
Abstract
Human T cell leukemia virus type-1 (HTLV-1) is prevalent worldwide with foci of high prevalence. However, to date no effective vaccine or drug against HTLV-1 infection has been developed. In efforts to define the role of antibodies in the control of HTLV-1 infection, we capitalized on the use of our previously defined anti-gp46 neutralizing monoclonal antibody (mAb) (clone LAT-27) and high titers of human anti-HTLV-1 IgG purified from HAM/TSP patients (HAM-IgG). LAT-27 and HAM-IgG completely blocked syncytium formation and T cell immortalization mediated by HTLV-1 in vitro. The addition of these antibodies to cultures of CD8+ T cell-depleted peripheral blood mononuclear cells (PBMCs) from HAM/TSP patients at the initiation of culture not only decreased the numbers of Tax-expressing cells and the production of HTLV-1 p24 but also inhibited the spontaneous immortalization of T cells. Coculture of in vitro-HTLV-1-immortalized T cell lines with autologous PBMCs in the presence of LAT-27 or HAM-IgG, but not an F(ab′)2 fragment of LAT-27 or nonneutralizing anti-gp46 mAbs, resulted in depletion of HTLV-1-infected cells. A 24-h 51Cr release assay showed the presence of significant antibody-dependent cellular cytotoxicity (ADCC) activity in LAT-27 and HAM-IgG, but not F(ab′)2 of LAT-27, resulting in the depletion of HTLV-1-infected T cells by autologous PBMCs. The depletion of natural killer (NK) cells from the effector PBMCs reduced this ADCC activity. Altogether, the present data demonstrate that the neutralizing and ADCC-inducing activities of anti-HTLV-1 antibodies are capable of reducing infection and eliminating HTLV-1-infected cells in the presence of autologous PBMCs.
Introduction
H
HTLV-1 is transmitted through contact with bodily fluids containing infected cells most often from mother to child through breast milk or via blood transfusion. It has been previously established that HTLV-1 efficiently spreads from cell to cell via the formation of virological synapses. 6 More recently, however, the formation of extracellular HTLV-1 viral particles similar to the formation of bacterial films has also been shown to be effective in viral transmission. 7 HTLV-1-antigen-expressing cells are difficult to detect at least in fresh peripheral blood mononuclear cells (PBMCs) from HTLV-1-infected individuals. 8 However, when these PBMCs are isolated from the blood and cultured in vitro, some T cells begin to produce HTLV-1 antigen 9,10 followed by spontaneous immortalization of the cells in media containing interleukin-2 (IL-2). 11
Although it has been suggested that HTLV-1 can stay dormant in infected cells and become resistant to immune effector mechanisms by ratcheting down its antigen production, 12 the continued presence of strong CD8+ cytotoxic T lymphocyte (CTL) responses 13 and readily detectable levels of antibodies specific for HTLV-1 antigens in HTLV-1-infected people 14 indicates that persistent production of HTLV-1 must occur in vivo to maintain such effector mechanisms. Escape from immune effector mechanisms by spontaneous mutation of key residues is unlikely, due to the high degree of genomic stability that is characteristic of the HTLV-1 genome. 15 It has been suggested that HTLV-1-infected cells expressing HTLV-1 antigens occur at a low enough frequency that they are constantly being eliminated by HTLV-1-specific CTL in vivo 10 without leading to immune exhaustion. Besides CTL and virus neutralizing antibodies, there has been renewed interest in the potential role of antibody-dependent cellular cytotoxicity (ADCC) as an effector mechanism against a number of viral infections. This view has been highlighted by the recent demonstration of the potential role of ADCC in the only known partially successful human RV144 trial of a vaccine against human immunodeficiency virus type-1 (HIV-1). 16 The ADCC activity against HTLV-1 was first reported by Miyakoshi et al. in 198417 followed by a number of other reports. 18 –21
So far, several lines of evidence show that the HTLV-1 envelope gp46 antigen serves as a major target of ADCC. 22 –24 Antibodies against gp46 antigen are commonly detected in the sera of HTLV-1-infected individuals. 25 –27 However, the precise role of ADCC effector mechanism(s) in controlling HTLV-1 infection has been lacking. A possible involvement of anti-HTLV-1 antibodies in the suppression of spontaneous HTLV-1 antigen expression by HTLV-1-infected cells was first reported by Tochikura et al. 28 These investigators showed that serum IgG from HTLV-1-infected donors interfered with HTLV-1 antigen expression by in vitro-cultured PBMCs from both ATL patients and healthy HTLV-1 carriers. However, the precise mechanism by which this was mediated remained unclear.
In efforts to define the role of antibodies with neutralizing and ADCC-inducing activities in the control of HTLV-1 infection, we capitalized on the use of our previously defined rat anti-gp46 neutralizing monoclonal antibody (mAb) (LAT-27) 29 and pooled human anti-HTLV-1 IgG purified from HAM/TSP patients (HAM-IgG). Studies were conducted to evaluate the potential of these antibodies to block HTLV-1 infection and eliminate HTLV-1-infected cells from autologous T cell cultures that had previously been infected with HTLV-1 in vivo or in vitro. Results of these studies show that monoclonal LAT-27 and the polyclonal HAM-IgG are not only capable of mediating neutralization and ADCC, but are also highly effective in the elimination of HTLV-1-infected cells in the presence of fresh autologous PBMCs while preventing de novo infection with HTLV-1.
Materials and Methods
Reagents
The medium used throughout was RPMI 1640 medium (Sigma-Aldrich. Inc., St. Louis, MO) supplemented with 10% fetal calf serum (FCS), 100 U/ml of penicillin, and 100 μg/ml of streptomycin (hereinafter called RPMI medium). Anti-human CD3 (clone OKT-3) and anti-CD28 (clone 28.2) mAbs were purchased from the American Type Culture Collection (Rockville, MD) and Biolegend (San Diego, CA), respectively.
