
Abstract
Liver disease is now a major cause of morbidity and mortality among persons infected with the human immunodeficiency virus (HIV). An increasing body of evidence suggests that HIV infection is associated with exacerbated liver fibrosis and that HIV has the ability to infect several hepatic cell types. Despite the recognized existence of genetically distinct subpopulations of HIV in the central nervous system and genital tract, viral diversity and compartmentalization in the liver have not been explored extensively. Therefore, phylogenetic analysis was performed on full-length env and nef sequences for four patients. Distinct clustering of viral variants was observed for all patients in both areas of the genome. Statistical evidence of HIV compartmentalization in the liver was demonstrated in 85.4% of comparisons. Signature sequence analysis identified several liver-specific amino acids in all patients. Thus, the current study demonstrates statistically significant evidence for HIV compartmentalization in the liver. Additionally, these data suggest that the hepatic microenvironment harbors unique selective pressures that drive viral adaptation.
Introduction
C
Emerging data suggest that HIV has the ability to infect cells of the liver, including endothelial sinusoidal cells, stellate cells, Kupffer cells, and hepatocytes. 11 –16 Collectively, these data indicate that HIV is present in several intrahost microenvironments, each harboring its own unique selective pressures, thus contributing to the extensive sequence diversity of HIV. However, to our knowledge, HIV diversity in the liver has been explored in only one previous study. 17 Therefore, the current analysis utilized complementary phylogenetic and statistical approaches to examine intrapatient HIV variability and compartmentalization in the liver.
Materials and Methods
All sequence data from patients HIVAms198, AM, AZ, and DY were previously published without phylogenetic or statistical evaluation of viral compartmentalization.
18,19
All tissue samples were obtained postmortem from patients who died of AIDS-related causes. For the current analysis of viral compartmentalization, alignments were performed using the neighbor-joining (NJ) approach as implemented in Clustal X2.
20
The statistical robustness and reliability of the branching order within each phylogenetic tree were confirmed by bootstrap analysis using 1,000 replicates.
21
The Viral Epidemiology Signature Pattern Analysis (VESPA) program was used to determine the frequency of each amino acid in liver-derived viral variants versus viral variants from other tissues for each individual (
Results
Phylogenetic analyses were performed on full-length HIV gp120 and nef sequences for patients AM, AZ, and DY, and on V3 loop sequences for patient HIVAms198. A single intrapatient neighbor-joining tree was compiled including sequence data from all tissues for subject HIVAms198 (Fig. 1A). Interestingly, liver-derived sequences formed a single monophyletic cluster that contained no non-liver-derived sequences. Peripheral blood mononuclear cell (PBMC)-derived sequences were interspersed throughout the phylogenetic tree but were not closely related to the liver sequences, suggesting that liver-derived sequences were not the result of peripheral lymphocyte infiltration. Neighbor-joining phylogenetic trees including sequence data from all tissues were created for HIV gp120 and nef sequences for patients AZ and DY (Figs. 1B and C and 3A and B). Due to tree size constraints, phylogenetic trees for HIV gp120 and nef sequences from patient AM are shown as paired comparisons of the liver and one other tissue (Figs. 2 and 4). Distinct clustering of liver variants was observed in patients AM, AZ, and DY as well. Evidence of viral compartmentalization was supported further by Mantel's tests results, with a p-value<0.05 for 41 of 48 comparisons involving the liver (85.4%) (Table 1).
