevalence of these TLR ligands responsible for mediating immune activation is associated with translocation of gut microbes and microbial products during HIV infection. 14 Recently, Funderburg et al. proposed that systemic exposure to microbial TLR agonists may drive T cell activation in chronic HIV infection. 12 Multiple studies report aberrant TLR expression and signaling in innate cells during treated and untreated HIV infection. 15 –19 Furthermore, in ART-treated subjects supplementing therapy with TLR7 antagonist is reported to reduce immune activation, 20 –22 indicating TLR signaling drives immune activation even during ART. Finally, a study by Baenziger et al. 23 –25 demonstrated that sustained exposure to TLR ligands leads to a B cell phenotype in mice, similar to that seen in HIV-infected subjects.
Human B cells like innate cells express TLRs and multiple studies demonstrate that ligation of these TLRs leads to activation and cytokine secretion, including during inflammation-associated diseases. 26 –28 Nonetheless, few studies have investigated B cell TLR expression and signaling during HIV infection, and previous studies were focused on TLR9 pathways. 29
In this comprehensive study we investigate the B cell expression/responsiveness to TLR2, TLR4, and TLR9. The properties of these TLRs have been well characterized in human B cells. 30 –32 Furthermore, we evaluated B cell expression of the proinflammatory cytokine IL-6 as well as T cell cognate ligands CD40 and CD54 (ICAM-1), as interleukin (IL)-6, CD40, and CD54 have been implicated in B cell-mediated trans infection of T cells with HIV. 30 –33 Our findings indicate that the blunted response of B cells from ART-treated individuals to further TLR stimulation suggests prior in vivo TLR stimulation. These data indicate a possible in vivo role for TLR signaling-mediated B cell activation during HIV infection that persists during ART.
Materials and Methods
Study participants
All studies were performed after an informed written research consent form was signed by each study participant. HIVART + subjects had an age range from 25 to 64 years, a viral load range of 40–1,094 copies/ml, a median of 40 copies/ml, and a CD4 count range of 103–1,236 cells/μl. HIV− subjects had an age range of 26–69 years. The study was reviewed and approved by the Institutional Review Boards of the Rush University Medical Center and Cook County Health and Hospitals System.
Isolation and TLR stimulation of PBMCs
Peripheral blood mononuclear cells (PBMCs) were isolated from whole blood using Ficoll gradient (Lympholyte Cell Separation Media, Cederlane). Live cells were enumerated by Trypan Blue exclusion. PBMCs (106) were incubated overnight either with medium alone or with 10 μg/ml CpG-B ODN2006, 2 μg/ml LPS (Invivogen), or 2 μg/ml PAM3CSK4 (Invivogen).
Immunophenotyping and intracellular cytokine staining
To phenotype B cells the following antibodies were used: CD19–ECD (Beckman Coulter), CD40–APC (BD Biosciences), CD54–PE (BD Biosciences), TLR2–Alexa Fuor 488 (BD Pharmingen), TLR4-APC (ebioscience), and TLR9-PE (BD Pharmingen). To determine intracellular cytokine expression, 4 h after stimulation with TLR ligands described above, cell cultures were supplemented with Brefeldin A (1:1,000) and incubated overnight. The cells were then fixed and permeabilized using the FIX&PERM kit (BD Biosciences, according to the manufacturer's instructions) and stained with IL-6-PE (BD Biosciences) and tumor necrosis factor (TNF)-α-PECy7 (BD Biosciences).
Gating strategy
Lymphocyte-sized cells were acquired (BD LSR II flow cytometer) based on forward-side scatter distribution and only live (determined by LIVE/DEAD Fixable Dead Cell Stain Kit, Invivogen) CD19+ cells were analyzed. Analysis was performed using Flow-jo software (Tree Star Inc.).
MAPK activation assay
The cells were stimulated overnight with the respective TLR ligands and after surface staining as described above, PBMCs were fixed with BD Cytofix buffer (Cat. No. 554655) for 10 min at 37°C and then permeabilized with the Phosflow BD Perm Buffer III (Cat. No. 558050) by incubating on ice for 30 min. Cells were then washed twice in BD Pharmingen Stain Buffer stained with PE-labeled anti-p38 MAPK (pT180/pY182, BD Biosciences) antibody and analyzed by flow cytometry.