The rat and mouse mAbs utilized in the studies reported herein were produced and characterized by our laboratory previously. 29 –34 These antibodies were rat IgG2b anti-gp46 (clones LAT-27 and LAT-25), rat IgG2a anti-gp46 (clone LAT-12), rat IgG2b anti-HCV (clone Mo-8), rat IgG2a anti HTLV-1 p24 (clone WAG-24), mouse IgG1 anti-HTLV-1 gp46 (clone MET-3), mouse IgG3 anti-HTLV-1 Tax (clone Lt-4), mouse IgG1 anti-p24 (clone NOR-1), and mouse IgG1 anti-HIV-1 p24 (clone 2C2). These in-house mAbs were purified from the ascites fluids of groups of CB.17-SCID mice carrying the appropriate hybridoma cell line. The ascites fluid was subjected to ammonium sulfate precipitation followed by gel filtration using Superdex G-200 (GE Healthcare, Tokyo, Japan). Aliquots of these mAbs were labeled with either fluorescein isothiocyanate (FITC), Alexafluor 488, Alexafluor 647, HRP (Dojindo, Kumamoto, Japan), or Cy-5 (GE Healthcare) according to the manufacturer's instructions. The FITC- or phycoerythrin (PE)-labeled mouse mAbs against human CD3, CD4, CD8, CD14, CD16, CD19, or CD56 and unlabeled mouse anti-CD16 and anti-CD32 mAbs were purchased from Abcam.
For cell depletion, magnetic beads labeled with anti-CD4, CD8, CD14, CD16, CD19, and antimouse IgG (Dynal) and those labeled with anti-CD56 mAb (LifeTec) were used according to the manufacturer's recommendations. Mitomycin-C (MMC) was commercially purchased from Kyowa Kirin (Tokyo, Japan) and used at 50 μg/ml in RPMI medium. A purified F(ab′)2 fragment of LAT-27 IgG generated by enzymatic digestion of LAT-27 IgG was purchased from IBL Inc. (Gunma, Japan). Human IgG was purified from pooled plasma from three normal donors (normal IgG) and three HAM patients (HAM-IgG) using protein-G affinity purification kits (GE Healthcare).
The protocols for the use of human PBMCs and animals were approved by the Human IRB and the Institutional Animal Care and Use Committee (IACUC) on clinical and animal research of the University of the Ryukyus prior to initiation of the present study.
Cell cultures
PBMCs were isolated from heparinized blood by standard density gradient centrifugation using Lympholyte (Cedarlane, Burlington, Canada). Some PBMCs were cryopreserved using a cell freezing media (Cell reservoir, Nakarai Tesque Inc., Kyoto, Japan). The method to activate PBMCs with anti-CD3 and CD28 mAbs has been described previously. 34 The HTLV-1-producing T cell lines utilized included MT-2, HUT102, IL-2-dependent CD4−CD8+ ILT-M1 cells derived from an HAM/TSP patient, CD4+CD8− ILT-H2 cells, ATL-3 cells derived from ATL patients, and a number of other T cell lines derived from normal PBMCs following in vitro immortalization by cocultivation with MMC-treated ILT-M1 cells. These cell lines were maintained in culture using RPMI medium containing 20 U/ml IL-2.
The syncytium inhibition assay was performed using an assay that involved the coculture of ILT-M1 and Jurkat cells. 35 A suspension of ILT-M1 cells in a volume of 25 μl containing 5×104 cells in 20 U/ml IL-2 media was mixed with 50 μl of serially diluted antibody to be tested in a flat-bottom 96-well microtiter plate for 5 min followed by the addition of 5×104 Jurkat cells in a volume of 25 μl of medium. After coculture for 18∼24 h at 37°C in a 5% CO2 humidified incubator, syncytium formation was microscopically observed using an inverted microscope and the minimum concentration of antibody that showed complete blocking of syncytium formation was determined. In some experiments, gp46 antigen that had been affinity purified from the culture supernatants of MT-2 cells using our anti-gp46 mAb (MET-3) antibody-coupled Sepharose 4B column (GE Healthcare) was used as a target antigen to serve as a specificity control to block the syncytia neutralization of antibodies. 36
The HTLV-1-immortalization inhibition assay was performed according to the method described previously with a slight modification. 29 Briefly, PBMCs from HTLV-1-negative healthy donors were activated with immobilized OKT-3 together with soluble anti-CD28 mAb overnight, and these cells (5×104 cells) were cocultured with an equal number of MMC-treated ILT-M1 cells in wells of round-bottom 96-well microtiter plates (BD) in 0.2 ml media containing 20 U/ml IL-2 at 37°C in a humidified 5% CO2 incubator in the presence or absence of the test antibodies. The medium was replaced with fresh IL-2-containing media with or without antibody every 3–5 days. Aliquots of the cocultured cells were monitored every week for intracellular expression of Tax antigen, and the culture supernatants were monitored for the production of p24.
The assay for inhibition of spontaneous HTLV-1 antigen expression in PBMCs from HAM/TSP patients was performed as follows. PBMCs from HAM/TSP patients after depletion of CD8+ cells were cultured in vitro at 1×106 cells/ml in 20 U/ml IL-2-containing RPMI medium at 37°C in a 24-well plate (BD) in the presence of various anti-HTLV-1 mAbs, HAM-IgG, or controls. After 24 h, cells were harvested and an aliquot stained with anti-CD3, CD4, or CD8 mAb, followed by fixation and subsequent intracellular Tax staining. The frequency and absolute cell numbers of Tax-positive cells were analyzed by flow cytometry (FCM) using the Flowcount (Coulter). The remaining cells were further cultured for 2∼6 weeks with a change of media with or without antibody every 3∼4 days. If necessary, cultures were split into 1:2 or 1:4.