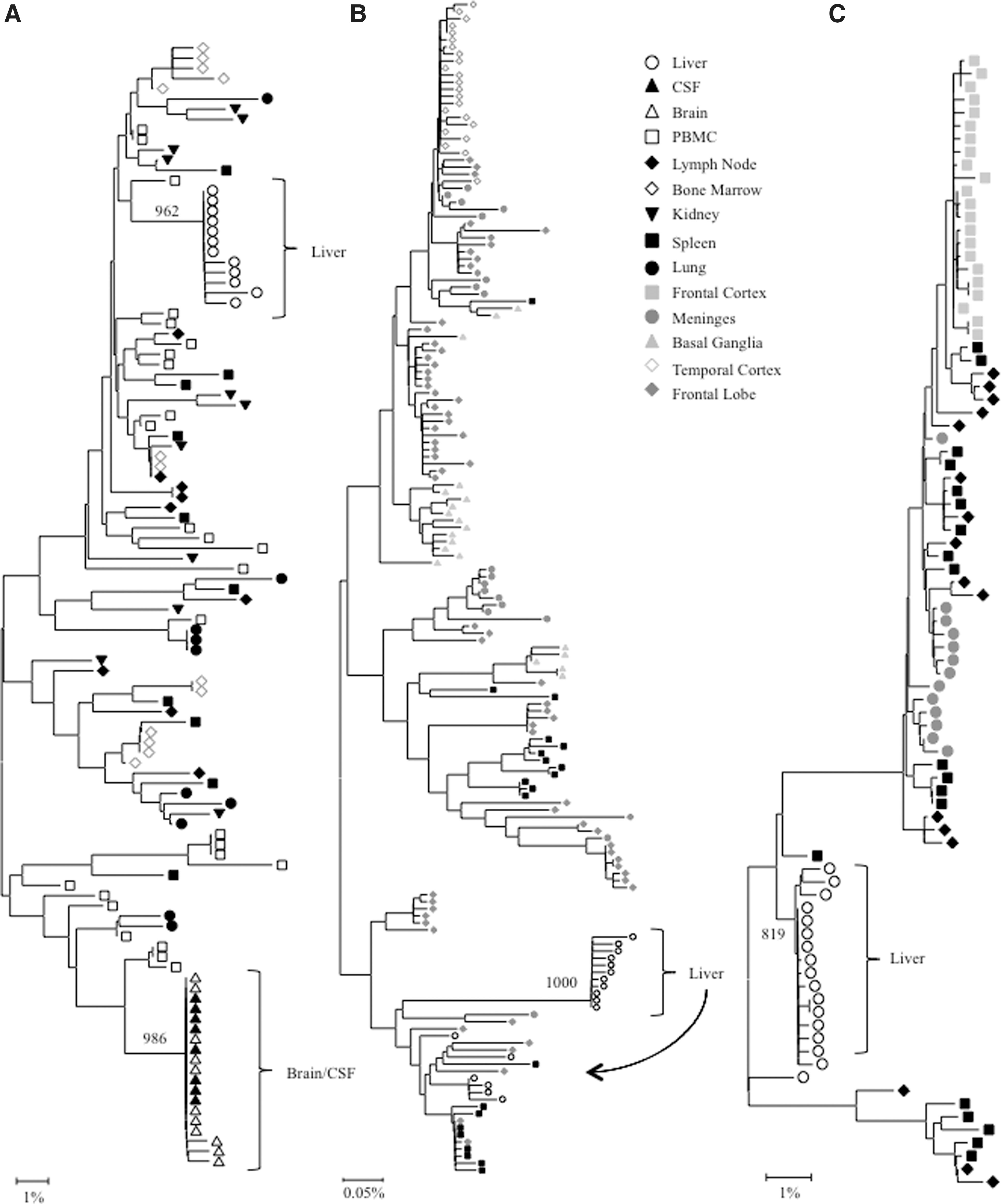
Intrapatient phylogenetic trees of
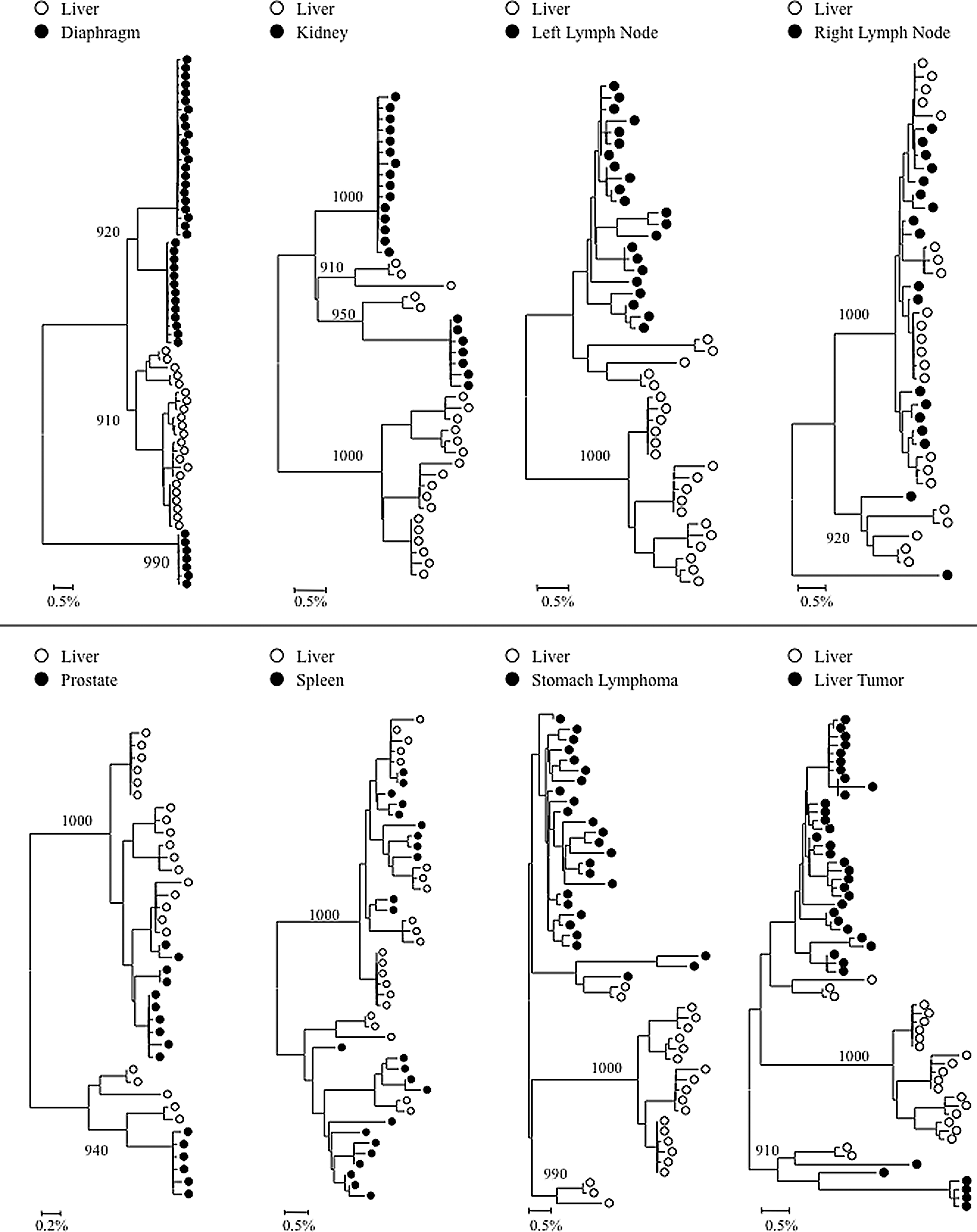
Intrapatient phylogenetic trees of gp120 variants from patient AM. Shown in the lower left corner is a bar depicting the percent genetic distance for each tree. Only relevant bootstrap values greater than 700 out of 1,000 are shown. Sequence data were originally reported in Lamers et al. 18
