
Abstract
Coreceptor switch from CCR5 to CXCR4 is associated with HIV disease progression. To document the evolution of coreceptor tropism during pregnancy, a longitudinal study of envelope gene sequences was performed in a group of pregnant women infected with HIV-1 of clade B (n=10) or non-B (n=9). Polymerase chain reaction (PCR) amplification of the V1-V3 region was performed on plasma viral RNA, followed by cloning and sequencing. Using geno2pheno and PSSMX4R5, the presence of X4 variants was predicted in nine of 19 subjects (X4 subjects) independent of HIV-1 clade. Six of nine X4 subjects exhibited CD4+ T cell counts <200 cells/mm3, and the presence of X4-capable virus was confirmed using a recombinant phenotypic assay in four of seven cases where testing was successful. In five of nine X4 subjects, a statistically significant decline in the geno2pheno false-positive rate was observed during the course of pregnancy, invariably accompanied by progressive increases in the PSSMX4R5 score, the net charge of V3, and the relative representation of X4 sequences. Evolution toward X4 tropism was also echoed in the primary structure of V2, as an accumulation of substitutions associated with CXCR4 tropism was seen in X4 subjects. Results from these experiments provide the first evidence of the ongoing evolution of coreceptor utilization from CCR5 to CXCR4 during pregnancy in a significant fraction of HIV-infected women. These results inform changes in host–pathogen interactions that lead to a directional shaping of viral populations and viral tropism during pregnancy, and provide insights into the biology of HIV transmission from mother to child.
Introduction
HIV-1
Long-term HIV-1 infection and accelerated clinical progression toward AIDS are associated with a shift in coreceptor usage from mainly R5 to R5X4 and X4 viruses. 17,18 Approximately 50% of subjects infected with HIV-1 subtype B show evidence of coreceptor switching, a varying incidence of which was observed in infections involving other HIV-1 clades. 1,19,20 Studies in simian-human immunodeficiency virus (SHIV)-infected rhesus macaques have shown that host HIV-specific immune responses exert an important influence on viral tropism and coreceptor usage, and that the loss of HIV-specific CD8+ T cells leads to the emergence of X4 viral variants. 21 During pregnancy, the establishment and maintenance of fetomaternal tolerance are associated with significant modulations of maternal immunity, including increased synthesis of immunoglobulins and inhibition of cell-mediated immune responses. 22 –24 Tscherning-Casper et al. reported broadening of coreceptor usage during pregnancy from CCR5 to dual CCR5 and CXCR6, an alternate coreceptor that can mediate HIV-1 entry into target cells, at least in vitro. 25 Yet the timing and manner in which the env gene and CXCR4 coreceptor tropism evolve during pregnancy remain poorly understood, in spite of the fact that these phenomena might be particularly relevant with regard to HIV disease progression and/or transmission of HIV-1 from mother to child. 26,27
In the present study, we examined evolution of the sequence of the HIV-1 env gene and of determinants associated with coreceptor tropism in a group of pregnant women infected with clade B or non-B HIV-1, and provide first evidence of coreceptor switching from CCR5 to CXCR4 during pregnancy. These results inform the interplay between the evolution of env sequences and coreceptor tropism in the context of pregnancy.
Materials and Methods
Study subjects
Study subjects were participants to the Centre maternel et infantile sur le SIDA mother-child cohort (CHU Sainte-Justine, Montreal, Canada). Nineteen pregnant HIV-infected women were enrolled in the study, of whom five were followed during two consecutive pregnancies (24 pregnancies total). Inclusion criteria were based on (1) availability of at least one serum sample per trimester of pregnancy and (2) HIV-1 viral load above the lower limit of detection. First, second, and third trimesters of pregnancy were defined as 1–13, 14–28, and 29–41 weeks of gestation, respectively. Gestational age and estimated date of fertilization were estimated based on the first day of the last menstrual period (LMP) or from fetal biometry. HIV-1 viral load was measured using the Versant HIV-1 RNA 3.0 assay (Bayer, Pittsburgh, PA), with a lower limit of detection of 1.70 log10 HIV-1 RNA copies/ml plasma. CD4+ and CD8+ T cell counts were measured by flow cytometry. HIV genotyping was based on sequencing of the HIV-1 pol and env genes. 28 Informed consent was obtained from all study participants. This research protocol was conducted according to the guidelines of the Ethics Review Board of CHU Sainte-Justine, Montreal, Canada.
Amplification and sequencing of the env gene
HIV-1 viral RNA was extracted from frozen plasma samples using the QIAamp viral extraction mini kit (Qiagen, Mississauga, Ontario, Canada) and a 945 nucleotides DNA fragment spanning the Env V1-V3 region (positions 6430–7374) was amplified as previously described. 29 Reverse transcription polymerase chain reaction (RT-PCR) was performed using primers E00 and ES8B, followed by nested PCR with primers E20 and E115. 29 PCR products were cloned into TOPO TA (Invitrogen, Mississauga, Ontario, Canada). A mean of 18 plasmid clones per time point were randomly selected for unidirectional DNA sequencing (ABI 3730xl, Applied Biosystems, Concord, Ontario, Canada).