ELISA assay for detection of TLR2-ligand, lipoteichoic acid (LTA) in serum
Serum was diluted (1:10) in ELISA coating buffer (KPL, Gaithersburg, MD) in microtiter plates and incubated overnight at 4°C. Bound protein was captured using rabbit polyclonal anti-LTA antibody (1:350, Abcam) and detected by goat polyclonal antirabbit horseradish peroxidase (HRP) (1:10,000, Abcam) antibody; optical density was measured at 450 nm.
Statistical analysis
Results are expressed as means±standard errors of the means (SEM) or as indicated. Graphpad Prism software, version 5.03 was used for all statistical analysis. The significance of differences between parameters was determined using an unpaired or paired Student's t test (with a confidence level of 95%) or a one-way ANOVA nonparametric test. The relationship between variables was calculated using the Spearman rank correlation analysis. p values of <0.05 were considered statistically significant.
Results
Constitutive elevated expression of T cell cognate ligands and spontaneous proinflammatory cytokine expression by B cells from ART-treated (HIVART +) subjects
Results from studies indicate that during HIV infection activated B cells exert pathogenic functions. 34,35 Furthermore, during ART B cell activation persists contributing to the genesis of B cell malignancies, which are highly prevalent in ART-treated HIV-infected subjects. 30 –33 Non-AIDS-defining malignancies are a major healthcare issue in the ART era; thus, understanding the mechanisms underlying B cell activation in ART-treated individuals might lead to therapeutic interventions. To address activation during ART, we assessed B cell activation markers from HIVART + individuals and found they exhibited significantly elevated expression of the immune activation markers and T cell cognate ligands CD40 (p=0.0475) and CD54 (p=0.0229; Fig. 1b). By intracellular cytokine staining (ICS), we determined that in HIVART + individuals a significantly higher percentage of B cells constitutively express IL-6 (p=0.0051); but not significantly more TNF-α (Fig. 1c). B cell expression of CD40, CD54, and proinflammatory cytokines has been implicated in the B cell trans infection of T cells with HIV. 11,15,36,37
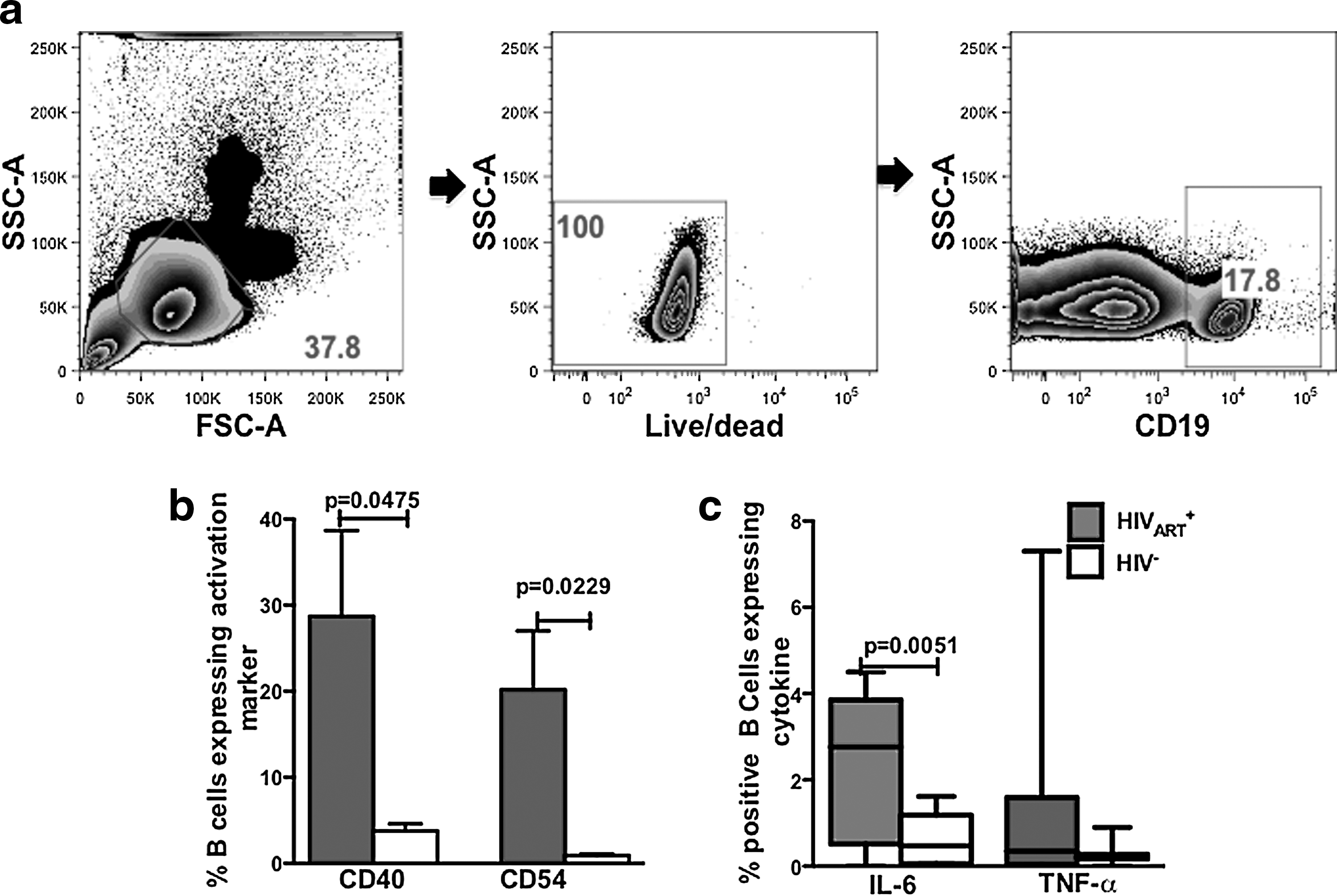
B cells from HIVART
+ subjects have a significantly higher percentage of cells constitutively expressing CD40, CD54, and interleukin (IL)-6. Peripheral blood mononuclear cells (PBMCs) (106) from HIV− and HIVART
+ subjects (n=14) were investigated by flow cytometry
B cells from HIVART + subjects are less responsive to in vitro TLR stimulation