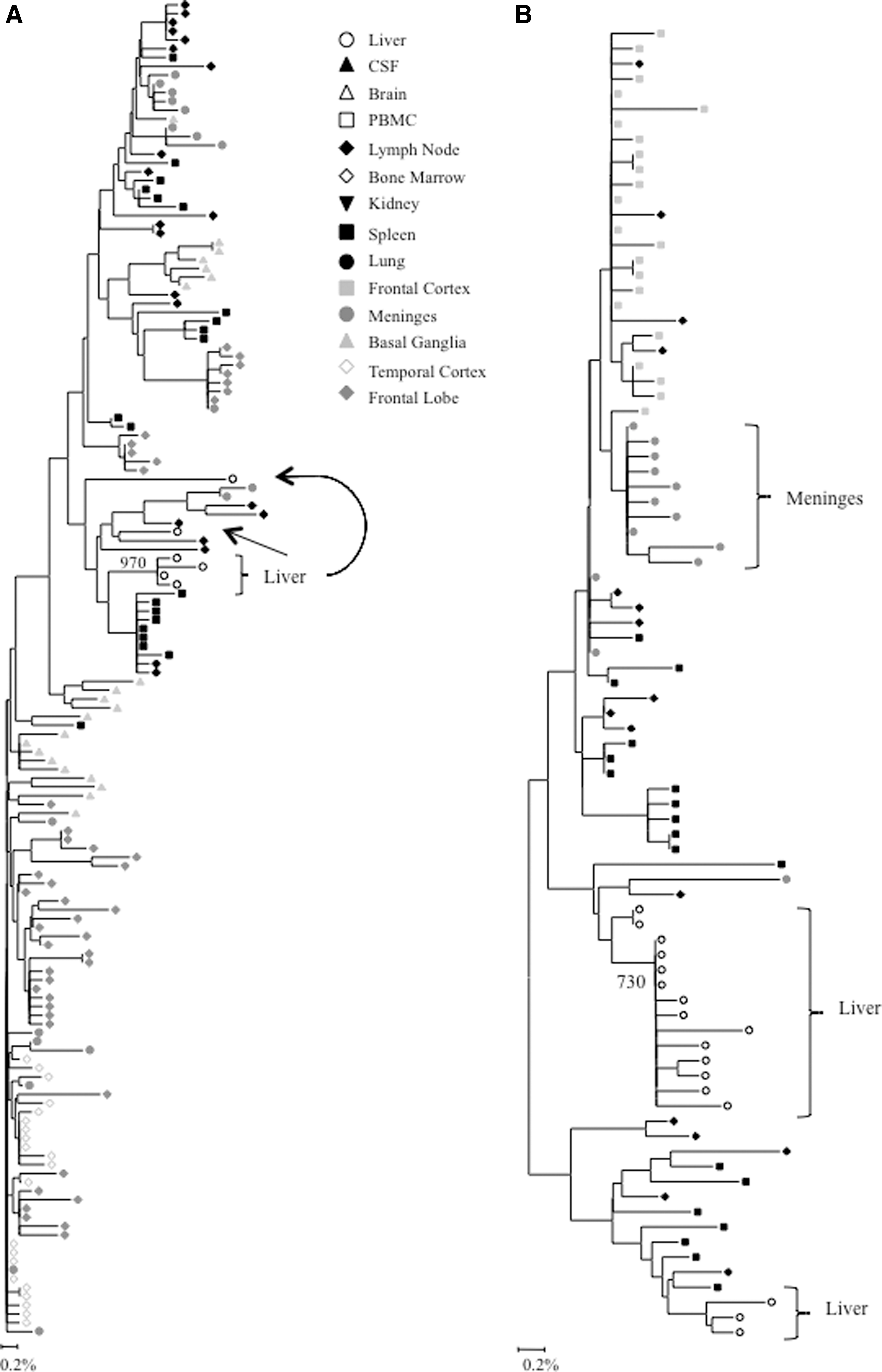
Intrapatient phylogenetic trees of

Intrapatient phylogenetic trees of nef variants from patient AM. Shown in the lower left corner is a bar depicting the percent genetic distance for each tree. Only relevant bootstrap values greater than 700 out of 1,000 are shown. Sequence data were originally reported in Lamers et al. 18
Sequence data for patient HIVams198 were originally reported by Van't Wout et al. 19 Sequence data for patients AM, AZ, and DY were originally reported in Lamers et al. 18
CSF, cerebrospinal fluid; PBMC, primary blood mononuclear cells; GM, gray matter; WM, white matter; NS, not significant; dash, no data available; p-values<0.05 were considered statistically significant.