Sequence analysis
Sequence alignment was carried out using the Clustal W tool embedded in MEGA version 4. 30 Sequences were manually edited and codon aligned with HMMER v.2.32. 31 Viral tropism was predicted for each individual clone based on the amino acid sequence of the V3 loop using (1) the geno2pheno algorithm with a false-positive rate (FPR) of 5.75% (G2P5.75) 32 and (2) PSSMX4R5 where sequences with scores ≤−6.96 were considered R5, sequences with scores ≥−2.88 were considered X4, and the tropism of sequences with scores in between these values was assigned based on whether they possessed basic amino acid residues at either position 11 or position 25. 33 Numbers of potential N-linked glycosylation sites (PNGS) were calculated using N-GlycoSite 34 and corrected manually. Subjects in whom X4 sequences were detected using either G2P5.75 or PSSMX4R5 were termed “X4 subjects,” and subjects in whom only R5 sequences were detected were termed “R5 subjects.” The net charges of V2 and V3 were calculated as the number of positively charged amino acid residues (R, K, H) minus the number of negatively charged (D, E) amino acid residues.
To provide an indicator of the proportion of X4-related mutations in a V2 sequence, a position-specific scoring matrix was assembled based on data published by Hoffman et al.
4
and Thielen et al.
6
These two studies have identified substitutions in V2 that occurred at different frequencies in X4 and R5 sequences. Based on these frequencies, a likelihood index for V2 was computed that is an indicator of the relative abundance of X4-associated substitutions within a given V2 sequence. Briefly, log likelihood ratios [ln(fX4/fR5)] were calculated for each of the 13 most relevant amino acid positions in V2 (i.e., 161, 164–166, 169, 173, 177, 182, 184, 190–192, and 195). Substitution frequencies are listed in Supplementary Table S1 (Supplementary Data are available online at
Recombinant phenotypic assay
When the HIV-1 viral load was >1,000 RNA copies per ml plasma, the tropism of HIV-1 isolates was determined using an enhanced version of the Trofile coreceptor tropism assay (Monogram Biosciences, South San Francisco, CA). 35,36 The enhanced Trofile assay, which is now used exclusively for patient tropism testing, has 100% sensitivity to detect X4 variants constituting as little as 0.3% of the total population. The Trofile DNA coreceptor tropism assay (Monogram) was used when the HIV-1 viral load was <1,000 RNA copies per ml plasma. 37 A recombinant phenotypic assay was performed only when X4-capable sequences were initially predicted using G2P5.75 or PSSMX4R5.
Statistical analysis
Data were expressed as median and interquartile range (IQR). The normality of data distribution was tested using the Kolmogorov–Smirnov test. Where data were normally distributed, Student's t test and one-way ANOVA with posttest for linear trend were used to test the significance of between-groups comparisons. Where data distribution deviated from normal, the Mann–Whitney U test and the Kruskal–Wallis test with Dunn's multiple comparison test were used. Relationships between variables were tested using Spearman's rank correlation test. p values<0.05 were considered statistically significant. Analyses were performed using GraphPad Prism 4 (GraphPad Software, San Diego, CA).
Results
Clinical characteristics of study subjects
One plasma sample per trimester was analyzed except in the case of subjects TV1021a and TV1263, where only sequences from the first trimester could be amplified, and subjects TV479b and TV481a, where the first sample was taken before pregnancy (not included in the longitudinal analyses of sequence evolution). The median age at initiation of pregnancy was 27 years (IQR=25–32). Ten of 19 subjects (52.6%) were infected with HIV-1 of clade B, six (31.6%) with clade C, one (5.26%) with clade D, one with clade A1, and one with CRF13_cpx (Tables 1 and 2). The median CD4 and CD8 counts at the time point closest to initiation of pregnancy were 260 (IQR=120–360) and 730 (IQR=480–855) cells/mm3, respectively. The median viral load at the time point closest to initiation of pregnancy was 4.27 log10 RNA copies/ml (IQR=3.81–4.58).
HIV-1 subtype, gestational age, absolute CD4+ and CD8+ T cell counts, HIV-1 viral load, fraction of CXCR4-capable sequences, and tropism were determined as described in Materials and Methods. a and b denote the first and second consecutive pregnancies in a given subject.
Asterisks denote the lower limit of detection of the viral load detection method.
ARV, antiretroviral; AZT, zidovudine; 3TC, lamivudine; d4T, stavudine; ddI, didanosine; ABC, abacavir; NVP, nevirapine; NFV, nelfinavir; RTV, ritonavir; LPV/r, ritonavir-boosted lopinavir; R5, CCR5 tropism; DM, dual-mixed tropism; —, not treated; nd, not determined; nt, not tested.
HIV-1 subtype, gestational age, absolute CD4+ and CD8+ T cell counts, and HIV-1 viral load were determined as described in Materials and Methods. a and b denote the first and second consecutive pregnancies in a given subject.
Asterisks denote the lower limit of detection of the viral load detection method.
ARV, antiretroviral; AZT, zidovudine; 3TC, lamivudine; ddI, didanosine; ABC, abacavir; IDV, indinavir; NFV, nelfinavir; EFV, efavirenz; RTV, ritonavir; LPV/r, ritonavir-boosted lopinavir; R5, CCR5 tropism; —, not treated; nd, not determined.
Five subjects were followed during two consecutive pregnancies, for a total of 24 pregnancies examined. In the case of consecutive pregnancies, the median interpregnancy interval was 1.5 years (IQR=0.88–4.21). In seven of 24 pregnancies, antiretroviral (ARV) treatment was introduced at a mean of 6.1 years before initiation of pregnancy (range=1.1–13.0 years). In 15 of 24 pregnancies (62.5%), ARV treatment was initiated between week 8 and week 20. Women were treated with protease inhibitor-based therapy in 16 of 24 (66.6%) pregnancies. None of the subjects was treated with HIV-1 entry inhibitors and subject TV1267 refused treatment. HIV-1 viral load values were normally distributed (p>0.0706; Kolmogorov–Smirnov test) and decreased significantly during the course of pregnancy (p<0.0001; one-way ANOVA with posttest for linear trend), consistent with the introduction or intensification of ARV treatment. CD4 and CD8 counts were also normally distributed (p>0.0915; Kolmogorov–Smirnov test), but no significant changes were observed over the course of pregnancy (p=0.3851 and p=0.7839, respectively; one-way ANOVA with posttest for linear trend) (data not shown). No cases of MTCT were reported in any of the study participants.