Conflicting results have been reported regarding the ex vivo responsiveness of innate cells from HIVART + individuals to in vitro TLR stimulation. 26 –28 Similarly, contradictory reports have been published regarding the responsiveness of B cells from HIV-infected individuals to TLR9 stimulation. 11,16,17 We investigated the response of B cells from HIV-infected subjects to further TLR stimulation by exposing PBMCs from HIVART + and HIV− subjects to TLR2, 4, or 9 ligands and evaluated the upregulation of CD40, CD54, and IL-6 expression. In HIVART + subjects TLR2 ligand exposure led to a significantly increased percentage of B cells expressing CD40 (Fig. 2a, p=0.0138), while TLR9 stimulation led to an elevated but not significantly increased percentage of CD40 B cells (Fig. 2a, p=0.0736); B cells were least responsive to TLR4 ligand exposure.

B cells from HIVART
+ subjects exhibit a blunted expression of CD40, CD54, and IL-6 in response to Toll-like receptor (TLR) 2, 4, and 9 stimulation. Peripheral blood mononuclear cells (PBMCs) (106) from HIV− and HIVART
+ subjects (n=10) were stimulated overnight either with medium or TLR2 (PAM3CSK4, 2 μg/ml), TLR4 (LPS, 2 μg/ml), or TLR9 (CpG-B, 10 μg/ml) ligands and expression of

Interestingly, in HIV− subjects, TLR2, 4, and 9 ligand exposure led to a significantly increased percentage of B cells expressing CD40 (Fig. 2a, p<0001, p=0.0101 and p=0.0017, respectively). In HIVART + subjects, a significantly elevated percentage of CD54-positive B cells was observed after TLR2 stimulation (Fig. 2b, p=0.0253) and TLR9 stimulation (Fig. 2b, p=0.0029). The same trend for CD54 expression was observed after TLR stimulation of cells from HIV− subjects (Fig. 2b, TLR2, p=0.0002; TLR9, p=0.0004, respectively). In Fig. 1c, we found that compared to HIV− subjects, a significantly higher percentage of B cells from HIVART + individuals constitutively expressed higher levels of IL-6 than TNF-α. We therefore investigated B cell IL-6 expression after in vitro TLR stimulation. In HIVART + individuals, TLR2, 4, and 9 stimulation led to moderately elevated IL-6 production (Fig. 2c, left panels). However, exposure of B cells from HIV− subjects to TLR2, TLR4, and TLR9 stimulation led to significantly elevated percentages of IL-6-positive cells (Fig. 2c, right panels p=0.0361, p=0.0419, and p=0.0104, respectively). Taken together, our data (summarized in Table 1) indicate that compared to B cells from HIV− subjects, B cells from In HIVART + individuals exhibit a blunted response to in vitro TLR stimulation.
IL-6, interleukin 6.