The elimination of HTLV-1 antigen-expressing cells was tested as follows. The IL-2-dependent HTLV-1-infected T cell lines established from PBMCs of normal donors (2×105 cells/ml) were cocultured with autologous fresh PBMCs (2×106 cells/ml) in 20 U/ml IL-2-containing RPMI medium in triplicate in a round-bottom 96-well microtiter plate (BD) in the presence or absence of various antibodies. After initial coculture for 3 days, these cultures were split, and one was cultured in the presence and the other in the absence of fresh PBMCs and antibodies for 3 days. If necessary, these cells were further treated with antibodies and fresh PBMCs every 3 days. These cell cultures were periodically monitored for changes in the levels of Tax-expressing cells and levels of p24 production.
Flow cytometry (FCM) and enzyme-linked immunosorbent assay (ELISA)
For the detection of HTLV-1 antigen-expressing cells, sample cells were analyzed using polychromatic FCM. Briefly, live cells were Fc receptor-blocked with 2 mg/ml pooled normal human IgG in FACS buffer [phosphate-buffered saline (PBS) containing 0.2% bovine serum albumin (BSA) and 0.1% sodium azide] for 10 min on ice, and prestained with fluorescent dye-labeled mAbs for 30 min. After washing with FACS buffer, the cells were fixed in 4% paraformaldehyde (PFA) in PBS for 5 min at room temperature followed by permeablization and washing in 0.5% saponin+1% BSA (Sigma) containing FACS buffer. The cells were incubated with 0.1 μg/ml of Cy5-labeled anti-Tax antibody (clone Lt-4) for 30 min. Negative control cells were stained with Cy5-Lt-4 in the presence of 50 μg/ml of unlabeled Lt-4. These cells were analyzed using a FACSCalibur (BD) and the data obtained were analyzed using the Cell Quest software (BD). Typical staining of HTLV-1-infected T cell lines with Lt-4 and LAT-27 is also shown in Supplementary Fig. S1 (Supplementary Data are available online at
Production of HTLV-1 was determined by the measurement of the HTLV-1 core p24 antigen levels in the culture supernatants using our in-house formulated and standardized ELISA kit using a pair of anti-HTLV-1 p24 mAbs. The sensitivity of this assay was determined to be 0.5 ng/ml of p24 (data not shown).
ADCC assay
HTLV-1-immortalized target cells from healthy donors were labeled with 51Cr for 60 min as described previously 37 and mixed with varying ratios of fresh PBMCs (varying effector-to-target cell ratios) in the presence or absence of various antibodies for the indicated period of time in 20 U/ml IL-2-containing medium. Appropriate controls were included with each assay including target cells cultured in media alone (spontaneous release) and in 0.5 N HCl (100% release). After brief centrifugation, supernatants were harvested and 51Cr activity in each sample was determined using a gamma counter. The net percentage 51Cr release was calculated using standard methods as follows (cpm in experiment – cpm in medium)/(cpm in 0.5 N HCl – cpm in medium)×100. In some experiments, PBMCs were depleted of CD4+, CD8+, CD14+, CD16+, CD19+, or CD56+ cells using appropriately conjugated immunomagnetic beads and tested for their effector activity.
Statistical analysis
Data were tested for statistical significance by the Student's t test using Prism software (GraphPad Software).
Results
HTLV-1 neutralizing activities of LAT-27 and human anti-HTLV-1-IgG in vitro
The syncytium inhibition assay has been generally used to evaluate HTLV-1 neutralization titers of anti-HTLV-1 antibodies. To optimize the syncytium inhibition assay, we screened various coculture combinations of HTLV-1-producing cells with a variety of HTLV-1-negative target cells, and selected the HTLV-1-producing T cell line ILT-M1 and the HTLV-1-negative T cell line Jurkat. Overnight coculture of the ILT-M1 and Jurkat cells resulted in the generation of numerous large syncytia (Fig. 1). Using this assay system, we titrated the syncytia-blocking activity of monoclonal LAT-27 and polyclonal IgG purified from pooled plasma from HAM patients (HAM-IgG). HAM-IgG was used as a positive anti-HTLV-1 antibody control because it contained high titers of antibodies against HTLV-1 antigens (Supplementary Fig. S2). The minimum concentrations required for the “complete” inhibition of syncytia formation by LAT-27 and HAM-IgG antibodies were calculated to be 5 μg/ml and 50 μg/ml, respectively (Supplementary Fig. S3). To adjust for decay in antibody activities during cultivation at 37°C, we used LAT-27 and HAM-IgG at concentrations of 10 and 100 μg/ml, respectively, in all subsequent experiments.

Human T cell leukemia virus type-1 (HTLV-1)-mediated syncytia formation. HTLV-1+ ILT-M1 and HTLV-1− Jurkat cells were either cultured alone or cocultured at a cell-to-cell ratio of 1:1 in the presence or absence of 10 μg/ml LAT-27 for 18 h. Syncytia were microscopically observed using an inverted microscope at a magnification of 100×. Representative data from three independent experiments are shown.
To confirm the gp46 specificity of LAT-27 and HAM-IgG in this syncytium inhibition assay, an affinity-column-purified gp46 antigen 36 was added to an aliquot of either LAT-27 or HAM-IgG solution prior to cocultivation. Controls consisted of incubating an aliquot of the cocultures in media alone (shaded bars denoted by 0) or media containing 10 μg/ml of gp46 (dark bars also denoted by 0). As shown in Fig. 2, HAM-IgG incubated in media alone clearly inhibited syncytia formation in a dose-dependent manner (at 12.5∼100 μg/ml). However, preincubation of the HAM-IgG at 12.5∼100 μg/ml with 10 μg/ml of affinity-purified gp46 resulted in significant reversal of inhibition, suggesting that gp46 was the main target for the neutralization activity present in the human anti-HTLV-1 antibodies. Similar results were obtained when LAT-27 instead of HAM-IgG was preincubated with gp46 (data not shown).