Determination of coreceptor tropism
A total of 1,240 env gene sequences were obtained and were subjected to the G2P5.75 and PSSMX4R5. Variable proportions of X4 sequences, defined as sequences with a G2P FPR≤5.75%, were observed in samples obtained from nine of 19 study subjects (47.4%), for a total of 11 pregnancies independent of HIV-1 clade (Table 1). These nine women were termed “X4 subjects,” whereas the 10 others were termed “R5 subjects” (Table 2). The presence of X4 sequences was supported by PSSMX4R5 in six of these nine X4 subjects, though this algorithm did not predict X4 sequences in three others (i.e., TV669, TV1217, and TV479). All three discordant subjects were infected with HIV-1 of clade B. In the case of patient TV669, V3 sequences that were obtained were 34 amino acid residues in length instead of 35, which is known to be associated with a higher likelihood of discordant tropism prediction as a result of incorrect alignment. 38 X4 sequences in subject TV1217 had a PSSMX4R5 score of −7.97 (lower than the PSSMX4R5 cut-off), 33 but exhibited G/R amino acid residues at positions 11/25 (data not shown), strongly suggestive that they were in fact X4 sequences. In contrast, PSSMX4R5 scores of X4 sequences from TV479 were lower than the cut-off and exhibited S/E or G/E residues at positions 11/25. Finally, the presence of X4-capable virus was confirmed using a recombinant phenotypic assay in four of nine X4 subjects (four of seven subjects for whom recombinant phenotypic assay was successfully performed) (TV643, TV995, TV1151, and TV1217) (Table 1).
In the case of subject TV1261, no particular feature of V3 would seem to explain the discordant results between G2P5.75, PSSMX4R5, and the Trofile assay. Such discordant instances were previously reported in the literature, 39,40 though no clear-cut explanation was provided for these observations. In the present study, at least three separate amplification reactions were performed for each serum sample/time point. Such replicate amplification and population sequencing were shown in previous reports to enhance the sensitivity of the genotyping tropism tests, 40,41 thus providing a possible reason for the discordant results obtained for TV1261 when using algorithms versus Trofile.
Overall, X4 subjects exhibited significantly lower median CD4+ T cell counts than R5 subjects during the course of pregnancy (p=0.0243; Student's t test) (Fig. 1A), with six of nine X4 subjects exhibiting median CD4 counts ≤200 cells/mm3 (Table 1), consistent with functional immunodeficiency. In addition, R5 subjects showed significantly greater CD4+ T cell recovery following initiation of therapy than X4 subjects (p=0.05; Student's t test) (Fig. 1B). Furthermore, a higher median HIV-1 viral load was seen in X4 subjects as compared to R5 subjects (p=0.0185; Mann–Whitney U test) (Fig. 1C). Both groups responded equally well to ARV therapy, as the decrease in HIV-1 viral load was not significantly different between the two groups (p=0.2817; Student's t test) (data not shown). CD8 counts, maternal age, and the composition of ARV regimens were not significantly associated with the presence of X4 sequences (data not shown). Finally, no statistically significant difference was found in the time from diagnosis between the two groups (p=0.396; Mann–Whitney U test) (data not shown).
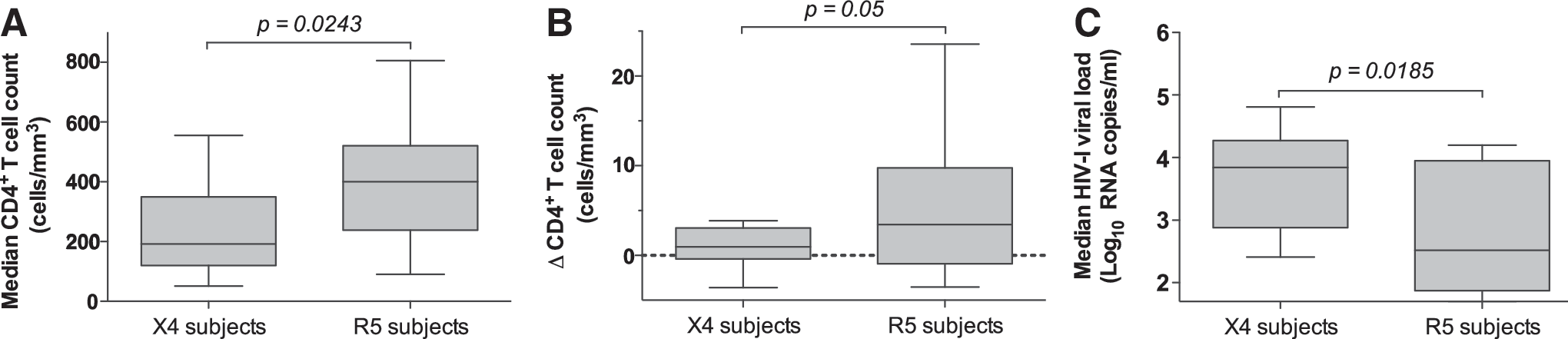
Clinical parameters in X4 and R5 subjects. CD4+ T cell counts
Evolution of V3 sequences during pregnancy
To examine the evolution of V3 sequences during pregnancy, the longitudinal changes in terms of G2P FPR, PSSMX4R5 scores, the net charge of V3, and the relative proportion of predicted X4-capable sequences were analyzed based on sets of viral sequences obtained using PCR, cloning, and sequencing at different time points during the course of pregnancy. In this analysis, statistically significant decreases in G2P FPR, statistically significant increases in PSSMX4R5 scores, and statistically significant increases in the net charge of V3 between the first and third trimesters were considered as evidence of evolution toward CXCR4 tropism. The progressive evolution of these three parameters was readily observed in X4 subjects, but not in R5 subjects (Fig. 2). Of 11 pregnancies analyzed among the nine X4 subjects, statistically significant decreases in G2P FPR were observed in five cases over the course of pregnancy (Fig. 2A), along with statistically significant increases in PSSMX4R5 scores (four of five cases) (Fig. 2B) and statistically significant increases in the net charge of V3 (five of five cases) (p≤0.05; Mann–Whitney U test) (Fig. 2C). Of these five subjects, four also showed a progressive increase in the relative proportion of predicted X4-capable sequences through time (i.e., TV643, TV715, TV995, and TV1261; Table 1).