B cells from HIVART + have significantly higher levels of TLR signaling-associated phosphorylated p38-MAPK
The blunted response of B cells from HIVART + subjects to in vitro TLR stimulation is an indication of prior in vivo stimulation during chronic HIV infection. Elevated ex vivo TLR expression on innate cells of HIV-infected individuals has been associated with persistent in vivo TLR stimulation. 16 –18,38 However, Jiang et al. 28 report a downregulation of TLR9 expression on B cells of HIV-infected individuals; therefore we assessed the expression of TLR2, 4, and 9 on ex vivo B cells from HIVART + individuals. We observed no statistically significantly differences in expression of TLR2, 4, and 9 between B cells from HIV− (n=11) and HIVART + individuals (n=12) (Fig. 3a). To determine if the constitutive activated phenotype observed in B cells from HIVART + individuals (Fig. 1) was associated with in vivo TLR stimulation, we assessed MAPK signaling. Activation of the MAPK signaling pathway, a critical regulator in the expression of proinflammatory cytokine and immune activation markers, 38 has been reported to be one of the earliest events in B cell signaling following TLR ligation. 39 We determined that compared to B cells from HIV− subjects (n=10), B cells from HIVART + individuals (n=12) exhibited significantly elevated constitutive levels of phosphorylated p38 (p<0.0001, Fig. 3b, left panel). In vitro, p38-MAPK activation has been previously shown only following TLR7 stimulation of B cells. 40 To investigate possible differential responses in p38-MAPK signaling to TLR stimulation, we exposed PBMCs from healthy controls (n=10) to TLR2, 4, or 9 ligands and by flow cytometry determined the level of p38-phosphorylation in B cells. We observed that only TLR2 exposure led to a significantly elevated expression of phosphorylated p38 (Fig. 3b, p=0.0134 right panel).

Compared to B cells from HIV− subjects, B cells from HIVART
+ subjects have no statistically significant difference in TLR2, TLR4, and TLR9 expression yet significantly elevated levels of p39-MAPK signaling.
Association between markers of HIV disease progression and B cell activation
Our data indicate that TLR2 is the most potent mediator of B cell activation in both HIV− and HIVART + individuals (Fig. 2). Biomarkers of systemic immune activation and microbial translocation have been associated with the elevated activation state of T cells and monocytes. 14 To determine the potential in vivo significance of TLR2-mediated stimulation of B cells during HIV infection, we evaluated ex vivo levels of TLR2 ligands. We determined the serum levels of LTA, an important Gram-positive TLR2 ligand known to stimulate innate cells. 41 –43 We found significantly (p=0.0207, Fig. 4a) higher levels of LTA in the serum of HIVART + individuals compared to healthy controls. The serum LTA levels positively correlated with viral load (Fig. 4b, r=0.6453, p=0.0127).

HIVART
+ subjects have higher serum levels of lipoteichoic acid (LTA, TLR2 ligand), positively correlating with viral load.
Discussion
Systemic chronic immune activation, a critical driver of HIV disease progression, persists even after ART, 44 contributing significantly to the genesis of non-AIDS-defining malignancies. In ART-treated subjects, supplementing therapy with TLR7 antagonists is reported to reduce immune activation, 20,21 indicating TLR signaling drives immune activation even during ART. Immune activation has been associated with microbial translocation 14 across the gut epithelial barrier during acute HIV infection resulting in systemic prevalence of microbes and microbial products. These microbial products, and likely HIV itself, activate innate cells via TLR signaling. 8,9,22 Accordingly, a huge body of work exists characterizing the TLR responsiveness of innate cells in treated and untreated HIV infection. 9,11,15 –18,36 Though B cells express a range of TLRs 45,46 and B cell TLR stimulation is well described in other disease settings, 24,25,45,47 only studies limited to B cell TLR9 responses have been reported in HIV infection. 26 –28
HIV infection-associated B cell activation as defined by expression of activation/costimulatory markers was identified over two decades ago 48 ; however, the underlying mechanisms remain poorly elucidated. Our data strongly suggest that during HIV infection B cells undergo activation via direct TLR stimulation. We determined that despite ART and controlled viremia, the B cells of HIVART + individuals had significantly elevated levels of activation and inflammation markers: IL-6, CD40, and CD54 (ICAM-I), which have been shown to mediate B cell trans infection of T cells with HIV. 30,31,33 This activated phenotype was associated with elevated levels of phosphorylated p38, a kinase downstream of B cell TLR signaling and master regulator of activation marker and proinflammatory cytokine expression. 38,49
Inflammation in ART-treated patients mediated by elevated serum IL-6 levels has been linked to an increased risk of mortality and morbidity. 2,3 Therefore our data suggest that during HIV infection B cells may contribute to the inflammatory milieu by secreting IL-6. This would corroborate reports of TLR-mediated B cell secretion of IL-6 in other inflammatory diseases. 50,51
Data from multiple studies suggest that HIV infection modulates TLR expression in vivo and in vitro. 11,16 –18 Jiang et al. 28 reported a reduction in TLR9 mRNA levels of B cells from HIV-infected individuals. We find that ex vivo B cells of HIVART + individuals exhibit modest but not statistically significant increased expression levels of TLR2, TLR4, and TLR9 compared to healthy controls. It is likely that ART treatment results in normalization of TLR expression levels.