Anti-gp46 antibodies are major HTLV-1 neutralizing antibodies in HAM-IgG. HAM-IgG at graded concentrations (0∼100 μg/ml) was preincubated with either affinity-purified gp46 antigen (black bars) at 10 μg/ml for 10 min or incubated with medium alone (gray shaded bars, labeled as “gp46-”) and tested for syncytia inhibition activity. The numbers of syncytia were manually counted using a “Burker-Turk” hemocytometer. Representative data from three independent experiments are shown.
LAT-27 as reported previously 29 and HAM-IgG completely inhibited HTLV-1-mediated T cell immortalization of normal activated T cells in vitro at concentrations of 10 μg/ml and 100 μg/ml, respectively (Fig. 3).
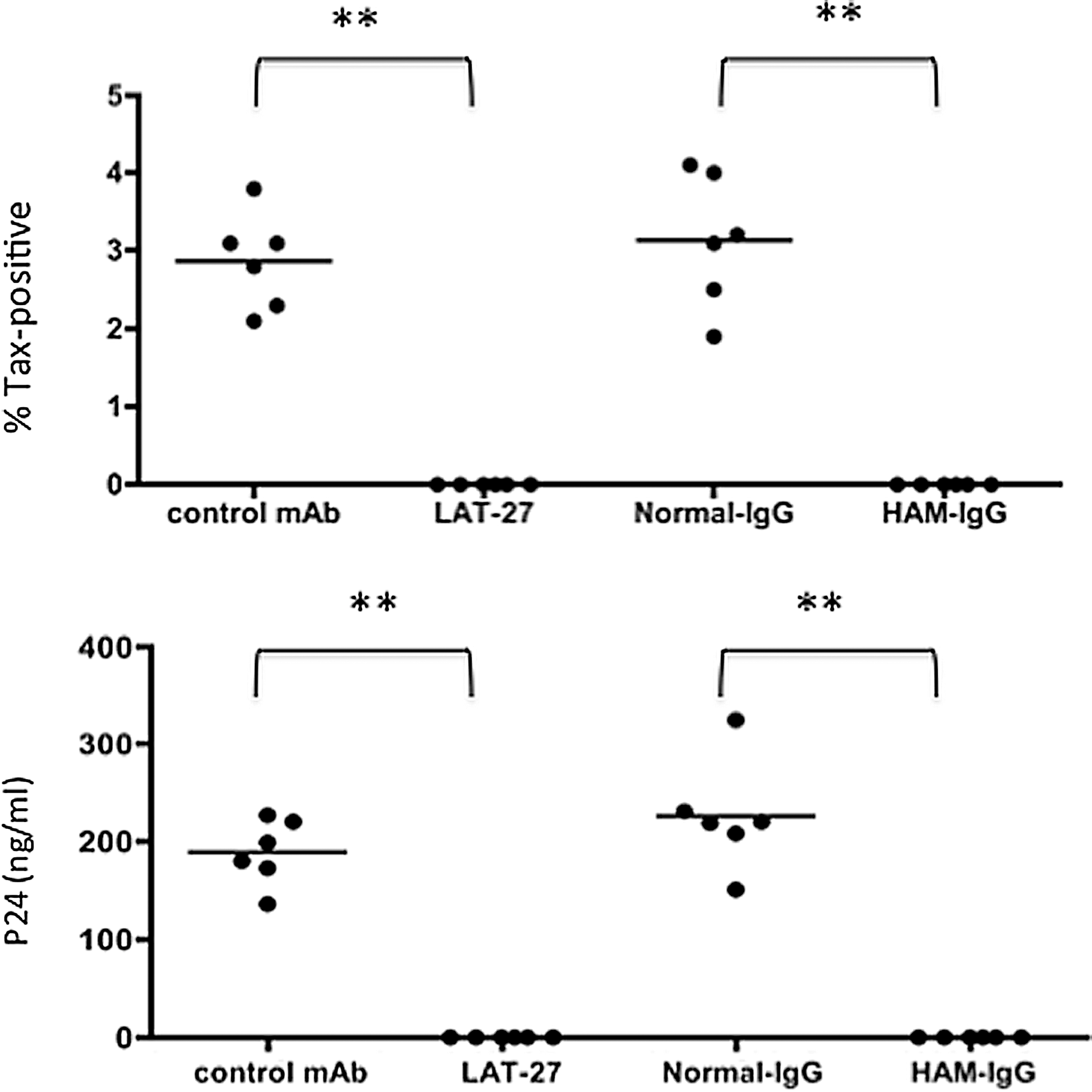
LAT-27 and HAM-IgG completely block HTLV-1-mediated T cell immortalization in vitro. Activated peripheral blood mononuclear cells (PBMCs) from normal donors were seeded into six wells of 96-well U-bottom plates (1×105 cells/0.1 ml/well) and cocultured with an equal number of mitomycin C-treated ILT-M1 cells in the presence or absence of 10 μg/ml of LAT-27 or rat isotype control, 100 μg/ml of normal human IgG, or HAM-IgG. Half of the medium was replaced every 3∼5 days with new similar fresh media, and if necessary, cultures were split into 1:2. Each data point reflects the frequency of Tax+ cells or the levels of p24 in the culture supernatants of each well 6 weeks after culture. Data shown are representative of three independent experiments. The differences between the controls and the experimental data were highly significant, denoted as **p<0.01. The negative control used for LAT-27 was an isotype control (rat IgG2b anti-HCV).