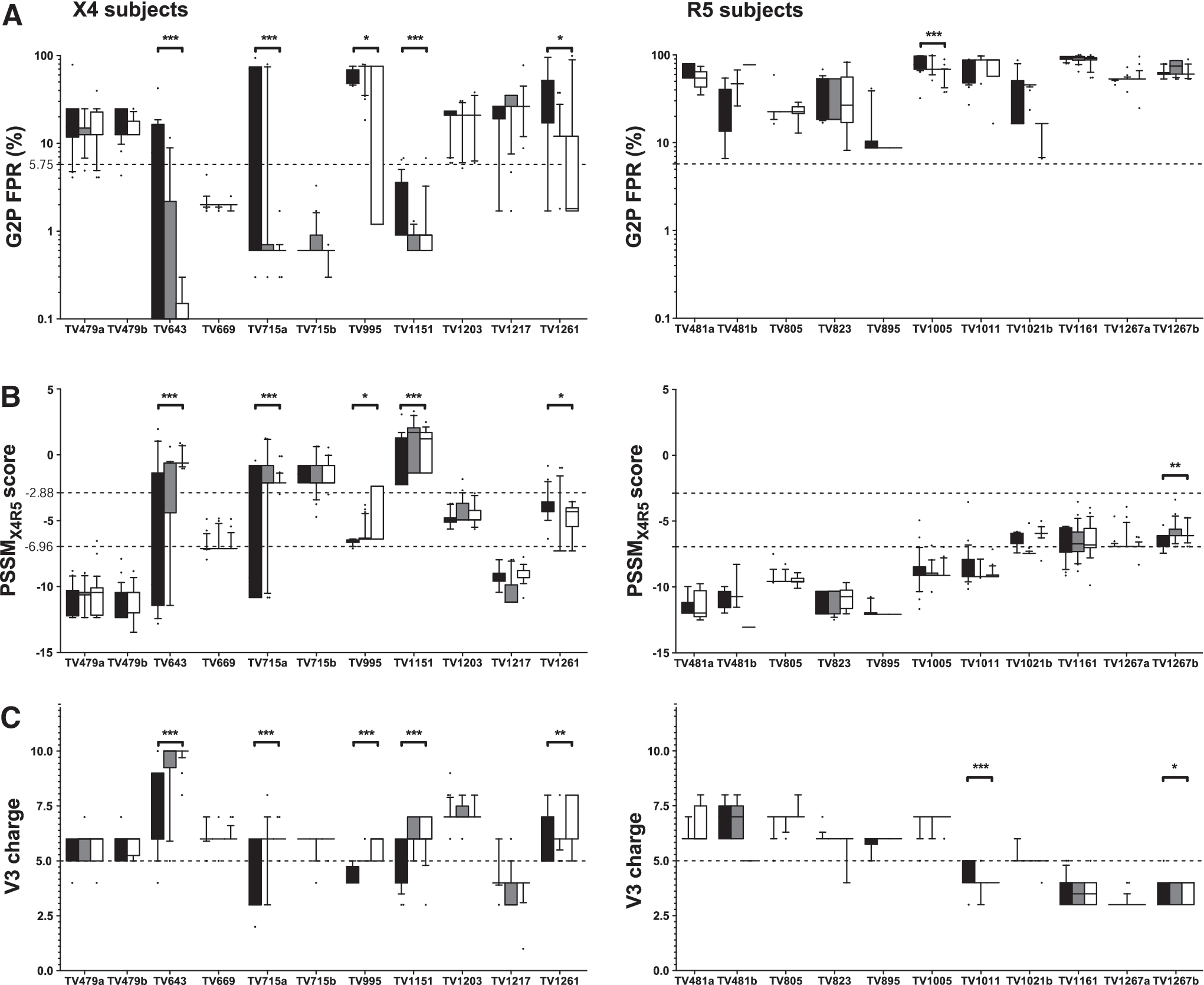
Evolution of V3 loop sequences during pregnancy in X4 and R5 subjects. Geno2pheno false-positive rate
In subject TV1151, the frequency of predicted X4-capable sequences remained above 90% throughout pregnancy. Subjects in whom the evolution of V3 sequences was not observed showed either 100% of predicted X4-capable sequences throughout pregnancy (i.e., TV669 and TV715b) or very low frequencies of X4-capable sequences over time (i.e., TV479, TV1203, and TV1217) (Table 1). In subject TV715, the frequency of X4-capable sequences rose from 45.0% to 100% during her first pregnancy (TV715a), and then remained at 100% throughout her second pregnancy (TV715b) (Table 1). The frequencies of X4-capable sequences were confirmed using PSSMX4R5 and other G2P false-positive rates (2% and 10%), although slightly different frequencies were obtained (data not shown). Tropism determinations were also confirmed using recombinant phenotypic assays in three of five subjects (three of four subjects for whom the recombinant phenotypic assay was successfully performed). Interestingly, in the case of subject TV1217, results of the recombinant phenotypic assay indicated a shift from dual-mixed in the first trimester to R5 in the second and third trimesters that was paralleled by a progressive decline in the frequencies of predicted X4-capable sequences (Table 1). Changes in V3 amino acid sequences that were associated with the evolution of coreceptor tropism were not different from those that were reported in nonpregnant subjects. In X4 subjects, mutations involved substitutions at positions 11 or 25 for positively charged residues and/or clade-specific evolution of the V3 crown and loss of PNGS at position 6 (data not shown). No such time-dependent evolution was seen in R5 subjects, except in subject TV823 (clade B), in whom a GPGK to (G/A)PGR change in the V3 crown was observed (data not shown).