Analysis of V3 loop sequences
Phylogenetic analysis demonstrated that V3 loop sequences from the brain and cerebral spinal fluid (CSF) exhibited distinct phylogenetic clustering. Likewise, V3 loop variants from the liver formed a distinct monophyletic group supported by a high bootstrap value (962 out of 1,000). No further compartmentalization was observed for any V3 loop sequences from the kidney, lung, lymph node, bone marrow, spleen, and PBMCs (Fig. 1A). Thus, HIV quasispecies compartmentalization was observed in the liver and brain/CSF only for patient HIVAms198. Evidence for significant V3 loop compartmentalization was further supported by Mantel's test, in which all pairwise genetic distance comparisons of V3 loop sequences involving the liver had a p-value<0.05 (Table 1). Signature sequence analysis identified specific amino acid differences between V3 loop variants from the liver and all other tissues (Supplementary Table S1; Supplementary Data are available online at
Analysis of gp120 sequences
Intrapatient phylogenetic trees including gp120 sequences from all available tissues were created for patients DY and AZ (Fig. 1B and C). Both patients exhibited distinct phylogenetic clustering of gp120 variants from the liver supported by high bootstrap values (1,000 for DY and 819 for AZ). Similar clustering was observed with temporal cortex variants from patient DY and frontal cortex variants from patient AZ. Phylogenetic trees containing paired comparisons involving the liver were created for patient AM. Viral variants from the liver and diaphragm were particularly distinct from one another, as complete separation between these tissues was observed (Fig. 2). All pairings, except liver–right lymph node and liver–spleen showed evidence of compartmentalization supported by high bootstrap values (Fig. 2). Despite the lack of visible phylogenetic separation between the liver and spleen variants from patient AM, Mantel's test comparing these two tissues was statistically significant (p=0.0071) (Table 1). In the analysis of gp120 sequences, Mantel's test confirmed significant evidence of compartmentalization for 17 of 20 comparisons involving the liver (85%) (Table 1). Signature sequence analysis highlighted multiple amino acid differences in HIV gp120 from the liver versus all other tissues for patient DY (range=21–41, mean=30.3), AZ (range=42–46, mean=43.3), and AM (range=2–50, mean=29.3) (Supplementary Tables S2–4). Liver sequence divergence was most apparent in patient AZ, as 42 of 48 total residue differences between liver-derived variants and variants derived from other tissues were found in all three paired comparisons. The liver–diaphragm comparison from patient AM had 50 amino acid substitutions, the highest of all paired comparisons (Supplementary Table S4). Several amino acid differences between gp120 variants from the liver and other tissues occurred in functional areas, including the LDI/LDV tripeptide in the V2 loop, several CD4 contact residues and coreceptor binding sites, the GPGR tip of the V3 loop, the CD4 binding loop, a lectin DC-SIGN binding site, monoclonal antibody glycosylation binding sites, and gp120 contact residues with gp41. 27 –30
Analysis of nef sequences
Complete or near complete separation of nef variants from the liver was observed in phylogenetic analyses of sequences from patients DY and AZ (Fig. 3). The statistical robustness of liver variant branching order was supported by high bootstrap values (970 for DY and 730 for AM). Distinct phylogenetic clustering was also demonstrated by nef variants from the meninges of patient AZ. No further phylogenetic evidence of compartmentalization was observed for any other tissue evaluated from patients DY and AZ (Fig. 3). Neighbor-joining trees were completed for paired comparisons involving the liver for patient AM (Fig. 4). Complete or near complete separation of viral variants was observed for liver–diaphragm, liver–kidney, liver–prostate, and liver–liver tumor comparisons. Although there was no visible phylogenetic separation of variants between the liver and left lymph node variants from patient AM, Mantel's test suggested that sequences from the liver shared more genetic identity with each other than with sequences from the left lymph node (p=0.0108) (Table 1). Furthermore, Mantel's test provided statistically significant evidence of viral compartmentalization for 16 of 20 HIV nef comparisons involving the liver (80 %) (Table 1). Signature sequence analysis of nef identified multiple amino acid substitutions between liver-derived variants and variants from all other tissues for patients DY (range=2–5, mean=3.1), AZ (range=1–2, mean=1.5), and AM (range=1–8, mean=2) (Supplementary Table S5A–C). The two comparisons with the highest frequency of amino acid differences were the liver–diaphragm comparison from patient AM (eight substitutions) and the liver–frontal lobe white matter comparison from patient DY (five substitutions). Several residue changes occurred in the nef core domain structure and the conserved SH3 binding pocket. 31
Discussion
A very limited number of studies have examined HIV variability in the liver. Van't Wout et al. performed a phylogenetic analysis on HIV proviral DNA from multiple tissues, including the liver, of one patient who died of AIDS-related causes and observed that HIV variants from the liver clearly clustered separately from variants of all other tissues analyzed. As well, there was no phylogenetic clustering observed between PBMC-derived and liver-derived HIV sequences, suggesting that HIV compartmentalization in the liver was not due to infiltrating lymphocytes. Despite this, the authors concluded that the monophyletic grouping of liver variants was the result of low proviral copy numbers, as all liver sequences were generated from a single PCR. 19 Only one other study to date has predicted liver-specific amino acid variations in env. 17 However, that study included only partial env sequences and, therefore, was unable to identify liver-specific signature sequences outside of the region amplified. Thus, in the current study, we sought to confirm these findings in an independent dataset, analyze the entire env gene, and examine compartmentalization of HIV nef sequences in the liver. Our results demonstrate that phylogenetically distinct HIV variants from the liver exist in three other patients who have died of AIDS-related causes. These findings were further supported statistically by Mantel's test. Collectively, these data suggest that viral compartmentalization may occur more frequently in the liver than previously recognized.