Effect of antibodies on spontaneous HTLV-1 antigen expression in vitro
To evaluate the role of anti-gp46 neutralizing antibodies against T cells carrying endogenous HTLV-1 from naturally infected donors, we cultured freshly isolated PBMCs from HAM/TSP patients in the presence or absence of various antibodies for 24 h. To exclude any potential effects of CD8+ CTL that are present within the PBMCs from the HAM/TSP patients, 10 PBMCs were depleted of CD8+ T cells prior to the assay. For quantitation of the frequencies of HTLV-1 antigen-expressing cells, we stained an aliquot of the cells for the expression of intracellular Tax antigen utilizing our standardized anti-Tax mAb, which has generally been used to detect HTLV-1-infected cells. 10,38 At a concentration of 10 μg/ml, LAT-27 reduced the frequency of Tax+ cells (Fig. 4A). This reduction was antigen and epitope specific since neither the isotype control rat IgG2b mAb (anti-HCV envelope) nor the anti-gp46 nonneutralizing mAb (LAT-25) and the other anti-gp46 nonneutralizing mAbs (clones LAT-12 and MET-3) that compete with LAT-27 in an antibody binding assay showed any detectable inhibitory effect (data not shown). The reduction in the frequency of Tax+ cells by LAT-27 was partially reversed by a mixture of anti-CD16 and anti-CD32 mAbs when added at the initiation of the assay, suggesting an involvement of Fc receptors in this reduction assay.
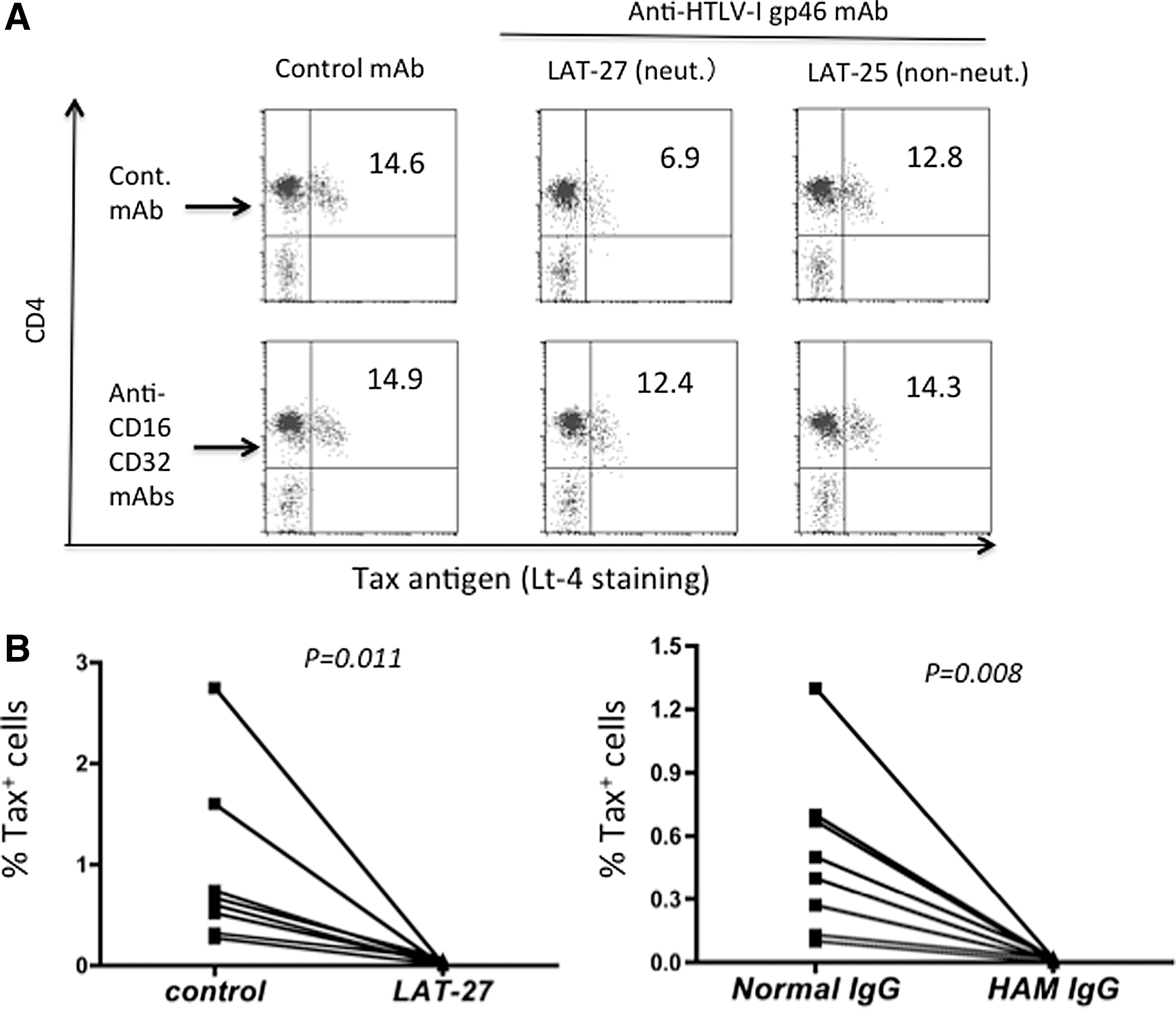
Reduction of Tax-expressing cells in in vitro cultures of PBMCs from HAM patients in the presence of HTLV-1 neutralizing monoclonal antibody (mAb).
As shown in Fig. 4B, after prolonged culture (2 weeks) the suppressive effect of LAT-27 became more evident since there remained few if any Tax+ cells in the LAT-27-treated cultures of PBMCs from each of the HAM patients tested. A similar suppressive effect was observed for HAM-IgG but not normal human IgG (Fig. 4B). It should be noted that in the present culture conditions, similar to what has been generally observed for the PBMC cultures from HTLV-1-infected donors, the frequency of Tax+ cells gradually decreased during 2 weeks in culture even in IL-2 medium alone and thus it was not likely due to an effect of the addition of the control rat isotype IgG or normal IgG. Spontaneous immortalization of T cells by HTLV-1 was observed in the PBMC cultures from two-thirds of the HAM patients treated with medium alone, isotype control, or normal IgG, but not in those treated with LAT-27 or HAM-IgG, as judged 6 weeks after culture (data not shown).