In sharp contrast, of 11 pregnancies analyzed among nine of the 10 R5 subjects, a statistically significant decrease in G2P FPR was observed in only one case (TV1005; p≤0.001; Mann–Whitney U test) (Fig. 2A), and it was not accompanied by statistically significant increases in either PSSMX4R5 scores (Fig. 2B) or the net charge of V3 (Fig. 2C). Similarly, subject TV1267 manifested a statistically significant increase in PSSMX4R5 scores during her second pregnancy (TV1267b; p≤0.01; Mann–Whitney U test) (Fig. 2B), but not in G2P FPR (Fig. 2A) or in the net charge of V3 (Fig. 2C). Finally, subject TV1011 exhibited a statistically significant decline in the net charge of V3 (Fig. 2C), but not in G2P FPR (Fig. 2A) or PSSMX4R5 scores (Fig. 2B). A statistically significant evolution of G2P FPR, PSSMX4R5 score, or the net charge of V3 during pregnancy was not observed among the 10 remaining subjects.
On the whole, determinants of viral tropism defined using prediction algorithms evolved during the course of pregnancy in five subjects in whom X4 sequences were identified using G2P5.75, including TV643 (clade B), TV715 (clade C), TV995 (clade D), TV1151 (clade A1), and TV1261 (clade C). Taken together, these results reflect the strong association between the presence of X4 variants and the subsequent evolution of V3 sequences in pregnant women.
Evolution of tropism determinants in V2
We sought to verify whether changes in coreceptor tropism predicted based on the amino acid sequence of V3 were also reflected in V2. Overall, consistent with previous reports, 6 the net charge of V2 was higher in X4 subjects than in R5 subjects, with mean values of +1.72±0.34 and +0.70±0.47, respectively. However, this difference did not attain statistical significance (p=0.0524; Student's t test) (data not shown). In addition, no significant evolution of the net charge of V2 was observed during the course of pregnancy.
The V2 likelihood index was computed based on differential frequencies of occurrence of certain mutations in X4-capable and R5 viruses. This index reflects the abundance of X4-related mutations relative to R5-related mutations in a given V2 sequence (i.e., an X4 V2 sequence is expected to have a greater V2 likelihood index than an R5 V2 sequence). V2 likelihood indexes were computed for each of the 1,240 HIV-1 env sequences obtained from our study group, and, as a validation, for a reference dataset composed of 52 X4 and 185 R5 V2 sequences of clade A, B, C, and D retrieved from the Los Alamos National Laboratory HIV Sequence Database. 42 As expected, when applied to the reference dataset, the V2 likelihood index was significantly higher in X4 isolates as compared to R5 isolates, with median values of −0.3463 [IQR=(−1.040)–(+1.463)] and −1.489 [IQR=(−2.569)–(−0.6064)], respectively (p<0.0001; Mann–Whitney U test) (Fig. 3A).

V2 likelihood index in R5 and X4 isolates and evolution during pregnancy. V2 likelihood index was calculated for 52 X4 and 185 R5 V2 sequences of clade A, B, C, and D retrieved from the Los Alamos National Laboratory HIV Sequence Database
42
When applied to the global dataset from our study group, significantly higher V2 likelihood indexes were observed in X4 subjects than in R5 subjects, with median values of −0.2735 [IQR=(−0.8147)–(+1.434)] and −1.6730 [IQR=(−2.8240)–(−0.9983)], respectively (p=0.0028; Mann-Whitney U test) (Fig. 3B). In addition, the V2 likelihood index was inversely correlated with the G2P FPR (r=−0.5122; p=0.0053; Spearman's rank correlation test) (data not shown).
To examine the evolution of V2 sequences during pregnancy, longitudinal changes in the V2 likelihood index were analyzed, and statistically significant increases in the V2 likelihood index between the first and third trimesters were considered as evidence of evolution toward CXCR4 tropism. Indeed, the progressive evolution of the V2 likelihood index was readily observed in five of nine X4 subjects (55.6%), with statistically significant differences noted in subjects TV479b, TV643, TV715a, TV995, and TV1151 (p≤0.01; Mann–Whitney U test). In contrast, these variations were not seen in R5 subjects, except in the case of subject TV1021b, who exhibited a significant increase in the V2 likelihood index between the first and third trimesters (p≤0.001; Mann–Whitney U test), and subject TV1267a, who exhibited a significant reduction in the V2 likelihood index (p≤0.001; Mann–Whitney U test) (Fig. 3C). On the whole, these results are concordant with those obtained upon examination of V3 sequences (see above).