Signature sequence analysis identified specific amino acids in both gp120 and nef that were unique to the liver. While a bioinformatics approach cannot specifically determine if the predicted amino acid variations are relevant in the context of a replicating virus, several of the predicted amino acid variations were identified only in liver-derived HIV sequences, thus strengthening the body of evidence that suggests viral compartmentalization in the liver. Importantly, many amino acid substitutions were found in conserved areas of gp120. This suggests that they did not simply occur as a result of random sequence variability; rather, there may be unique selective pressures acting on HIV in the liver that are not found elsewhere in the body. For example, CD4-independent HIV interaction with cells is known to occur through gp120 binding of C-type lectin receptors, such as the mannose receptor or the DC-SIGN receptor on macrophages and dendritic cells. 32,33 Indeed, even minor mutations in gp120 can drastically alter the requirement for entry receptors and cell tropism. Such mutations may be the result of strong selective pressures on HIV resulting in liver-specific viral variants.
Compartmentalization can also occur without adaptive mutation, when the selective pressures in separate organs differ such that the fitness of a particular preexisting viral subpopulation is greater in one organ than another. The predicted amino acid variations discovered in this study may have arisen from adaptive mutations or selective pressures on an existent viral subpopulation, as both mechanisms can result in viral compartmentalization. The lack of in vitro analysis in the current study does not allow for the determination of specific cell types that may support HIV replication in the liver. However, compelling evidence indicates that HIV has the ability to infect endothelial sinusoidal cells, stellate cells, Kupffer cells, and hepatocytes. 11 –16 Studies also demonstrate that the gp120 antigen may engage apoptotic pathways without direct viral infection. 34 Both direct infection of liver cells and antigen activation of liver cells may contribute to viral compartmentalization in the liver. Future in vitro studies are warranted to determine the exact mechanisms by which HIV interacts with specific liver cell types and to confirm the role of organ-specific signature sequences.
Recent data suggest a role for HIV in liver disease. 35,36 This study provides evidence of HIV compartmentalization in the liver and identifies signature amino acids that may characterize liver-specific quasispecies. These data strongly suggest that HIV variants differ in their ability to infect various cell types. Furthermore, HIV mutation may be required for attachment, entry, and replication within a given cell type. Thus, these investigations may provide additional insight into the immune response and selective pressures unique to the hepatic microenvironment.
There are several limitations to the current analysis including the small number of patients, the lack of sequence data outside the env and nef genes, and limited information of whether other comorbid conditions may impact HIV diversity and/or compartmentalization. As noted in our previous study, 17 cross-sectional analyses do not permit a detailed examination of liver-specific HIV variants over time. Similarly, the cloning strategy utilized—while robust–may not have amplified all minor variants present in a given tissue/cell type. Thus, additional studies are now warranted in larger patient populations at varying stages of liver disease progression. It is recommended that future studies concerning HIV compartmentalization in the liver compare viral variants from (1) HIV monoinfected individuals with no history of liver complication, (2) HIV monoinfected individuals with liver disease, and (3) HIV/HCV coinfected individuals. However, it should be noted that due to the rapid adaptation of HIV to its host, organ-specific (including liver-specific) signature sequences may differ between patients. The phenotypic implications of liver-specific HIV mutations should be further examined, as they may aid in the development of liver-specific treatment modalities.
Footnotes
Acknowledgments
The authors would like to thank Ms. Eleanor Powell for her review of this manuscript and the University Honors Program for funding this summer undergraduate research project and for facilitating interactions between College of Medicine faculty and undergraduate students.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.