Importantly, neither LAT-27 nor HAM-IgG (data not shown) showed any detectable blocking effects on HTLV-1 Tax expression (Fig. 5) and p24 antigen production (data not shown) in long-term cultured HTLV-1-infected cell lines. It is known that the levels of Tax-positive cells vary depending on the cell lines being utilized due to DNA methylation, hypoacetylation of histones, or epigenetic changes of provirus. 39 Thus, we reasoned that the addition of the neutralizing antibodies blocks de novo expansion of HTLV-1 infection and/or eliminates the HTLV-1 gp46 expressing T cells via an FcR-dependent pathway in combination with effector cells contained within the PBMCs.

LAT-27 alone does not affect long-term cultured HTLV-1-infected T cells. A standard HTLV-1-infected cell line HUT-102 (HUT), an IL-2-dependent CD4+ T cell line (ATL-3, generated from an ATL patient), and an IL-2-dependent CD8+ T cell line (ILT-M1) were cultured in the presence of 10 μg/ml of either LAT-27 or isotype control (control) for 4 days, and the frequencies of Tax+ cells were determined by flow cytometry (n=3).
FcR-dependent elimination of HTLV-1-infected cells by antibodies
Due to the limitations on the availability of PBMC samples from HAM/TSP patients, we established a number of IL-2-dependent HTLV-1-infected CD4+ T cell lines from PBMCs of normal donors to determine whether ADCC was involved in the suppression of HTLV-1-infected cells. These HTLV-1+CD4+ T cell lines were cocultured with autologous fresh PBMCs in the presence or absence of various antibodies including F(ab′)2 of LAT-27, which showed HTLV-1 neutralization at a minimum concentration of 2.5 μg/ml (data not shown). HAM-IgG was included as an ADCC-positive control. After 3 days in culture, these cells were stained for cell surface CD4 and intracellular Tax antigen, and analyzed on a gated population of cells that displayed high forward and side scatters, which included a majority of the HTLV-1-infected CD4+ T cells but not normal PBMCs. However, no detectable reduction of Tax+ cells was observed in the cultures treated with either LAT-27 or HAM-IgG cocultured in the presence of PBMCs (data not shown). Thus, these cells were washed and cocultured again for an additional 3 days with the same antibodies and fresh PBMCs.
As shown in Fig. 6A, although fresh PBMCs alone reduced the frequency of Tax+ cells to some extent, a marked net reduction was seen in the presence of LAT-27 and HAM-IgG. In a similar fashion, the production of HTLV-1 p24 in the culture supernatants was markedly reduced by LAT-27 and HAM-IgG in the presence of autologous PBMCs. As shown in Fig. 6B, when these cultures were exposed one more time to the same antibodies and fresh PBMCs, LAT-27 IgG and HAM-IgG, but not F(ab′)2 of LAT-27 or normal IgG, further reduced the frequency of Tax+ cells. These data suggest that the addition of LAT-27 as well as HAM-IgG eliminates the HTLV-1 gp46 antigen-expressing cells via an FcR-dependent manner while blocking the spread of HTLV-1 to new target cells including fresh PBMCs in the same cell cultures in vitro. The involvement of complement-dependent cytotoxicity was ruled out because the fetal calf serum used in the present study was heat inactivated prior to use.
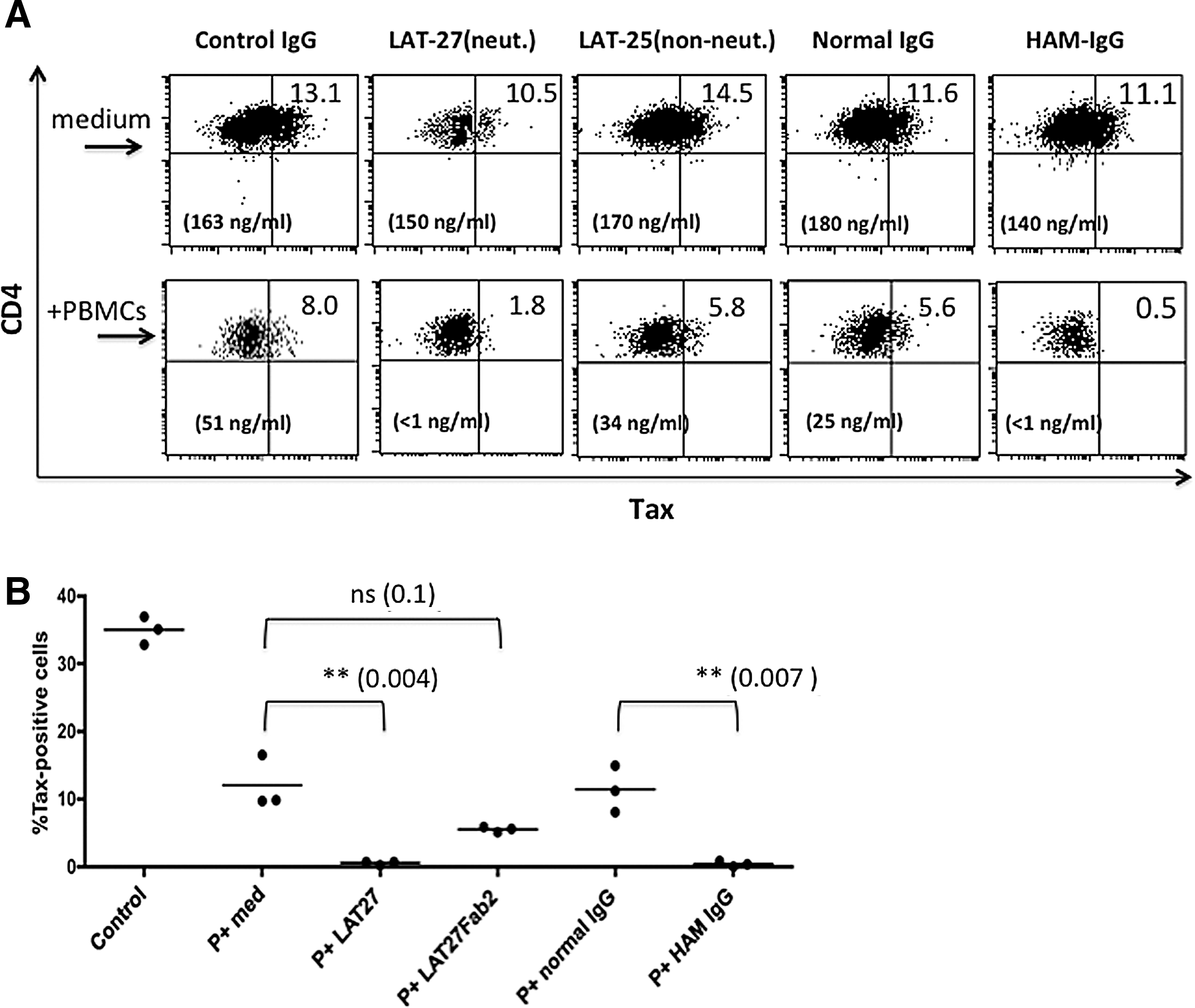
Elimination of Tax+ cells and reduction of HTLV-1 p24 production in IL-2-dependent HTLV-1-infected T cells cocultured with autologous PBMCs in the presence of LAT-27 or HAM-IgG.