Length of V1-V2 and numbers of PNGS in subjects infected with subtype C
Because changes in tropism toward CXCR4 usage have been associated with genetic divergence in the V1-V2 loop in HIV-1 subtype C isolates, particular attention was drawn to the two subtype C-infected subjects in whom X4 sequences were predicted (TV715 and TV1261). In these two subjects, X4 clones had significantly longer V1-V2 regions (p<0.0001; Mann-Whitney U test) and greater numbers of PNGS (p<0.0015; Mann–Whitney U test) than what was observed in R5 clones, a difference that was not seen in other study subjects (data not shown). A statistically significant increase in the median length of the V1-V2 loop was observed during the course of the first pregnancy in subject TV715 (TV715a) (p<0.0001; Kruskal–Wallis test), and this increase was mainly accounted for by differences between the first and second trimester and between the first and third trimesters (p<0.001; Dunn's multiple comparison test) (Fig. 4A). Similarly, a statistically significant increase in the median number of PNGS in V1-V2 was observed in the course of the first pregnancy in subject TV715 (TV715a) (p<0.0001; Kruskal–Wallis test), this increase again being accounted for by differences between the first and second trimester and between the first and third trimesters (p<0.01; Dunn's multiple comparison test) (Fig. 4B).
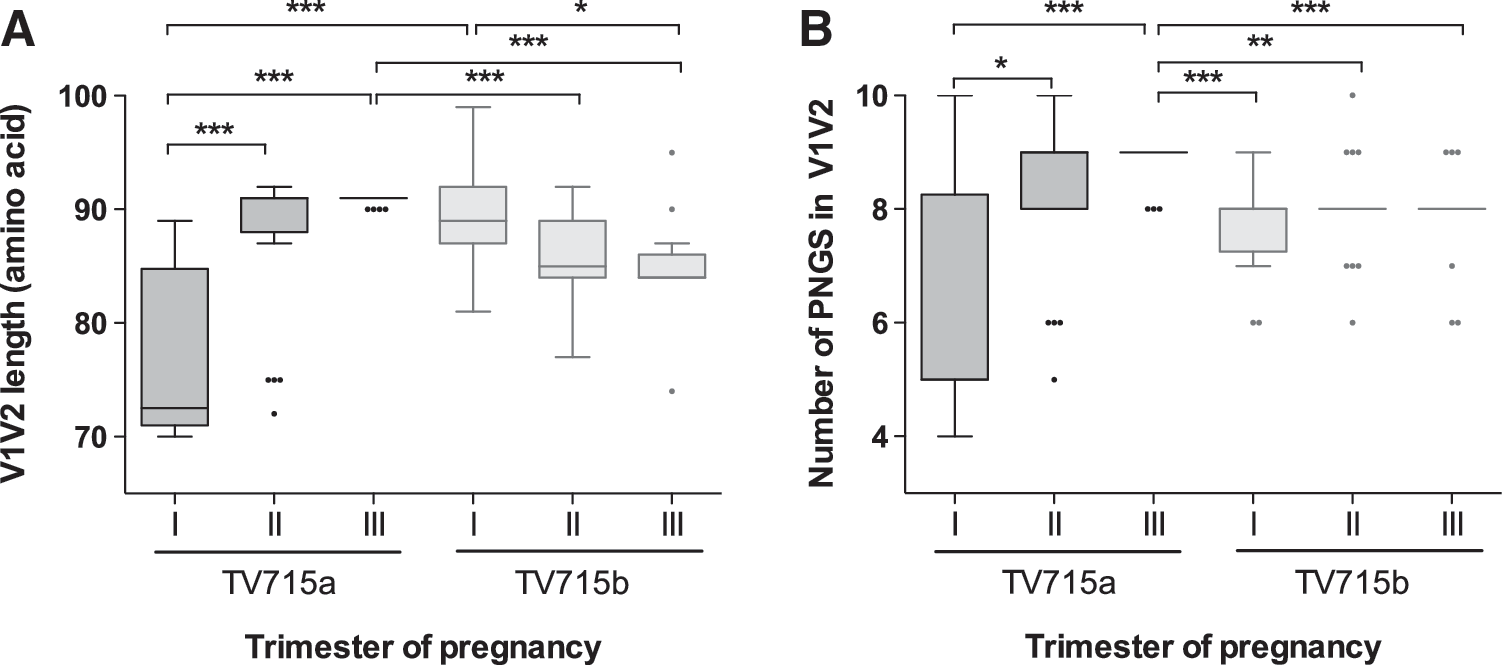
Evolution of the length of the VIV2 loop and number of potential N-linked glycosylation sites (PNGS) during two consecutive pregnancies in HIV-1 subtype C-infected subject TV715. The length in amino acids
During the second pregnancy of subject TV715 (TV715b), there was a statistically significant decline in the length of the V1-V2 loop (p=0.0028; Kruskal–Wallis test), which was explained by the difference between the first and second trimesters and between the first and third trimesters (p<0.05; Dunn's multiple comparison test). However, in contrast to TV715a, there was no statistically significant change in the number of PNGS throughout this subject's second pregnancy (p=0.6480; Kruskal–Wallis test). Overall, both median V1-V2 length and median numbers of PNGS were higher in the first pregnancy than in the second one, with differences involving values observed in the third trimester of the first pregnancy being the most salient (p<0.001, Kruskal–Wallis test; p<0.01, Dunn's multiple comparison test) (Fig. 4A and B). Such differences were not observed in subject TV1261 (data not shown).