Along similar lines, Hernandez et al. 16 reported that ART leads to a reduction of TLR expression levels compared to levels in treatment-naive subjects. Interestingly, in the highly relevant nonhuman primate model of HIV infection, 52 we determined that pathogenic chronically SIV-infected pigtailed macaques exhibited significantly elevated B cell TLR2 expression (data not shown) not observed in SIV-infected nonpathogenic African Green monkeys. This suggests that in vivo persistent B cell TLR2 stimulation is associated with pathogenic SIV infection.
Conflicting results have been published regarding the responsiveness of innate cells from HIV-infected individuals to further in vitro TLR stimulation. 15,17,18,37,53 Similarly, there are conflicting reports about the responsiveness of B cells from HIV-infected individuals to further TLR9 stimulation. 27,28 Since a mixed cell culture as opposed to purified cell populations more accurately reflects the in vivo situation, we exposed PBMCs from healthy controls and HIV-infected subjects to TLR2, TLR4, and TLR9 ligands to investigate the effect of HIV infection on the responsiveness to TLR stimulation. We find that in HIVART + individuals the responsiveness to further TLR stimulation was dependent on the marker of activation tested. There was significant upregulation of CD40 and CD54 but not IL-6. Overall, these B cells were less responsive to TLR agonists compared to B cells from HIV− subjects. This attenuated response by B cells of HIVART + individuals is most likely due to TLR tolerance induced by persistent in vivo TLR stimulation. Published data suggest that persistent TLR7 B cell stimulation leads to TLR7 tolerance. 54
More evidence suggesting in vivo TLR stimulation during HIV infection was provided by our finding that TLR-stimulated B cells from HIV− subjects mimic the activated phenotype seen in B cells of HIVART + individuals: elevated levels of p38 phosphorylation, upregulated expression of CD40 and CD54, and heightened IL-6 but not TNF-α (not shown) production. p38 activation has only been shown to be activated downstream of TLR7 in B cells. 40 Here we show that TLR2 as well as TLR9 stimulation activates p38 signaling, although in our hands statistically significant phospho-p38 levels were observed only after TLR2 stimulation. This observation follows the trend we observe, that in B cells of HIVART + and HIV− subjects TLR2 stimulation was a consistently potent mediator of B cell activation. This prompted us to question whether during HIV infection, there was a serum prevalence of TLR2 ligands in serum as has been reported for TLR4 and TLR9 ligands. 14,55 Indeed, we found that LTA from translocated Gram-positive bacteria can also be detected in the serum of HIV-infected individuals. It is likely that translocated Gram-positive bacteria directly stimulate B cells via TLR2 signaling or preexposure of B cells to HIV-encoded TLR7 ligands, 56 which might enhance B cells responses to TLR2 ligands as has been reported to be the case for monocytes. 57
Taken together, our data provide compelling evidence that during HIV infection TLR signaling contributes to B cell activation. It is possible that TLR-activated B cells exert a pathogenic role and during ART, persistently TLR-stimulated B cells might contribute to the inflammatory milieu and the genesis of HIV-associated B cell malignancies. Interestingly, dysregulated TLR2 expression/signaling has been reported in certain B cell lymphomas. 58,59 Investigations elucidating TLR-mediated activation in the genesis of non-AIDS-defining B cell malignancies could lead to the development of biomarkers and therapeutic interventions.
Footnotes
Acknowledgments
This study was supported by funding from the National Institutes of Health–Developmental Center for AIDS Research (P30 AI082151-01 and P01AI076174-01A1). We thank Jeffrey Martinson of the Rush University Medical Center Flow Cytometry Facility. We also thank Dr. Alison Finnegan, Jeffrey Martinson, Jill Plants, Sonya Rigielski, and Kirstin Bush of Rush University Medical Center for critically reading the manuscript.
Author Disclosure Statement
No competing financial interests exist.