ADCC against HTLV-1-infected cells by LAT-27
To examine whether LAT-27 could mediate ADCC in the present culture conditions, IL-2-dependent HTLV-1-infected T cells established from normal donors were labeled with 51Cr and cocultured with fresh autologous PBMCs in the presence or absence of antibodies. Significant ADCC activity was induced by HAM-IgG, but not LAT-27, by 6 h (data not shown). However, after 24 h at a high effector-to-target cell ratio, LAT-27, but not the F(ab′)2 fragment of LAT-27, showed significant cytotoxicity (p<0.01) (Fig. 7A). When the effector PBMCs were depleted of either CD16+ or CD56+ cells, but not CD14+ or CD19+ cells, the ADCC activity mediated by either LAT-27 or HAM-IgG was significantly reduced (p<0.01) (Fig. 7B and C). These data suggest that the CD16+ CD56+ subpopulation of PBMCs [representing natural killer (NK) cells] were most likely the main effector cells involved in the cell lysis. These results demonstrate that the monoclonal LAT-27, similar to the polyclonal HAM-IgG, is able to induce ADCC against HTLV-1-infected cells by autologous NK cells while protecting the spread of new infection with HTLV-1.

The CD16+ CD56+ PBMCs mediate antibody-dependent cellular cytotoxicity (ADCC) in the presence of LAT-27 or HAM-IgG.
Discussion
The present study demonstrates that the monoclonal anti-HTLV-1 gp46 antibody clone LAT-27 generated by our laboratory mediates both HTLV-1 neutralization and HTLV-1-specific ADCC, and such ADCC activity might be capable of eliminating HTLV-1-infected T cells in vitro in the presence of autologous fresh PBMCs. Although fresh PBMCs alone showed a partial but significant inhibitory activity against HTLV-1-infected cells during prolonged in vitro cultivation, the data obtained here suggest that the HTLV-1-specific ADCC activity is the direct mechanism for this eradication. Similar suppressive activities were demonstrated for human IgG from HAM patients. This mechanism may explain the previous findings reported by Tochikura et al. 28 on the HTLV-1 suppressing activity of human anti-HTLV-1 antibodies. Furthermore, this mechanism may also explain in part why HTLV-1 antigen-expressing cells are not found in vivo in anti-HTLV-1 antibody-positive individuals. Although it is not known where and when HTLV-1 is produced in vivo in the infected individual, the continued presence of CD8+ T cells and antibodies specific for HTLV-1 indicates that HTLV-1 should be expressed periodically. Based on the results presented in this article, it might be possible that HTLV-1 expression occurs upon T cell stimulation in the periphery, but as soon as the cells express HTLV-1 gp46 antigen they might be instantly killed by the combination of anti-HTLV-1 ADCC-inducing antibodies and activated NK cells.
We submit that the addition of fresh PBMCs to the autologous HTLV-1-producing T cell cultures may result in it becoming readily infected and immortalized by HTLV-1. Thus, it is clear that the presence of neutralizing antibody is essential for the prevention of new infection of PBMCs and since ADCC effector mechanisms are functional during this time period, their contribution to the control of infection deserves merit. Interestingly, the ADCC induced by LAT-27 progressed slowly and the elimination of Tax+ cells became evident only after two consecutive exposures every 3 days in the present cell culture conditions. Since there was heterogeneity of the intensity of gp46 expression among cells in a single HTLV-1-infected cell line (data not shown), the findings suggest that the lysis of such gp46low cells by ADCC requires a prolonged incubation period. Alternatively, since the repeated exposure against PBMCs resulted in an accumulation of live PBMCs, it is possible that a large number of effector fresh PBMCs might be required for the complete eradication by LAT-27, possibly due to the relatively low affinity of LAT-27 for human FcR.