Discussion
In this study, the evolution of coreceptor tropism determinants in the V1-V3 segment of the HIV-1 env gene was examined during the course of pregnancy in 19 women. Prediction of coreceptor tropism using G2P5.75 signaled the presence of X4-capable variants in nine of 19 study subjects independent of HIV-1 subtype. This prediction was confirmed using PSSMX4R5 in six of nine subjects, and using recombinant phenotypic assay in four of nine subjects. Thus, a concordance of 84.2% was found between G2P5.75 and PSSMX4R5, similar to what was reported by Seclen et al., although that particular study was performed using bulk sequences and a G2P false-positive rate of 10%. 38 CD4+ T cell counts were lower in X4 than in R5 subjects and six of nine X4 subjects exhibited CD4 counts ≤200, consistent with the fact that X4 variants are predominantly found in advanced HIV disease. 43 As the time from seroconversion was unknown for all study participants, the possibility that X4 subjects were infected for a longer duration of time than R5 subjects cannot be ruled out.
Moreover, a progressive and significant evolution of coreceptor tropism determinants in the V1, V2, and V3 regions (G2P FPR, PSSM score, net charge of V3, V2 likelihood index, length of V1-V2, and number of PNGS in V1-V2) as well as the increased frequency of X4-capable sequences observed over time were compatible with the ongoing evolution of viral coreceptor usage toward CXCR4 in five of 19 study subjects. The net charge of V3 is one of the main determinants of HIV-1 tropism 44,45 and R5 to X4 tropism switch is associated with an increase in the positive charge of V3, 1,46 which is necessary for an electrostatic interaction between V3 and CXCR4. 47 In terms of evolution, the charge of V3 increased significantly during pregnancy in subjects TV643, TV715a, TV995, TV1151, and TV1261, and this by itself can attest of the ongoing evolution of coreceptor usage toward CXCR4. These results are in agreement with Mild et al., 45 who showed an increase in the net charge of V3 over time in switch versus nonswitch viral populations. This increase was accompanied by a decrease in G2P FPR and an increase in PSSMX4R5 scores in these subjects, consistent with the accumulation of X4-related mutations in V3 over time. Furthermore, increasing proportions of X4-capable sequences with time were predicted by G2P5.75 and PSSMX4R5 in four of these subjects.
In three cases (TV643, TV715a, and TV1261), predicted X4 sequences were present as early as the first trimester. In two of them (TV643 and TV715), frequencies of X4-capable sequences increased progressively through pregnancy to reach 100% in the third trimester. In one subject infected with subtype D (TV995), X4 sequences appeared in the third trimester but G2P FPR had already started to decrease in the second trimester. In one subject infected with subtype A1 (TV1151), the frequency of X4-capable sequences was close to 100% throughout pregnancy while G2P FPR, PSSM scores, and the net charge of V3 showed a statistically significant evolution towards X4 tropism. In addition, recombinant phenotypic assay testing revealed that subject TV1151 was a carrier of dual-mixed virus throughout pregnancy. Taken together, these observations indicate that even when all sequences analyzed are predicted to be X4-capable, the evolution of viral tropism determinants can continue, perhaps reflective of the increased affinity/avidity for CXCR4. 48 Finally, shifting from dual-mixed to R5 tropism was observed during the course of this study, but only once in the 24 pregnancies that were analyzed (TV1217). All X4 subjects were treated with combination antiretroviral therapy, consistent with the lack of interdependence between antiretroviral treatment, the evolution of viral tropism determinants, and coreceptor switching. 49
Mounting evidence supports the involvement of V2 in determining coreceptor tropism. 4,6 Similar to V3, the net charge of V2 was higher in X4 subjects, but no evolution of net charge over time was observed. V2 likelihood indexes were computed to monitor the evolution of amino acid substitutions in V2 sequences. Calculations based on a reference dataset showed that the V2 index is higher in X4 variants than in R5 variants. Similar to a PSSM score, the V2 likelihood index reflects the relative frequency of X4-related substitutions in a given V2 sequence. However, as the association between V2 amino acid sequences and tropism is weaker than for V3, establishing a threshold value for tropism prediction is impractical. 6 Nonetheless, V2 indexes increased with the progression of pregnancy in the same subjects (i.e., TV643, TV715a, TV995, and TV1151) who showed a significant evolution of V3 features associated with CXCR4 usage (G2P FPR, PSSM scores, net charge of V3). On the whole, these results indicate that the presence of amino acid substitutions consistent with the evolution of coreceptor tropism toward CXCR4 can be observed in V2. However, whether these mutations are directly responsible for a switch in phenotype is still unclear. They could also represent gain-of-fitness mutations that compensate for fitness losses induced by other amino acid substitutions that are directly responsible for coreceptor switching, as previously discussed by Pastore et al. 5
It is well established that the length of V1-V2 and the number and pattern of PNGS within gp120 impact HIV-1 tropism, phenotype, and disease progression. 45,50,51 In addition, previous studies have suggested that lengthening of V2 may be transiently required for the R5 to X4 switch. 52,53 One explanation for the lower incidence of X4 variants within clade C is that conformational constraints in V3 limit adaptation to CXCR4. 54 It was also proposed that extensive env mutations, particularly in V1-V2, must take place for clade C viruses to use CXCR4, and that additional compensatory mutations outside of V3 are required to maintain viral fitness. 55 The lengthening of V1-V2, which was observed in two X4 subjects infected with clade C but not in R5 subjects, is characteristic of coreceptor switch in clade C and correlates with the number of PNGS. 55,56 It is thought that Env sequences, hence coreceptor tropism, can evolve in response to immune pressure. 21,57 Heavy glycosylation contributes to the efficacy of viral infection, 58 but it is also helpful to evade immune surveillance. 59,60 In addition to conformational constraints, it is possible that the V3 region of X4 variants from clade C is more susceptible to antibody neutralization than the V3 of R5 variants. 61 Therefore, the lengthening of V1-V2 and the addition of PNGS could serve to shield V3 and increase viral fitness. 62,63
While an earlier study reported the evolution of HIV-1 coreceptor usage from CCR5 to CXCR6 during pregnancy, 25 ours is the first to document evidence by G2P and PSSM of tropism switching from R5 to R5/X4 and R5/X4 to X4, and to provide evidence of evolution toward CXCR4 usage among women infected with viral isolates belonging to a wide variety of HIV-1 clades. Particular attention was paid to a subtype C-infected subject (TV715), in whom the evolution of coreceptor tropism over the course of two successive pregnancies was accompanied by a dramatic increase in the length of V1-V2 and in the number of PNGS peaking in the third trimester of the first pregnancy, followed by a significant decline during the course of the second pregnancy. Overall, these results provide substantiation for the known transmission of X4 variants from mother to child during the course of pregnancy. 12 –16,64
Why should pregnancy induce changes in viral tropism? Are these changes specifically driven by immune or environmental conditions unique to pregnancy, or does this simply reflect a general trend toward X4 evolution in the context of disease progression? Pregnancy is associated with significant modulations in maternal immune responses that are thought to play an essential role in the maintenance of tolerance to paternal alloantigens expressed by the fetus. 65 Normal pregnancy is associated with an increased total Ig synthesis, particularly of IgG. 66,67 These levels decline with the progression of pregnancy, due in part to maternofetal IgG transfer that takes place at an accelerating rate toward the end of gestation via receptor-mediated transplacental transport. 68,69 These complex changes, which take place at the level of immunoregulation, continue to be incompletely understood. 70 A dramatic increase in the length of variable loops in HIV-1 Env was associated with a vigorous humoral immune response during primary infection. 60 Thus, it is possible that observed changes reflect modulations of HIV-specific antibody responses that are associated with initiation and/or progression of pregnancy, including up-regulation of Th2 responses. 22 –24,65,71 –73 Indeed, many antibody-mediated autoimmune diseases, such as systemic lupus erythematosus, flare up during pregnancy. 74 It could thus be speculated that the emergence of X4 variants was linked to active, pregnancy-associated remodeling of an HIV-specific antibody response.
Alternatively, coreceptor switching could result from the general inhibition of cell-mediated immune responses, which is associated with the establishment and maintenance of fetomaternal tolerance. 24,68 A case in point is that depletion of CD8+ T cells in rhesus macaques infected with R5 and X4 SHIV recombinants leads to the rapid reemergence of the X4 variant, suggesting that CD8+ T cells are responsible for keeping replication of X4 variants under control, and, conversely, that impairment of this response directs coreceptor switching. 21 While the depletion of CD20+ B cells did not lead to coreceptor switching in this system, 75 treatment with progestin-based contraceptives did result in the preferential replication of X4 variants. 76 On the whole, results from the present study are compatible with a model in which the evolution of tropism determinants would be actively driven, or accelerated, by immune selective pressures exerted by differential HIV-specific immune responses associated with the initiation and/or progression of pregnancy.
However, some features of the present study should temper these interpretations. First and foremost, limitations in sample size prevented longitudinal characterization of humoral and cell-mediated HIV-specific immune responses in study participants. In addition, although multiple time points were analyzed during pregnancy, no samples could be obtained before and after pregnancy. Thus, it was not possible to analyze the potential influence that initiation and termination of pregnancy per se could have on coreceptor tropism.
To our knowledge, this is the first study to provide evidence of the evolution of tropism toward the utilization of CXCR4 during the course of pregnancy. Taken together, these results inform viral population dynamics and provide insights into maternal immunity that could lead to a better understanding of HIV pathogenesis and MTCT. Concordance between tropism prediction algorithms, various structural and physicochemical properties of gp120 variable loops, and recombinant phenotypic assays strongly supports the validity of these observations. It is possible that these observations reflect changes in selective pressures on the HIV-1 envelope resulting from the modulation of immune response associated with the initiation and/or progression of pregnancy, although we cannot exclude that tropism determinants evolved in a manner similar to or indistinguishable from that seen in nonpregnant subjects. Although safety and pharmacokinetics data are currently insufficient to recommend the use of Maraviroc in pregnancy, 77 coreceptor tropism testing should be performed in case such usage is ever considered.
Sequence Data
All sequence data were deposited in GenBank (accession numbers KF038436–KF039675).
Footnotes
Acknowledgments
Doris G. Ransy performed experiments, analyzed data, and wrote the manuscript. Alena Motorina, Natacha Merindol, and Bertine S. Akouamba performed experiments. Johanne Samson was the coordinator of the CMIS Mother-Child Cohort. Yolanda Lie and Laura Napolitano performed experiments and wrote the manuscript. Normand Lapointe and Marc Boucher provided access to study subjects and wrote the manuscript. Hugo Soudeyns designed the study, supervised the experiments, analyzed the data, and wrote the manuscript. The authors wish to thank Silvie Valois and Martine Caty for technical assistance. They also thank the Monogram Biosciences Clinical Reference Laboratory for the performance of all Trofile and Trofile DNA assays. Supported by grants from the CIHR-Health Canada Research Initiative on HIV-AIDS (grant HOP-75352) and by an infrastructure grant from le Réseau SIDA-maladies infectieuses of le Fonds de la recherche du Québec-santé (FRQ-S) to H.S. L.A.N. and Y.L. are employees of Monogram Biosciences. N.M. was the recipient of scholarships from Fondation CHU Sainte-Justine, FRQ-S, and the Cole Foundation.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.