Cell depletion experiments in the present study showed that the effector cells involved in the HTLV-1-specific ADCC in fresh PBMCs were either CD16+ or CD56+ cells, representing the cytolytic human NK cell subset, although it remains to be confirmed with purified NK cells. Because there are abundant circulating NK cells in the periphery in healthy donors, these findings strongly suggest that the HTLV-1-specific ADCC responses in the presence of neutralizing antibodies might have a role in controlling HTLV-1 in vivo in concert with HTLV-1-sepecific CTL responses in healthy HTLV-1 carriers. This view is supported by the findings that the ADCC effector function of PBMCs is lower in both HAM/TSP and ATL patients than healthy HTLV-1 carriers or normal donors, 17,40 suggesting that defects in functional ADCC activities may contribute to the onset of HTLV-1-related diseases.
The level of ADCC of HTLV-1+ cells by LAT-27 was weaker than that induced by human polyclonal anti-HTLV-1 IgG. This might be due to the fact that LAT-27 is of rat origin and recognizes a single epitope on the gp46 (amino acids 191–196) 29 in contrast to the fact that HAM-IgG is of human origin and consists of high titers of polyclonal antibodies against multiple epitopes on gp46. In addition, it has been shown that mouse and rat IgG exhibit different ADCC activities with human NK cells depending on their subclasses, and that rat IgG2b (the subclass of LAT-27), but not IgG2a, triggers effective ADCC with human NK cells. 41 Along these lines, it is possible that a humanized form of LAT-27 utilizing the human IgG1- or IgG3-Fc portion as a backbone would be far more effective than even the rat IgG2b of LAT-27.
This hypothesis has been confirmed by preliminary experiments using humanized LAT-27 consisting of human IgG1, which was generated in collaboration with Dr. Shimizu of IBL Inc. (Tanaka et al., unpublished observations). In addition, epitope specificity and/or the affinity of anti-gp46 antibodies may also be involved in determining the ADCC-inducing activities. For example, LAT-25, which belongs to the rat IgG2b subclass and recognizes a C-terminal region of the gp46, did not eradicate HTLV-1+ cells (Fig. 7). Similarly, Kuroki et al. showed that a human mAb recognizing gp46 amino acids 191–196 (similar to the epitope recognized by LAT-27) could induce ADCC, but another human mAb recognizing the gp46 amino acids 187–193 could not, even though the two mAbs bind similarly to the cell surface of HTLV-1-infected cells and belong to the ADCC-inducing human IgG1. 22
It remains to be determined whether there are clonal populations of human IgGs that can mediate both the neutralization and ADCC against HTLV-1. So far, it has been shown that the two activities could be operating separately by different epitope-specific human mAbs against gp46. 22 Recently, Kuo et al. 24 showed that both neutralizing and nonneutralizing mouse anti-gp46 mAbs can activate neutrophils and mediate its burst activity in the presence of an HTLV-1-infected MT-2 cell line, and concluded that HTLV-1-specific ADCC capacity is not coupled to the neutralizing capacity of the antibody. Thus, these articles highlight the finding of LAT-27 as a special antibody. Analyses of the conformational and antigenic structure of gp46 expressed on the cell surface will be necessary to address this issue further.
Another possible target for ADCC on HTLV-1-expressing cells is the envelope gp21; however, it has been unclear whether human anti-gp21 antibodies function in ADCC. In addition, the recent finding that the glycosylation of Fc-IgG plays an important role in anti-HIV-1 ADCC effector mechanisms 42 suggests that this issue needs to also be considered in the evaluation of anti-HTLV-1 gp46 antibodies and for vaccine formulations in general. Nevertheless, it is clear that the simultaneous operation of neutralization and ADCC by single or polyclonal antibodies is essential to recognize and eliminate HTLV-1+ cells since not only T cells but also the NK cells are permissive to HTLV-1 infection. 43
The present study also showed that fresh PBMCs had a partial and significant but not complete suppressive activity against autologous HTLV-1-infected cells in the absence of anti-HTLV-1 antibodies. Our preliminary experiments indicate that monocytes might be involved in this partial suppression because PBMCs depleted of CD14+ cells, but not of NK cells, were no longer suppressive in the absence of LAT-27 (data not shown). Since HTLV-1-infected T cells are continuously activated due to the Tax antigen, one possible mechanism is a monocyte-dependent cell death (MDCD) against activated autologous T cells. 44 Further studies are in progress to address this mechanism.
Based on the data presented herein, it is suggested that humanized LAT-27 mAb might have potential as a passive vaccine against HTLV-1 infection for HTLV-1-uninfected individuals at high risk of HTLV-1 infection, including babies born to HTLV-1 carriers and drug abusers who are also at high risk of HIV infection, and for HTLV-1 carriers whose anti-HTLV-1 neutralizing and ADCC-inducing antibody titers are low. One concern is the potential interference of LAT-27 activity by other nonneutralizing or non-ADCC-inducing antibodies that may interfere with the binding of LAT-27 to gp46. We have performed some experiments and obtained data showing that LAT-12, which blocked the binding of LAT-27 to HTLV-1-infected cells, did not interfere with either LAT-27-mediated syncytium blocking 29 and/or the eradication of HTLV-1-infected cells with autologous PBMCs (Supplementary Fig. S4). It seems likely that the binding affinities of neutralizing antibodies to gp46 expressed on actively living cells are higher than those of nonneutralizing antibodies. Thus, validation of humanized LAT-27 in animal models is currently one of our objectives.
Footnotes
Acknowledgments
This work was supported by grants from the Ministry of Education, Culture, Sports, Science, and Technology and the Ministry of Health, Labor, and Welfare of Japan.
Y.Tak. and A.H. carried out the ADCC assays. R.T. and A.K. produced, purified, labeled monoclonal antibodies, confirmed their specificities, and made in-house EILSA for p24. M.S. participated in the determination of proviral loads and performed the statistical analysis. M.K. established HTLV-1-infected cells from patients and participated in the design of the study. A.A.A. participated in the design of the study and helped to draft the manuscript. Y.T. conceived the study, participated in its design and coordination, carried out the coculture assays, and drafted the manuscript. All authors read and approved the final manuscript.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.